
Trait-Based Vaccination of Individual Meerkats (Suricata suricatta) against Tuberculosis Provides Evidence to Support Targeted Disease Control


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Data Collection
2.3. Treatments
2.4. Data Analysis
2.4.1. Infection Data
2.4.2. Longevity Data
2.5. Ethical Approval
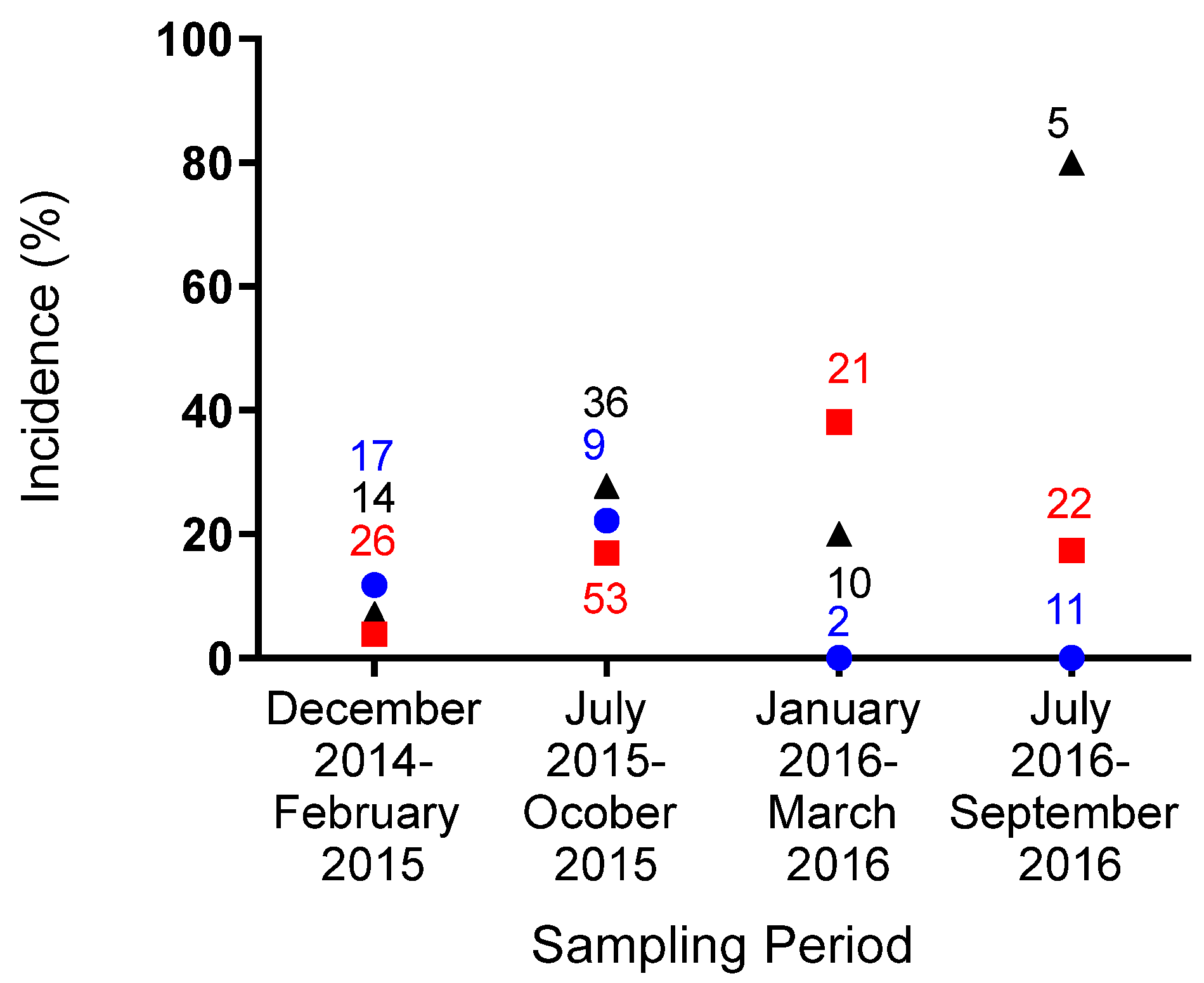
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woolhouse, M.E.J.; Dye, C.; Etard, J.F.; Smith, T.; Charlwood, J.D.; Garnett, G.P.; Hagan, P.; Hii, J.L.K.; Ndhlovu, P.D.; Quinnell, R.J.; et al. Heterogeneities in the transmission of infectious agents: Implications for the design of control programs. Proc. Natl. Acad. Sci. USA 1997, 94, 338–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolzoni, L.; Real, L.; de Leo, G. Transmission Heterogeneity and Control Strategies for Infectious Disease Emergence. PLoS ONE 2007, 2, e747. [Google Scholar] [CrossRef] [Green Version]
- Heesterbeek, J.A.P.; Dietz, K. The concept of Ro in epidemic theory. Stat. Neerl. 1996, 50, 89–110. [Google Scholar] [CrossRef]
- Anderson, R.; May, R.M. Population biology of infectious diseases: Part I. Nature 1979, 280, 361–367. [Google Scholar] [CrossRef] [PubMed]
- John, T.J.; Samuel, R. Herd immunity and herd effect: New insights and definitions. Eur. J. Epidemiol. 2000, 16, 601–606. [Google Scholar] [CrossRef]
- Hampson, K.; Dushoff, J.; Cleaveland, S.; Haydon, D.; Kaare, M.; Packer, C.; Dobson, A. Transmission Dynamics and Prospects for the Elimination of Canine Rabies. PLoS Biol. 2009, 7, e1000053. [Google Scholar] [CrossRef]
- Anderson, R.; May, R. Immunisation and herd immunity. Lancet 1990, 335, 641–645. [Google Scholar] [CrossRef]
- Kim, T.H.; Johnstone, J.; Loeb, M. Vaccine herd effect. Scand. J. Infect. Dis. 2011, 43, 683–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd-Smith, J.O.; Schreiber, S.J.; Kopp, P.E.; Getz, W. Superspreading and the effect of individual variation on disease emergence. Nature 2005, 438, 355–359. [Google Scholar] [CrossRef]
- Fine, P.E.M. Herd immunity: History, theory, practice. Epidemiol. Rev. 1993, 15, 265–302. [Google Scholar] [CrossRef]
- Wallinga, J.; Heijne, J.C.M.; Kretzschmar, M. A Measles Epidemic Threshold in a Highly Vaccinated Population. PLoS Med. 2005, 2, e316. [Google Scholar] [CrossRef] [PubMed]
- Rhyan, J.C.; Nol, P.; Quance, C.; Gertonson, A.; Belfrage, J.; Harris, L.; Straka, K.; Robbe-Austerman, S. Transmission of brucellosis from elk to cattle and bison, Greater Yellowstone area, U.S.A. 2002–2012. Emerg. Infect. Dis. 2013, 19, 1992–1995. [Google Scholar] [CrossRef]
- Chambers, M.A.; Carter, S.P.; Wilson, G.J.; Jones, G.; Brown, E.; Hewinson, R.G.; Vordermeier, M. Vaccination against tuberculosis in badgers and cattle: An overview of the challenges, developments and current research priorities in Great Britain. Veter-Rec. 2014, 175, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Medlock, J.; Galvani, A.P. Optimising Influenza Vaccine Distribution. Science 2009, 325, 1705–1708. [Google Scholar] [CrossRef] [PubMed]
