


Trends in Reproductive Indicators of Green and Hawksbill Sea Turtles over a 30-Year Monitoring Period in the Southern Gulf of Mexico and Their Conservation Implications

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Data Preprocessing
2.3. Trends of Nesting Abundance
2.4. Carapace Length Distributions
2.5. Clutch Size, Hatching, and Emergence Success
2.6. Statistical Analysis
3. Results
3.1. Trends in Nest Abundance
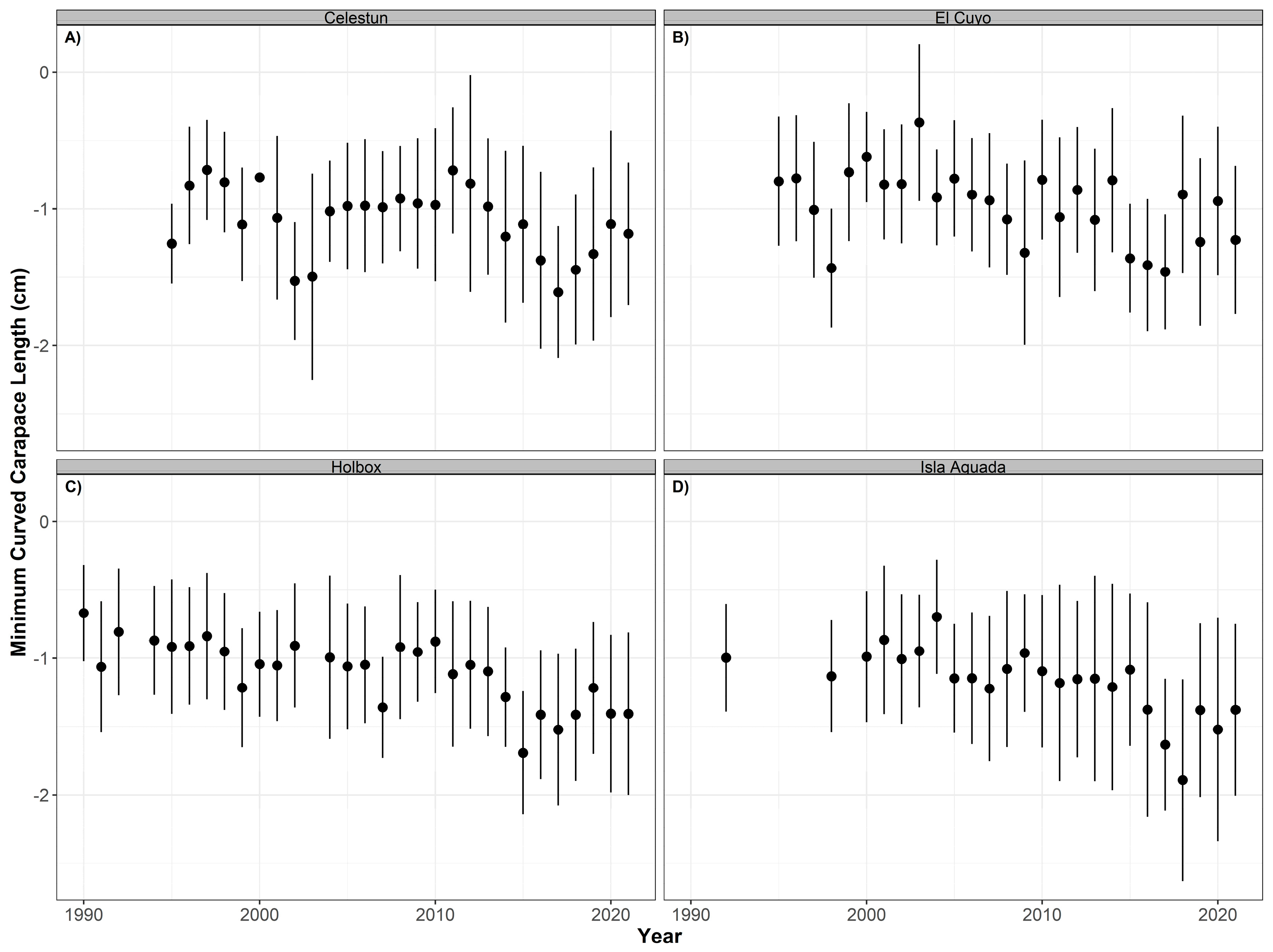
3.2. Carapace Length Distributions
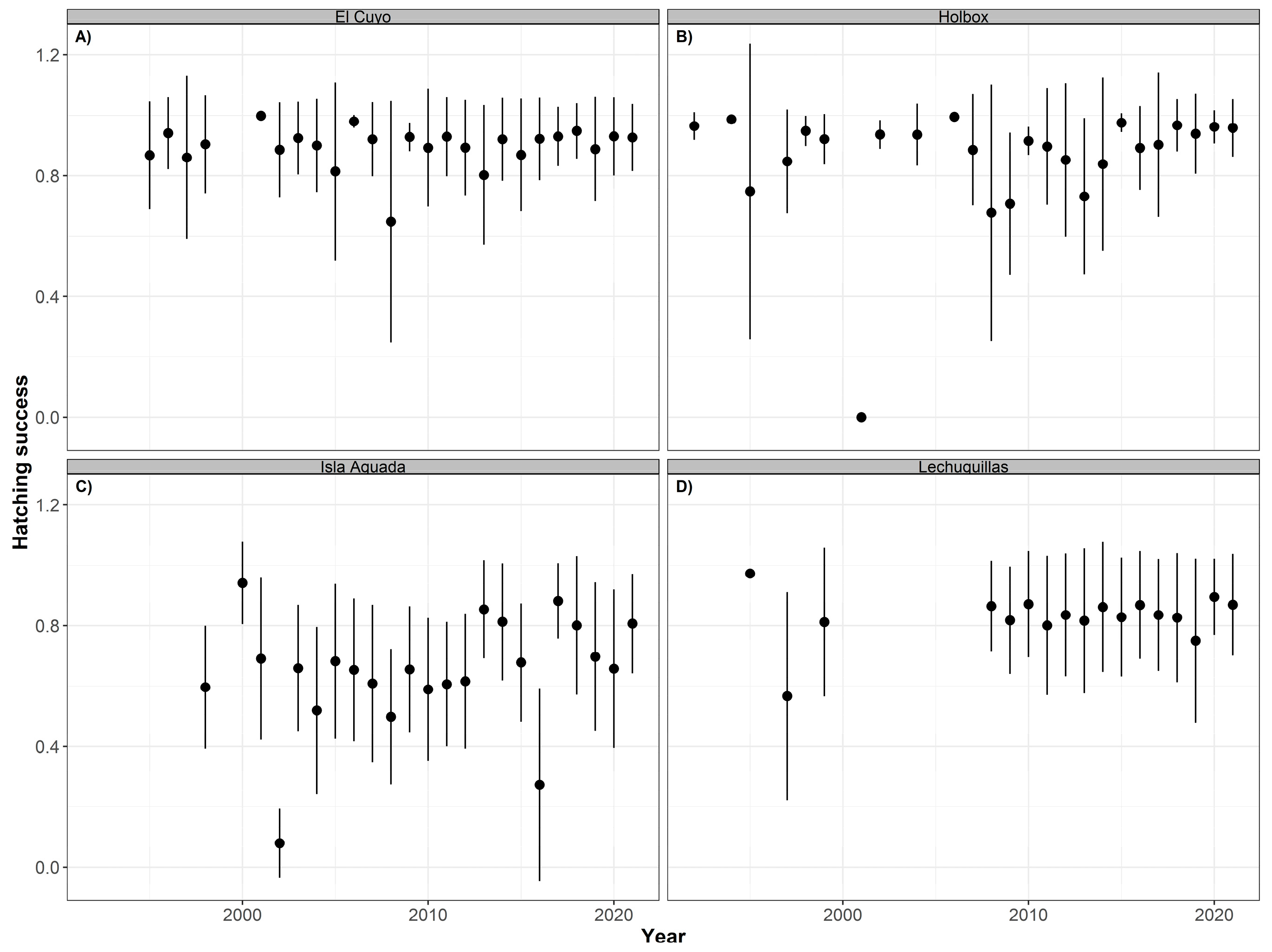
3.3. Temporal Distribution of Clutch Size, Hatching, and Emergence Success
4. Discussion
4.1. Trends in Nest Abundance
4.2. Carapace Length Distribution through Time
4.3. Clutch Size, Hatching, and Emergence Success
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bjorndal, K.A.; Bolten, A.B. From ghosts to key species: Restoring sea turtle populations to fulfill their ecological roles. Mar. Turt. Newsl. 2003, 100, 16–21. [Google Scholar]
- Magurran, A.E.; Baillie, S.R.; Buckland, S.T.; Dick, J.M.; Elston, D.A.; Scott, E.M.; Smith, R.I.; Somerfield, P.J.; Watt, A.D. Long-term datasets in biodiversity research and monitoring: Assessing change in ecological communities through time. Trends Ecol. Evol. 2010, 25, 574–582. [Google Scholar] [CrossRef]
- Mazaris, A.D.; Schofield, G.; Gkazinou, C.; Almpanidou, V.; Hays, G.C. Global sea turtle conservation successes. Sci. Adv. 2017, 3, e1600730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, K.J.; Birks, H.J.B. What is natural? The need for a long-term perspective in biodiversity conservation. Science 2006, 314, 1261–1265. [Google Scholar] [CrossRef] [Green Version]
