
Size-Mediated Sea Turtle Behavioral Responses at Artificial Habitats in the Northern Gulf of Mexico
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
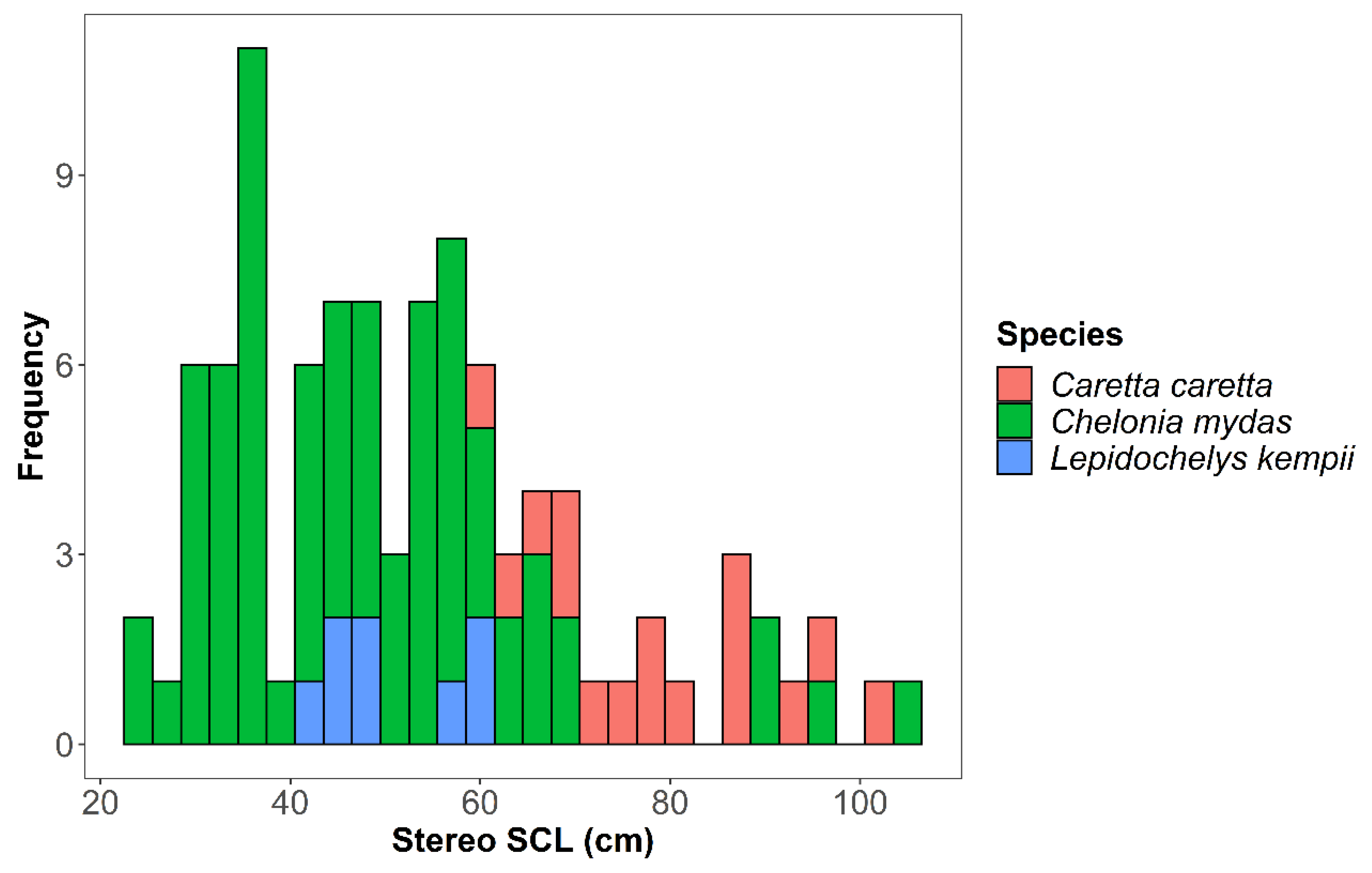
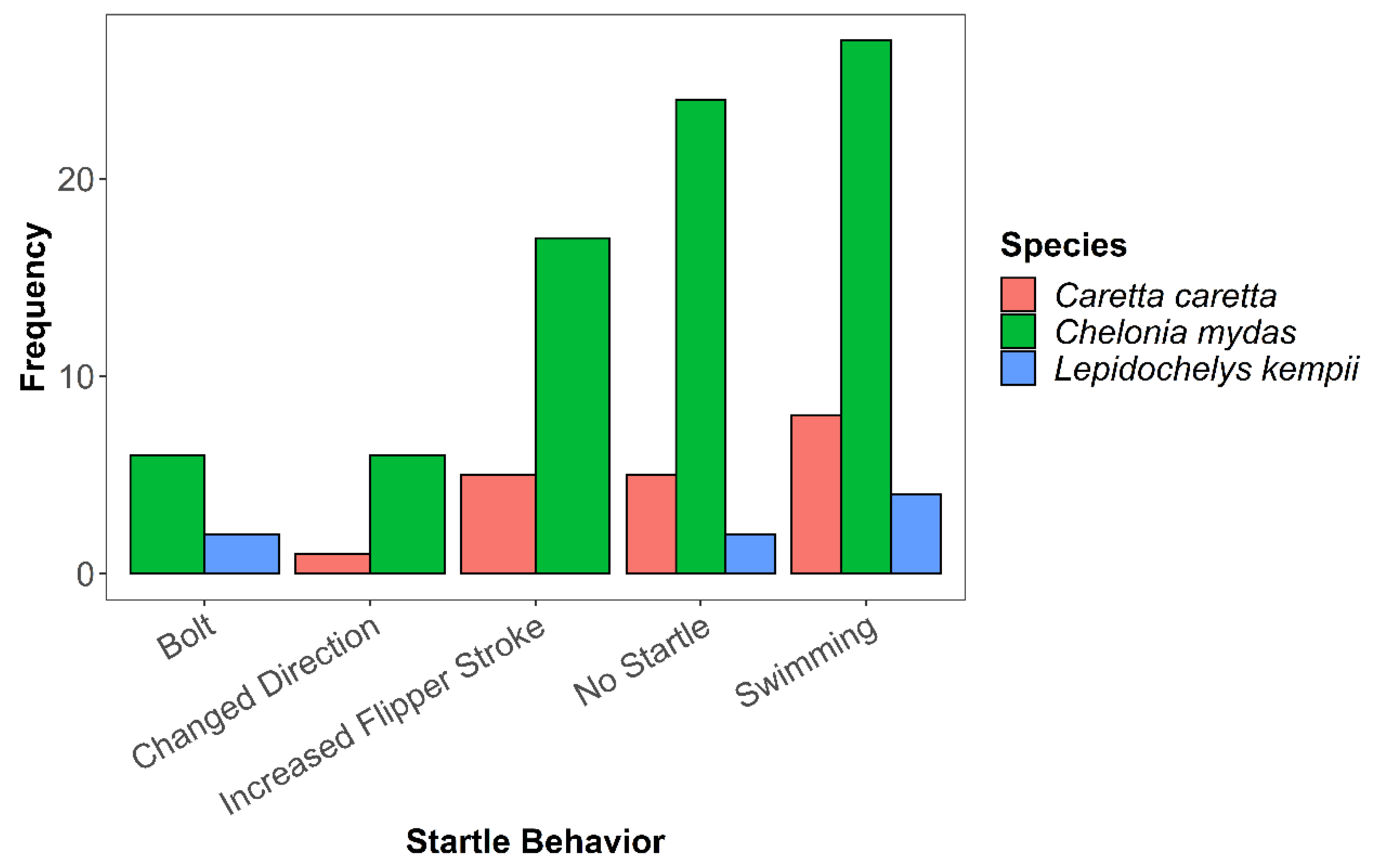
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dammhahn, M. Are Personality Differences in a Small Iteroparous Mammal Maintained by a Life-History Trade-Off? Proc. R. Soc. B 2012, 279, 2645–2651. [Google Scholar] [CrossRef] [Green Version]
- Parkos III, J.J.; Wahl, D.H. Influence of Body Size and Prey Type on the Willingness of Age-0 Fish to Forage under Predation Risk. Trans. Am. Fish. Soc. 2010, 139, 969–975. [Google Scholar] [CrossRef]
- Wikelski, M.; Carrillo, V.; Trillmich, F. Energy Limits to Body Size in a Grazing Reptile, the Galapagos Marine Iguana. Ecology 1997, 78, 2204–2218. [Google Scholar] [CrossRef]
- Schmitt, R.J.; Holbrook, S.J. Gape-Limitation, Foraging Tactics and Prey Size Selectivity of Two Microcarnivorous Species of Fish. Oecologia 1984, 63, 6–12. [Google Scholar] [CrossRef]
- Scharf, F.S.; Juanes, F.; Rountree, R.A. Predator Size-Prey Size Relationships of Marine Fish Predators: Interspecific Variation and Effects of Ontogeny and Body Size on Trophic-Niche Breadth. Mar. Ecol. Prog. Ser. 2000, 208, 229–248. [Google Scholar] [CrossRef]
- Heithaus, M.R.; Frid, A.; Wirsing, A.J.; Dill, L.M.; Fourqurean, J.W.; Burkholder, D.; Thomson, J.; Bejder, L. State-Dependent Risk-Taking by Green Sea Turtles Mediates Top-down Effects of Tiger Shark Intimidation in a Marine Ecosystem. J. Anim. Ecol. 2007, 76, 837–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heithaus, M.R.; Wirsing, A.J.; Thomson, J.A.; Burkholder, D.A. A Review of Lethal and Non-Lethal Effects of Predators on Adult Marine Turtles. J. Exp. Mar. Biol. Ecol. 2008, 356, 43–51. [Google Scholar] [CrossRef]
- Ovadia, O.; Schmitz, O.J. Linking Individuals with Ecosystems: Experimentally Identifying the Relevant Organizational Scale for Predicting Trophic Abundances. Proc. Natl. Acad. Sci. USA 2002, 99, 12927–12931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovadia, O.; Schmitz, O.J. Scaling from Individuals to Food Webs: The Role of Size-Dependent Responses of Prey to Predation Risk. Isr. J. Zool. 2004, 50, 273–297. [Google Scholar] [CrossRef]