- Carne, C.; Semple, S.; Morrogh-Bernard, H.; Zuberbühler, K.; Lehmann, J. Predicting the Vulnerability of Great Apes to Disease: The Role of Superspreaders and Their Potential Vaccination. PLoS ONE 2013, 8, e84642. [Google Scholar] [CrossRef] [Green Version]
- Cross, P.C.; Getz, W.M. Assessing vaccination as a control strategy in an ongoing epidemic: Bovine tuberculosis in African buffalo. Ecol. Model. 2006, 196, 494–504. [Google Scholar] [CrossRef]
- Delahay, R.J.; Wilson, G.J.; Smith, G.C.; Cheeseman, C.L. Vaccinating badgers (Meles meles) against Mycobacterium bovis: The ecological considerations. Vet. J. 2003, 166, 43–51. [Google Scholar] [CrossRef]
- Blancou, J.; Artois, M.; Gilot-Fromont, E.; Kaden, V.; Rossi, S.; Smith, G.C.; Hutchings, M.R.; Chambers, M.A.; Houghton, S.; Delahay, R.J. Options for the control of disease 1: Targetting the infectious or parasitic agent. In Management of Disease in Wild Mammals; Delahay, R.J., Smith, G.C., Hutchings, M.R., Eds.; Springer: Tokyo, Japan, 2009. [Google Scholar]
- Anderson, R.; May, R.M. Spatial, temporal, and genetic heterogeneity in host populations and the design of immunization programmes. Math. Med. Biol. A J. IMA 1984, 1, 233–266. [Google Scholar] [CrossRef]
- Hadjichrysanthou, C.; Sharkey, K.J. Epidemic control analysis: Designing targeted intervention strategies against epidemics propagated on contact networks. J. Theor. Biol. 2015, 365, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Rushmore, J.; Caillaud, D.; Hall, R.J.; Stumpf, R.M.; Meyers, L.; Altizer, S. Network-based vaccination improves prospects for disease control in wild chimpanzees. J. R. Soc. Interface 2014, 11, 20140349. [Google Scholar] [CrossRef] [Green Version]
- Drewe, J.A.; Madden, J.R.; Pearce, G.P. The social network structure of a wild meerkat population: 1. Inter-group interactions. Behav. Ecol. Sociobiol. 2009, 63, 1295–1306. [Google Scholar] [CrossRef]
- Madden, J.R.; Drewe, J.A.; Pearce, G.P.; Clutton-Brock, T.H. The social network structure of a wild meerkat population: 2. Intragroup interactions. Behav. Ecol. Sociobiol. 2009, 64, 81–95. [Google Scholar] [CrossRef]
- Madden, J.R.; Drewe, J.A.; Pearce, G.P.; Clutton-Brock, T.H. The social network structure of a wild meerkat population: 3. Position of individuals within networks. Behav. Ecol. Sociobiol. 2011, 65, 1857–1871. [Google Scholar] [CrossRef]
- Drewe, J.A. Who infects whom? Social networks and tuberculosis transmission in wild meerkats. Proc. R. Soc. B Boil. Sci. 2009, 277, 633–642. [Google Scholar] [CrossRef]
- Patterson, S.; Drewe, J.A.; Pfeiffer, D.U.; Clutton-Brock, T.H. Social and environmental factors affect tuberculosis related mortality in wild meerkats. J. Anim. Ecol. 2017, 86, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Clutton-Brock, T.H.; Brotherton, P.N.M.; Russell, A.F.; O’Riain, M.J.; Gaynor, D.; Kansky, R.; Griffin, A.; Manser, M.; Sharpe, L.; McIlrath, G.M.; et al. Cooperation, control, and concession in meerkat groups. Science 2001, 291, 478–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clutton-Brock, T.H.; Brotherton, P.N.M.; Smith, R.; McIlrath, G.M.; Kansky, R.; Gaynor, D.; O’Riain, M.J.; Skinner, J.D. Infanticide and expulsion of females in a cooperative mammal. Proc. R. Soc. B Boil. Sci. 1998, 265, 2291–2295. [Google Scholar] [CrossRef] [PubMed]