- Ceriani, S.A.; Casale, P.; Brost, M.; Leone, E.H.; Witherington, B.E. Conservation implications of sea turtle nesting trends: Elusive recovery of a globally important loggerhead population. Ecosphere 2019, 10, e02936. [Google Scholar] [CrossRef] [Green Version]
- Piacenza, S.E.; Balazs, G.H.; Hargrove, S.K.; Richards, P.M.; Heppell, S.S. Trends and variability in demographic indicators of a recovering population of green sea turtles Chelonia mydas. Endanger. Species Res. 2016, 31, 103–117. [Google Scholar] [CrossRef] [Green Version]
- Chaloupka, M.; Bjorndal, K.A.; Balazs, G.H.; Bolten, A.B.; Ehrhart, L.M.; Limpus, C.J.; Suganuma, H.; Troëng, S.; Yamaguchi, M. Encouraging outlook for recovery of a once severely exploited marine megaherbivore. Glob. Ecol. Biogeogr. 2008, 17, 297–304. [Google Scholar] [CrossRef]
- Margaritoulis, D. Nesting Activity and Reproductive Output of Loggerhead Sea Turtles, Caretta caretta, over 19 Seasons (1984-2002) at Laganas Bay, Zakynthos, Greece: The Largest Rookery in the Mediterranean. Chelonian Conserv. Biol. 2005, 4, 916–929. [Google Scholar]
- Le Gouvello, D.Z.M.; Girondot, M.; Bachoo, S.; Nel, R. The good and bad news of long-term monitoring: An increase in abundance but decreased body size suggests reduced potential fitness in nesting sea turtles. Mar. Biol. 2020, 167, 112. [Google Scholar] [CrossRef]
- García-Cruz, M.A.; Lampo, M.; Peñaloza, C.L.; Kendall, W.L.; Solé, G.; Rodríguez-Clark, K.M. Population trends and survival of nesting green sea turtles Chelonia mydas on Aves Island, Venezuela. Endanger. Species Res. 2016, 29, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Castañeda, R.; Salum-Fares, A.; Defeo, O. Reproductive patterns of the hawksbill turtle Eretmochelys imbricata in sandy beaches of the Yucatan Peninsula. J. Mar. Biol. Assoc. UK 2007, 87, 815–824. [Google Scholar] [CrossRef]