- Moll, R.J.; Millspaugh, J.J.; Beringer, J.; Sartwell, J.; He, Z. A New ‘View’ of Ecology and Conservation through Animal-Borne Video Systems. Trends Ecol. Evol. 2007, 22, 660–668. [Google Scholar] [CrossRef]
- Hussey, N.E.; Kessel, S.T.; Aarestrup, K.; Cooke, S.J.; Cowley, P.D.; Fisk, A.T.; Harcourt, R.G.; Holland, K.N.; Iverson, S.J.; Kocik, J.F.; et al. Aquatic Animal Telemetry: A Panoramic Window into the Underwater World. Science 2015, 348, 1255642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shillinger, G.; Bailey, H.; Bograd, S.; Hazen, E.; Hamann, M.; Gaspar, P.; Godley, B.; Wilson, R.; Spotila, J. Tagging through the Stages: Technical and Ecological Challenges in Observing Life Histories through Biologging. Mar. Ecol. Prog. Ser. 2012, 457, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Luschi, P.; Hays, G.C.; Papi, F. A Review of Long-Distance Movements by Marine Turtles, and the Possible Role of Ocean Currents. Oikos 2003, 103, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Smolowitz, R.J.; Patel, S.H.; Haas, H.L.; Miller, S.A. Using a Remotely Operated Vehicle (ROV) to Observe Loggerhead Sea Turtle (Caretta caretta) Behavior on Foraging Grounds off the Mid-Atlantic United States. J. Exp. Mar. Biol. Ecol. 2015, 471, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Hays, G.C.; Mortimer, J.A.; Ierodiaconou, D.; Esteban, N. Use of Long-Distance Migration Patterns of an Endangered Species to Inform Conservation Planning for the World’s Largest Marine Protected Area: Turtle Migration from Chagos Archipelago. Conserv. Biol. 2014, 28, 1636–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schofield, G.; Esteban, N.; Katselidis, K.A.; Hays, G.C. Drones for Research on Sea Turtles and Other Marine Vertebrates—A Review. Biol. Conserv. 2019, 238, 108214. [Google Scholar] [CrossRef]
- Reina, R.D.; Abernathy, K.J.; Marshall, G.J.; Spotila, J.R. Respiratory Frequency, Dive Behaviour and Social Interactions of Leatherback Turtles, Dermochelys coriacea during the Inter-Nesting Interval. J. Exp. Mar. Biol. Ecol. 2005, 316, 1–16. [Google Scholar] [CrossRef]
- Schofield, G.; Katselidis, K.; Dimopoulos, P.; Pantis, J.; Hays, G. Behaviour Analysis of the Loggerhead Sea Turtle Caretta caretta from Direct In-Water Observation. Endanger. Species Res. 2006, 2, 71–79. [Google Scholar] [CrossRef]