- Clutton-Brock, T.H. Reproductive skew, concessions and limited control. Trends Ecol. Evol. 1998, 13, 288–292. [Google Scholar] [CrossRef]
- Doolan, S.P.; Macdonald, D.W. Diet and foraging behaviour of group-living meerkats, Suricata suricatta, in the southern Kalahari. J. Zool. 1996, 239, 697–716. [Google Scholar] [CrossRef]
- Drewe, J.A. Social Networks and Infectious Disease Transmission: Epidemiology of Tuberculosis in Wild Meerkats. Ph.D. Thesis, Cambridge University, Cambridge, UK, 2008. [Google Scholar]
- Thavarajah, N.; Fenkes, M.; Clutton-Brock, T. The determinants of dominance relationships among subordinate females in the cooperatively breeding meerkat. Behaviour 2014, 151, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Dippenaar, A.; Parsons, S.D.C.; Sampson, S.L.; van der Merwe, R.; Drewe, J.A.; Abdallah, A.M.; Siame, K.K.; van Pittius, N.C.G.; van Helden, P.D.; Pain, A.; et al. Whole genome sequence analysis of Mycobacterium suricattae. Tuberculosis 2015, 95, 682–688. [Google Scholar] [CrossRef] [Green Version]
- Clutton-Brock, T.H.; Manser, M. Meerkats: Cooperative breeding in the Kalahari. In Cooperative Breeding in Vertebrates: Studies of Ecology, Evolution, and Behaviour; Koenig, W.D., Dickinson, J.L., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 294–317. [Google Scholar]
- Drewe, J.A.; Dean, G.S.; Michel, A.L.; Pearce, G.P. Accuracy of three diagnostic tests for determining Mycobacterium bovis infection status in live-sampled wild meerkats (Suricata suricatta). J. Vet. Diagn. Investig. 2009, 21, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Clarke, C.; Patterson, S.J.; Drewe, J.A.; van Helden, P.D.; Miller, M.A.; Parsons, S.D.C. Development and evaluation of a diagnostic cytokine-release assay for Mycobacterium suricattae infection in meerkats (Suricata suricatta). BMC Veter-Res. 2016, 13, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spong, G.F.; Hodge, S.J.; Young, A.J.; Clutton-Brock, T.H. Factors affecting the reproductive success of dominant male meerkats. Mol. Ecol. 2008, 17, 2287–2299. [Google Scholar] [CrossRef] [PubMed]
- Drewe, J.; Foote, A.; Sutcliffe, R.; Pearce, G. Pathology of Mycobacterium bovis infection in wild meerkats (Suricata suricatta). J. Comp. Pathol. 2009, 140, 12–24. [Google Scholar] [CrossRef]
- Young, A.J.; Spong, G.; Clutton-Brock, T. Subordinate male meerkats prospect for extra-group paternity: Alternative reproductive tactics in a cooperative mammal. Proc. R. Soc. B Boil. Sci. 2007, 274, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Kutsukake, N.; Clutton-Brock, T.H. Grooming and the value of social relationships in cooperatively breeding meerkats. Anim. Behav. 2010, 79, 271–279. [Google Scholar] [CrossRef]
- Patterson, S.J.; Clarke, C.; Clutton-Brock, T.H.; Miller, M.A.; Parsons, S.D.C.; Pfeiffer, D.U.; Vergne, T.; Drewe, J.A. Combining Analytical Approaches and Multiple Sources of Information to Improve Interpretation of Diagnostic Test Results for Tuberculosis in Wild Meerkats. Animals 2021, 11, 3453. [Google Scholar] [CrossRef] [PubMed]