- Wallace, B.P.; DiMatteo, A.D.; Hurley, B.J.; Finkbeiner, E.M.; Bolten, A.B.; Chaloupka, M.Y.; Hutchinson, B.J.; Abreu-Grobois, F.A.; Amorocho, D.; Bjorndal, K.A.; et al. Regional Management Units for Marine Turtles: A Novel Framework for Prioritizing Conservation and Research across Multiple Scales. PLoS ONE 2010, 5, e15465. [Google Scholar] [CrossRef]
- CIT. Selección de Playas Índice en la Región CIT y Lineamientos para la Recolección de Datos CIT-CC10-2013-Tec.5; CIT: Danville, VA, USA, 2013. [Google Scholar]
- CIT. Análisis de Datos de Anidación en Playas Índices de la Convención Interamericana Para la Protección y Conservación de las Tortugas Marinas (2009–2013) CIT-CC11-2014-Tec.7; CIT: Danville, VA, USA, 2014. [Google Scholar]
- Peñaflores, C.; Márquez, R. El programa nacional para la conservación de las tortugas marinas: 50 años de historia. In Las Tortugas Marinas en México: Logros y Perspectivas para su Conservación, 1st ed.; Gaona, O., Barragán, A., Eds.; Soluciones Ambientales ITZENI: Mexico City, Mexico, 2016; pp. 159–188. [Google Scholar]
- Eckert, K.; Eckert, A. An Atlas of Sea Turtle Nesting Habitat for the Wider Caribbean Region; WIDECAST Technical Report 19; WIDECAST: Godfrey, IL, USA, 2019. [Google Scholar]
- Kot, C.Y.; Fujioka, E.; DiMatteo, A.D.; Wallace, B.P.; Hutchinson, B.J.; Cleary, J.; Halpin, P.; Mast, R. The State of the World’s Sea Turtles Online Database: Data Provided by the SWOT Team and Hosted on OBIS-SEAMAP. 2021. Available online: http://seamap.env.duke.edu/swot (accessed on 15 November 2022).