- Seminoff, J.; Zárate, P.; Coyne, M.; Foley, D.; Parker, D.; Lyon, B.; Dutton, P. Post-Nesting Migrations of Galápagos Green Turtles Chelonia mydas in Relation to Oceanographic Conditions: Integrating Satellite Telemetry with Remotely Sensed Ocean Data. Endanger. Species. Res. 2008, 4, 57–72. [Google Scholar] [CrossRef]
- Lamont, M.M.; Alday, J.; Alday, C. Interspecific Interactions among Three Species of Sea Turtle Using a Common Resting Area. Ecology 2022, 103, e3861. [Google Scholar] [CrossRef]
- Martínez-Estévez, L.; Steller, D.L.; Zilliacus, K.M.; Cuevas Amador, J.P.; Amador, F.C.; Szuta, D.; Miller, S.D.; Dayton, G.H.; Tershy, B.R.; Croll, D.A. Foraging Ecology of Critically Endangered Eastern Pacific Hawksbill Sea Turtles (Eretmochelys imbricata) in the Gulf of California, Mexico. Mar. Environ. Res. 2022, 174, 105532. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, K.M.; Kuhnz, L.A.; Ruhl, H.A.; Huffard, C.L.; Caress, D.W.; Henthorn, R.G.; Hobson, B.W.; McGill, P.; Smith, K.L. An Evaluation of Deep-Sea Benthic Megafauna Length Measurements Obtained with Laser and Stereo Camera Methods. Deep-Sea Res. 1 2015, 96, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Langlois, T.; Goetze, J.; Bond, T.; Monk, J.; Abesamis, R.A.; Asher, J.; Barrett, N.; Bernard, A.T.F.; Bouchet, P.J.; Birt, M.J.; et al. A Field and Video Annotation Guide for Baited Remote Underwater Stereo-video Surveys of Demersal Fish Assemblages. Methods Ecol. Evol. 2020, 11, 1401–1409. [Google Scholar] [CrossRef]
- Siegfried, T.R.; Fuentes, M.M.P.B.; Ware, M.; Robinson, N.J.; Roberto, E.; Piacenza, J.R.; Piacenza, S.E. Validating the Use of Stereo-video Cameras to Conduct Remote Measurements of Sea Turtles. Ecol. Evol 2021, 11, 8226–8237. [Google Scholar] [CrossRef] [PubMed]
- Harvey, E.; Cappo, M.; Shortis, M.; Robson, S.; Buchanan, J.; Speare, P. The Accuracy and Precision of Underwater Measurements of Length and Maximum Body Depth of Southern Bluefin Tuna (Thunnus Maccoyii) with a Stereo–Video Camera System. Fish. Res. 2003, 63, 315–326. [Google Scholar] [CrossRef]