- Patterson, S. Translating Theory into Practice: Targeted Disease Control in Wild Meerkats (Suricata Suricatta). Ph.D. Thesis, University of London, London, UK, 2017. [Google Scholar]
- Microsoft Corporation. Microsoft Excel. 2010. Available online: https://office.microsoft.com/excel (accessed on 5 January 2022).
- Cox, D.R.; Oakes, D. Time Dependant Covariates. In Analysis of Survival Data; Monographs on Statistics and Applied Probability; Cox, D.R., Oakes, D., Eds.; Chapman and Hall/CRC: Boca Raton, FL, USA, 1984. [Google Scholar]
- Therneau, T.M.; Grambsch, P.M. Modelling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Sharpe, L.L. Frequency of social play does not affect dispersal partnerships in wild meerkats. Anim. Behav. 2005, 70, 559–569. [Google Scholar] [CrossRef]
- Grambsch, P.M.; Therneau, T.M. Proportional hazards tests and diagnostics based on weighted residuals. Biometrika 1994, 81, 515–526. [Google Scholar] [CrossRef]
- Carter, S.P.; Chambers, M.A.; Rushton, S.P.; Shirley, M.; Schuchert, P.; Pietravalle, S.; Murray, A.; Rogers, F.; Gettinby, G.; Smith, G.C.; et al. BCG Vaccination Reduces Risk of Tuberculosis Infection in Vaccinated Badgers and Unvaccinated Badger Cubs. PLoS ONE 2012, 7, e49833. [Google Scholar] [CrossRef] [PubMed]
- Paniw, M.; Maag, N.; Cozzi, G.; Clutton-Brock, T.; Ozgul, A. Life history responses of meerkats to seasonal changes in extreme environments. Science 2019, 363, 631–635. [Google Scholar] [CrossRef] [Green Version]
- Sah, P.; Mann, J.; Bansal, S. Disease Implications of animal social network structure: A synthesis across social systems. J. Anim. Ecol. 2018, 87, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Colditz, G.; Berkey, C.S.; Mosteller, F.; Brewer, T.F.; E Wilson, M.; Burdick, E.; Fineberg, H.V. The efficacy of bacillus Calmette-Guerin vaccination of newborns and infants in the prevention of tuberculosis: Meta-analyses of the published literature. Paediatrics 1995, 96, 29–35. [Google Scholar] [CrossRef]
- Hawn, T.R.; Day, T.A.; Scriba, T.J.; Hatherill, M.; Hanekom, W.A.; Evans, T.G.; Churchyard, G.; Kublin, J.G.; Bekker, L.-G.; Self, S.G. Tuberculosis vaccines and prevention of infection. Microbiol. Mol. Biol. Rev. 2014, 78, 650–671. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, J.; Monies, R.; Gavier-Widen, M.; Rule, B. Role of infected, non-diseased badgers in the pathogenesis of tuberculosis in the badger. Vet. Rec. 1998, 142, 710–714. [Google Scholar] [CrossRef]
- Donner, A.; Klar, N. Pitfalls of and Controversies in Cluster Randomization Trials. Am. J. Public Health 2004, 94, 416–422. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Time Block | Treatment Set | Odds of Testing Positive | Odds Ratios | ||
|---|---|---|---|---|---|
| Control Start | High Susceptibility Start | High Contact Start | |||
| Study start (September–November 2014) | Control | 0.83 (22) | - | - | - |
| High Susceptibility | 0.18 (26) | 4.58 (1.18–17.79, p = 0.03) | - | - | |