- Spotila, J.R. Sea Turtles: A Complete Guide to Their Biology, Behavior, and Conservation; The Johns Hopkins University Press and Oakwood Arts: Baltimore, MD, USA, 2004. [Google Scholar]
- Guzmán-Hernández, V.; del Monte-Luna, P.; López-Castro, M.C.; Uribe-Martínez, A.; Huerta-Rodríguez, P.; Gallegos-Fernández, S.A.; Azanza-Ricardo, J.; Martínez-Portugal, R.C.; Barragán-Zepeda, A.K.; Quintana-Pali, G.P.; et al. Recovery of green turtle populations and their interactions with coastal dune as a baseline for an integral ecological restoration. Acta Bot. Mex. 2022, 129, e1954. [Google Scholar] [CrossRef]
- Guzmán-Hernández, V. Informe Técnico 2017 del Programa de Conservación de Tortugas Marinas en Laguna de Términos, Campeche, México. Contiene información de: 1. CPCTM Isla Aguada y 2. Reseña Estatal; APFFLT/RPCyGM/CONANP/SEMARNAT: Ciudad del Carmen, Campeche, México, 2018. [Google Scholar]
- Lira-Reyes, D.; Nava Serrano, S.; Franco Franco, F.; Salinas Salazar, R.; Mezquita Ruíz, J.; López-Castro, M.C. Informe Final de la Temporada de Anidación 2021 del Programa para la Conservación de la Tortuga Marina en El Cuyo, Yucatán e Isla Holbox, Quintana Roo; Pronatura Península de Yucatán: Mérida, Yucatán, México, 2021. [Google Scholar]
- Scientific Advisory Board. The State of the World’s Sea Turtles (SWOT) Minimum Data Standards for Nesting Beach Monitoring; State of the World’s Sea Turtles: Arlington, VA, USA, 2011. [Google Scholar]
- D.O.F. Norma Oficial Mexicana NOM-162-SEMARNAT-2012. Que Establece las Especificaciones para la Protección, Recuperación y Manejo de las Poblaciones de las Tortugas Marinas en su Hábitat de Anidación. Available online: https://dof.gob.mx/nota_detalle.php?codigo=5286506&fecha=01/02/2013#gsc.tab=0 (accessed on 15 November 2022).
- Wyneken, J. The Anatomy of Sea Turtles; U.S. Department of Commerce NOAA Technical Memorandum NMFS-SEFSC-470; NOAA: Washington, DC, USA, 2001. [Google Scholar]
- Miller, J.D. Determining clutch size and hatching success. In Research and Management Techniques for the Conservation of Sea Turtles, IUCN/SSC Marine Turtle Specialist Group Publication No. 4.; Eckert, K.L., Bjorndal, K.A., Abreu-Grobois, F.A., Donelly, M., Eds.; Consolidated Graphic Communications: Blanchard, PA, USA, 1999; pp. 124–129. [Google Scholar]
- Chim-Vera, Y.A. Evaluación del Esfuerzo de Monitoreo del éxito de Emergencia en Nidos de Tortuga Carey y Blanca en la Península de Yucatán; Instituto Tecnológico de Conkal: Mérida, Mexico, 2009. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Bjorndal, K.A.; Bolten, A.B. Comparison of straight-line and over-the-curve measurements for growth rates of green turtles, Chelonia mydas. Bull. Mar. Sci. 1989, 45, 189–192. [Google Scholar]
- Gulick, A.G.; Ewen, K.A.; Pollock, C.G.; Hillis-Starr, Z.M. Trends in abundance and reproductive success of the hawksbill turtle nesting population at Buck Island Reef National Monument, St. Croix, US Virgin Islands. Endanger. Species Res. 2022, 48, 191–198. [Google Scholar] [CrossRef]
- Korkmaz, S.; Goksuluk, D.; Zararsiz, G. MVN: An R Package for Assessing Multivariate Normality. R. Journal 2014, 6, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. 2011, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Troëng, S.; Rankin, E. Long-term conservation efforts contribute to positive green turtle Chelonia mydas nesting trend at Tortuguero, Costa Rica. Biol. Conserv. 2005, 121, 111–116. [Google Scholar] [CrossRef]
- Durán-Quesada, A.M.; Sorí, R.; Ordoñez, P.; Gimeno, L. Climate perspectives in the Intra-Americas seas. Atmosphere 2020, 11, 959. [Google Scholar] [CrossRef]
- Antuña-Marrero, J.C.; Otterå, O.H.; Robock, A.; Mesquita, M.D.S. Modelled and observed sea surface temperature trends for the Caribbean and Antilles. Int. J. Climatol. 2016, 36, 1873–1886. [Google Scholar] [CrossRef]
- Butt, N.; Whiting, S.; Dethmers, K. Identifying future sea turtle conservation areas under climate change. Biol. Conserv. 2016, 204, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Heppell, S.S.; Heppell, S.; Read, A.; Crowder, L.B. Effects of fishing on long-lived marine organisms. Mar. Conserv. Biol. Sci. Maint. Sea’s Biodivers. 2005, 9, 211–231. [Google Scholar]
- Uusi-Heikkilä, S. Implications of size-selective fisheries on sexual selection. Evol. Appl. 2020, 13, 1487–1500. [Google Scholar] [CrossRef] [PubMed]
- Avens, L.; Ramirez, M.; Goshe, L.; Clark, J.; Meylan, A.; Teas, W.; Shaver, D.J.; Godfrey, M.H.; Howell, L. Hawksbill sea turtle life-stage durations, somatic growth patterns, and age at maturation. Endanger. Species Res. 2021, 45, 127–145. [Google Scholar] [CrossRef]
- Herrera-Pavón, R.; Arenas-Martínez, A.; Iturbe, I.; Gomez, L.; Zurita, J. More reports of living tag green turtles in Xcacel, Quintana Roo, México. In Proceedings of the Twenty-Seventh Annual Symposium on Sea Turtle Biology and Conservation, Myrtle Beach, SC, USA, 22–28 February 2007; Rees, A., Frick, M., Panagopoulou, A., Williams, K., Eds.; U.S. DEPARTMENT OF COMMERCE: Miami, FL, USA, 2015; p. 261. [Google Scholar]
- Guzmán-Hernández, V.; Cuevas-Flores, E.; García-Alvarado, P.; González-Ruíz, T. Biological Monitoring of Sea Turtle Nesting Beaches: Datasets and Basic Evaluations; Nova Science Publishers, Inc.: New York, NY, USA, 2015. [Google Scholar]
- Hays, G.C.; Taxonera, A.; Renomen, B.; Fairweather, K.; Lopes, A.; Cozens, J.; Laloë, J.-O. Changes in mean body size in an expanding population of a threatened species. Proc. R. Soc. B 2022, 289, 20220696. [Google Scholar] [CrossRef] [PubMed]
- Bjorndal, K.A.; Bolten, A.B.; Chaloupka, M.; Saba, V.S.; Bellini, C.; Marcovaldi, M.A.G.; Santos, A.J.B.; Bortolon, L.F.W.; Meylan, A.B.; Meylan, P.A.; et al. Ecological regime shift drives declining growth rates of sea turtles throughout the West Atlantic. Glob. Chang. Biol. 2017, 23, 4556–4568. [Google Scholar] [CrossRef] [PubMed]
- Bjorndal, K.A.; Chaloupka, M.; Saba, V.S.; Diez, C.E.; Van Dam, R.P.; Krueger, B.H.; Horrocks, J.A.; Santos, A.J.; Bellini, C.; Marcovaldi, M.A.; et al. Somatic growth dynamics of West Atlantic hawksbill sea turtles: A spatio-temporal perspective. Ecosphere 2016, 7, e01279. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, J.A.; Bickford, D. Shrinking body size as an ecological response to climate change. Nat. Clim. Chang. 2011, 1, 401–406. [Google Scholar] [CrossRef]
- Kamel, S.J.; Delcroix, E. Nesting Ecology of the Hawksbill Turtle, Eretmochelys imbricata, in Guadeloupe, French West Indies from 2000–07. J. Herpetol. 2009, 43, 367–376. [Google Scholar] [CrossRef]
- Azanza Ricardo, J.; Ruisanchez Carrasco, Y.; Ibarra, M.E.; Ruiz Urquiola, A.; Castellanos, C.Y.L.; Rìos Tamayo, D. Indicadores del éxito reproductivo de la tortuga verde (Chelonia mydas) en tres playas de la península de guanahacabibes, Pinar del Rio, Cuba. Rev. Investig. Mar. 2006, 27, 69–78. [Google Scholar]