| High Contact | 0.45 (29) | 1.85 (0.59–5.85, p = 0.29) | 2.48 (0.66–9.31, p = 0.18) | - | |
| Study end (July–September 2016) | Control | 4.5 (11) | 5.4 (0.94–30.98, p = 0.058) | - | - |
| High Susceptibility | 0.27 (14) | - | 1.5 (0.28–7.91, p = 0.63) | - | |
| High Contact | 0.45 (29) | - | - | 1.0 (0.33–3.04, p = 1) | |
| Variable | Category | HR * | 95% Confidence Interval | Wald Test p-Value for Variable |
|---|---|---|---|---|
| Vaccinated as a pup | No | 0.305 | ||
| Yes | 0.82 | 0.56–1.20 | ||
| Vaccinated as an adult | No | 0.180 | ||
| Yes | 0.63 | 0.32–1.24 | ||
| Dominance | No | 0.622 | ||
| Yes | 0.92 | 0.66–1.28 | ||
| Sex | F | 0.095 | ||
| M | 1.26 | 0.96–1.66 | ||
| Age | 0–6 months | 0.020 | ||
| 6–12 months | 0.55 | 0.37–0.82 | ||
| 12–24 months | 0.69 | 0.48–1.00 | ||
| >24 months | 0.87 | 0.61–1.24 | ||
| Previous history of TB within social group | No | <0.0001 | ||
| Yes | 2.56 | 1.95–3.37 | ||
| Treatment Set | Control | 0.141 | ||
| High Susceptibility | 0.79 | 0.56–1.13 | ||
| High Contact | 1.14 | 0.85–1.53 |
| Variable | Category First Period (0–180 Days) | Second Period (181–760 Days) | |||||
|---|---|---|---|---|---|---|---|
| HR * | 95% Confidence Interval | p-Value | HR * | 95% Confidence Interval | p-Value | ||
| Sex | F | ||||||
| M | 0.48 | 0.25–0.93 | 0.030 | 1.96 | 1.35–2.84 | <0.001 | |
| Age | 0–6 months | ||||||
| 6–12 months | 0.49 | 0.21–1.13 | 0.094 | 0.67 | 0.39–1.15 | 0.149 | |
| 12–24 months | 0.22 | 0.07–0.76 | 0.016 | 0.94 | 0.59–1.50 | 0.798 | |
| >24 months | 0.38 | 0.15–0.96 | 0.040 | 1.13 | 0.66–1.94 | 0.646 | |
| Previous history of TB within social group | No | ||||||
| Yes | 98.75 | 21.35–456.84 | <0.0001 | 2.14 | 1.18–3.88 | 0.012 | |
| Treatment Set | Control | ||||||
| High Susceptibility | 2.70 | 0.99–7.35 | 0.053 | 0.72 | 0.44–1.18 | 0.188 | |
| High Contact | 3.34 | 1.21–9.24 | 0.020 | 0.50 | 0.29–0.88 | 0.017 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patterson, S.J.; Clutton-Brock, T.H.; Pfeiffer, D.U.; Drewe, J.A. Trait-Based Vaccination of Individual Meerkats (Suricata suricatta) against Tuberculosis Provides Evidence to Support Targeted Disease Control. Animals 2022, 12, 192. https://doi.org/10.3390/ani12020192
Patterson SJ, Clutton-Brock TH, Pfeiffer DU, Drewe JA. Trait-Based Vaccination of Individual Meerkats (Suricata suricatta) against Tuberculosis Provides Evidence to Support Targeted Disease Control. Animals. 2022; 12(2):192. https://doi.org/10.3390/ani12020192
Chicago/Turabian StylePatterson, Stuart J., Tim H. Clutton-Brock, Dirk U. Pfeiffer, and Julian A. Drewe. 2022. "Trait-Based Vaccination of Individual Meerkats (Suricata suricatta) against Tuberculosis Provides Evidence to Support Targeted Disease Control" Animals 12, no. 2: 192. https://doi.org/10.3390/ani12020192
APA StylePatterson, S. J., Clutton-Brock, T. H., Pfeiffer, D. U., & Drewe, J. A. (2022). Trait-Based Vaccination of Individual Meerkats (Suricata suricatta) against Tuberculosis Provides Evidence to Support Targeted Disease Control. Animals, 12(2), 192. https://doi.org/10.3390/ani12020192