- Vander Zanden, H.B.; Pfaller, J.B.; Reich, K.J.; Pajuelo, M.; Bolten, A.B.; Williams, K.L.; Frick, M.G.; Shamblin, B.M.; Nairn, C.J.; Bjorndal, K.A. Foraging areas differentially affect reproductive output and interpretation of trends in abundance of loggerhead turtles. Mar. Biol. 2014, 161, 585–598. [Google Scholar] [CrossRef]
- Broderick, A.C.; Glen, F.; Godley, B.J.; Hays, G.C. Variation in reproductive output of marine turtles. J. Exp. Mar. Biol. Ecol. 2003, 288, 95–109. [Google Scholar] [CrossRef] [Green Version]
- Cardona, L.; Clusa, M.; Eder, E.; Demetropoulos, A.; Margaritoulis, D.; Rees, A.F.; Hamza, A.A.; Khalil, M.; Levy, Y.; Türkozan, O.; et al. Distribution patterns and foraging ground productivity determine clutch size in Mediterranean loggerhead turtles. Mar. Ecol. Prog. Ser. 2014, 497, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Bruno, J.F. The impact of climate change on the world’s marine ecosystems. Science 2010, 328, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Bates, N.R.; Johnson, R.J. Acceleration of ocean warming, salinification, deoxygenation and acidification in the surface subtropical North Atlantic Ocean. Commun. Earth Environ. 2020, 1, 33. [Google Scholar] [CrossRef]
- Cabral, H.; Fonseca, V.; Sousa, T.; Leal, M.C. Synergistic effects of climate change and marine pollution: An overlooked interaction in coastal and estuarine areas. Int. J. Environ. Res. Public Health 2019, 16, 2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laloë, J.O.; Cozens, J.; Renom, B.; Taxonera, A.; Hays, G.C. Climate change and temperature-linked hatchling mortality at a globally important sea turtle nesting site. Glob. Chang. Biol. 2017, 23, 4231–4922. [Google Scholar] [CrossRef]
- Patrício, A.R.; Hawkes, L.A.; Monsinjon, J.R.; Godley, B.J.; Fuentes, M.M. Climate change and marine turtles: Recent advances and future directions. Endanger. Species Res. 2021, 44, 363–395. [Google Scholar] [CrossRef]
- Booth, D.T.; Freeman, C. Sand and nest temperatures and an estimate of hatchling sex ratio from the Heron Island green turtle (Chelonia mydas) rookery, Southern Great Barrier Reef. Coral Reefs 2006, 25, 629–633. [Google Scholar] [CrossRef]
- Hernández-Cortés, J.A.; Núñez-Lara, E.; Cuevas, E.; Guzmán-Hernández, V. Natural Beach Vegetation Coverage and Type Influence the Nesting Habitat of Hawksbill Turtles (Eretmochelys imbricata) in Campeche, Mexico. Chelonian Conserv. Biol. 2018, 17, 94–103. [Google Scholar] [CrossRef]
- Pronatura Península de Yucatán, A.C. El Proyecto de Cambio Climático de Pronatura Península de Yucatán, A.C.; Pronatura Península de Yucatán A.C.: Mérida, Mexico, 2021. [Google Scholar]
- Flores-Aguirre, C.D.; Díaz-Hernández, V.; Salgado Ugarte, I.H.; Sosa Caballero, L.E.; Méndez de la Cruz, F.R. Feminization tendency of Hawksbill Turtles (Eretmochelys imbricata) in the western Yucatán Peninsula, Mexico. Amphib. Reptile Conserv. 2020, 14, 190–202. [Google Scholar]
- Méndez, D.; Cuevas, E.; Navarro, J.; Blanca, I.; Guzmán-Hernández, G.V. Rastreo satelital de las hembras de tortuga blanca Chelonia mydas y evaluación de sus ámbitos hogareños en la costa norte de la península de Yucatán, México. Rev. Biol. Mar. Oceanogr. 2013, 48, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Cuevas, E.; Abreu-Grobois, F.A.; Guzmán-Hernández, V.; Liceaga-Correa, M.A.; van Dam, R.P. Post-nesting migratory movements of hawksbill turtles Eretmochelys imbricata in waters adjacent to the Yucatan Peninsula, Mexico. Endanger. Species Res. 2008, 10, 123–133. [Google Scholar] [CrossRef]
- Uribe-Martínez, A.; Liceaga-Correa, M.D.L.A.; Cuevas, E. Critical In-Water Habitats for Post-Nesting Sea Turtles from the Southern Gulf of Mexico. J. Mar. Sci. Eng. 2021, 9, 793. [Google Scholar] [CrossRef]
- Poloczanska, E.S.; Limpus, C.J.; Hays, G.C. Vulnerability of marine turtles to climate change. Adv. Mar. Biol. 2009, 56, 151–211. [Google Scholar] [CrossRef]
- Stephenson, M.L. The Effects of Ocean Acidification on Hawksbill Sea Turtles (Eretmochelys imbricata): An Ecosystem Approach. Indian Ocean. Turt. Newsl. 2014, 19, 5–8. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nesting Beach | Hawksbill | Green |
|---|---|---|
| Number of Nests | Number of Nests | |
| (Mean ± S. D.) | (Mean ± S. D.) | |
| (Range) | (Range) | |
| Cayos Arcas | ---No nesting records | 369.76 ± 240.84 |
| (64–956) | ||
| Celestún | 246.22 ± 124.92 | 1.41 ± 3.78 |
| (38–526) | (0–16) | |
| El Cuyo | 405.56 ± 183.49 | 1134.41 ± 1629.78 |
| (67–818) | (31–6679) | |
| Hobox | 514.84 ± 302.11 | 202.50 ± 238.01 |
| (67–1409) | (4–915) | |
| Isla Aguada | 250.41 ± 120 | 1400.31 ± 1747.01 |
| (124–655) | (38–5974) | |
| Lechuguillas | 1.7 ± 1.16 | 2460.29 ± 2888.12 |
| (0–4) | (61–11,044) | |
| San Lorenzo | 85.89 ± 72.52 | No nesting records |
| (19–365) |
| N Nest as Function of s (Season, by = Beach) + Beach (Negative Binomial) | ||||
|---|---|---|---|---|
| Hawksbill Turtle | ||||
| Estimate | Std Error | z Value | Pr (>|t|) | |
| Intercept | 5.444081 | 0.05783 | 94.081 | <2 × 10−16 *** |
| El Cuyo | 0.50269 | 0.08143 | 6.173 | 6.69 × 10−10 *** |
| Holbox | 0.68002 | 0.08138 | 8.356 | <2 × 10−16 *** |
| Isla Aguada | 0.03654 | 0.08172 | 0.447 | 0.655 |
| San Lorenzo | −1.07578 | 0.09144 | −11.764 | <2 × 10−16 *** |
| R-sq (adj) = 0.782 | Deviance explained = 85.2% | REML = 942 | Scale est. = 1 | n = 156 |
| N Nest as Function of s (Season, by = Beach) + Beach (Negative Binomial, Link = Log) | ||||
|---|---|---|---|---|
| Chelonia mydas | ||||
| Estimate | Std Error | t Value | Pr (>|t|) | |
| Cayo Arcas | 5.4049 | 0.2735 | 19.762 | <2 × 10−16 *** |
| El Cuyo | 1.0033 | 0.2995 | 3.350 | 0.000809 *** |
| Holbox | −0.5224 | 0.2999 | −1.742 | 0.081529 |
| Isla Aguada | 1.2739 | 0.2995 | 4.253 | 2.11 × 10− 5 *** |
| Lechuguillas | 1.9379 | 0.3040 | 6.374 | 1.84 × 10−10 *** |
| R-sq. (adj) = 0.651 | Deviance explained = 79.05% | REML = 1011.5 | Scale est. = 1 | n = 141 |
| Nesting Site | Female Size (CCLmin, cm) | Clutch Size (Egg Number) | Hatching Success (%) | Emergence Success (%) | Study Period | |
|---|---|---|---|---|---|---|
| Hawksbill Turtle | ||||||
| Celestún | Mean | 89.15 | 131.34 | 83.36 | 77.68 | 1995–2021 |
| S.D. | 5.20 | 31.87 | 25.14 | 30.39 | ||
| Range | 70–110 | 33–193 | 0–100 | 0–100 | ||
| N | 1298 | 4236 | 4236 | 4236 | ||
| El Cuyo | Mean | 90.01 | 136.69 | 80.98 | 74.24 | 1995–2021 |
| S.D. | 5.10 | 32.84 | 25.92 | 31.75 | ||
| Range | 67–120 | 33–193 | 0–100 | 0–100 | ||
| N | 2734 | 5202 | 5202 | 5202 | ||
| Holbox | Mean | 88.72 | 121.17 | 91.72 | 87.00 | 1990–2021 |
| S.D. | 4.75 | 33.30 | 22.75 | 28.25 | ||
| Range | 67–109 | 33–193 | 0–100 | 0–100 | ||
| N | 1721 | 7436 | 7436 | 7436 | ||
| Isla Aguada | Mean | 89.99 | 128.25 | 56.24 | 40.84 | 1998–2021 |
| S.D. | 5.51 | 32.10 | 30.92 | 34.27 | ||
| Range | 69.6–113 | 34–193 | 0–100 | 0–100 | ||
| N | 1669 | 3270 | 3270 | 3270 | ||
| Lechuguillas | Mean | 104 | 94.5 | 94.09 | 93.08 | 2018 |
| S.D. | 4 | 6.36 | 2.64 | 1.21 | ||
| Range | 100–108 | 90–99 | 92.22–95.95 | 92.22–93.93 | ||
| N | 3 | 2 | 2 | 2 | ||
| San Lorenzo | Mean | 84.63 | 146.05 | 61.59 | 58.37 | 2003–2016 |
| S.D. | 5.15 | 30.48 | 23.33 | 22.84 | ||
| Range | 71.08–98 | 60–193 | 1.18–100 | 0–93.57 | ||
| N | 141 | 220 | 220 | 220 | ||
| Green turtle | ||||||
| Cayos Arcas | Mean | 103.19 | 116.64 | 71.56 | 58.29 | 2002–2014 |
| S.D. | 5.17 | 20.62 | 28.26 | 34.80 | ||
| Range | 76.9–116.34 | 56–153 | 0–100 | 0–100 | ||
| N | 505 | 427 | 427 | 427 | ||
| Celestún | Mean | 105.94 | 116.89 | 79.90 | 76.88 | 1995–2021 |
| S.D. | 8.55 | 19.97 | 17.21 | 21.16 | ||
| Range | 90.76–115.35 | 71–152 | 7.84–100 | 3.92–100 | ||
| N | 6 | 19 | 19 | 19 | ||
| El Cuyo | Mean | 103.72 | 110.27 | 89.40 | 86.43 | 1995–2021 |
| S.D. | 6.25 | 19.81 | 16.71 | 20.80 | ||
| Range | 80.9–124.05 | 54–153 | 0–100 | 0–100 | ||
| N | 3386 | 5476 | 5476 | 5476 | ||
| Holbox | Mean | 103.13 | 113.05 | 91.20 | 88.08 | 1990–2021 |
| S.D. | 5.59 | 19.99 | 17.62 | 21.59 | ||
| Range | 85.85–124.21 | 54–153 | 0–100 | 0–100 | ||
| N | 532 | 1335 | 1335 | 1335 | ||
| Isla Aguada | Mean | 105.66 | 106.52 | 65.31 | 52.26 | 1998–2021 |
| S.D. | 5.38 | 20.05 | 26.01 | 33.77 | ||
| Range | 78.1–124 | 54–153 | 0–100 | 0–100 | ||
| N | 7184 | 12720 | 12720 | 12720 | ||
| Lechuguillas | Mean | 103.95 | 101.81 | 84.09 | 81.19 | 1995–2021 |
| S.D. | 5.45 | 18.54 | 19.81 | 22.65 | ||
| Range | 85–122.94 | 54–154 | 0–100 | 0–100 | ||
| N | 1746 | 26,110 | 26,110 | 26,110 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Castro, M.C.; Cuevas, E.; Guzmán Hernández, V.; Raymundo Sánchez, Á.; Martínez-Portugal, R.C.; Reyes, D.J.L.; Chio, J.Á.B. Trends in Reproductive Indicators of Green and Hawksbill Sea Turtles over a 30-Year Monitoring Period in the Southern Gulf of Mexico and Their Conservation Implications. Animals 2022, 12, 3280. https://doi.org/10.3390/ani12233280
López-Castro MC, Cuevas E, Guzmán Hernández V, Raymundo Sánchez Á, Martínez-Portugal RC, Reyes DJL, Chio JÁB. Trends in Reproductive Indicators of Green and Hawksbill Sea Turtles over a 30-Year Monitoring Period in the Southern Gulf of Mexico and Their Conservation Implications. Animals. 2022; 12(23):3280. https://doi.org/10.3390/ani12233280
Chicago/Turabian StyleLópez-Castro, Melania C., Eduardo Cuevas, Vicente Guzmán Hernández, Ángeles Raymundo Sánchez, Rosa C. Martínez-Portugal, Diana J. Lira Reyes, and Jorge Ángel Berzunza Chio. 2022. "Trends in Reproductive Indicators of Green and Hawksbill Sea Turtles over a 30-Year Monitoring Period in the Southern Gulf of Mexico and Their Conservation Implications" Animals 12, no. 23: 3280. https://doi.org/10.3390/ani12233280
APA StyleLópez-Castro, M. C., Cuevas, E., Guzmán Hernández, V., Raymundo Sánchez, Á., Martínez-Portugal, R. C., Reyes, D. J. L., & Chio, J. Á. B. (2022). Trends in Reproductive Indicators of Green and Hawksbill Sea Turtles over a 30-Year Monitoring Period in the Southern Gulf of Mexico and Their Conservation Implications. Animals, 12(23), 3280. https://doi.org/10.3390/ani12233280