- Harvey, E.; Fletcher, D.; Shortis, M. Estimation of Reef Fish Length by Divers and by Stereo-Video A FIrst Comparison of the Accuracy and Precision in the Field on Living Fish under Operational Conditions. Fish. Res 2002, 11, 255–265. [Google Scholar] [CrossRef]
- Araujo, G.; Legaspi, C.; Ferber, S.; Murray, R.; Burdett, K.; Grundy, S.; Labaja, J.; Snow, S.; Yaptinchay, A.; Ponzo, A. In-Water Methods Reveal Population Dynamics of a Green Turtle Chelonia mydas Foraging Aggregation in the Philippines. Endanger. Species. Res. 2019, 40, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Goetze, J.S.; Bond, T.; McLean, D.L.; Saunders, B.J.; Langlois, T.J.; Lindfield, S.; Fullwood, L.A.F.; Driessen, D.; Shedrawi, G.; Harvey, E.S. A Field and Video Analysis Guide for Diver Operated Stereo-Video. Methods Ecol. Evol. 2019, 10, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Smulders, F.O.H.; O’Shea, O.R.; Christianen, M.J.A. Animal-Borne Video Reveals Atypical Behaviour in Provisioned Green Turtles: A Global Perspective of a Widespread Tourist Activity. Glob. Ecol. Conserv. 2021, 25, e01417. [Google Scholar] [CrossRef]
- Thomson, J.A.; Heithaus, M.R. Animal-Borne Video Reveals Seasonal Activity Patterns of Green Sea Turtles and the Importance of Accounting for Capture Stress in Short-Term Biologging. J. Exp. Mar. Biol. Ecol. 2014, 450, 15–20. [Google Scholar] [CrossRef]
- Heithaus, M.R.; McLash, J.J.; Frid, A.; Dill, L.M.; Marshall, G.J. Novel Insights into Green Sea Turtle Behaviour Using Animal-Borne Video Cameras. J. Mar. Biol. Assoc. 2002, 82, 1049–1050. [Google Scholar] [CrossRef] [Green Version]
- Hayes, C.T.; Baumbach, D.S.; Juma, D.; Dunbar, S.G. Impacts of Recreational Diving on Hawksbill Sea Turtle (Eretmochelys imbricata) Behaviour in a Marine Protected Area. J. Sustain. Tour. 2017, 25, 79–95. [Google Scholar] [CrossRef]
- Wood, L.D.; Milton, S.L.; Maple, T.L. Foraging Behavior of Wild Hawksbill Turtles (Eretmochelys imbricata) in Palm Beach County, Florida, USA. Chelonian Conserv. Biol. 2017, 16, 70–75. [Google Scholar] [CrossRef]
- Letessier, T.B.; Bouchet, P.J.; Reisser, J.; Meeuwig, J.J. Baited Videography Reveals Remote Foraging and Migration Behaviour of Sea Turtles. Mar. Biodivers. 2015, 45, 609–610. [Google Scholar] [CrossRef]
- Siegfried, T.; Noren, C.; Reimer, J.; Ware, M.; Fuentes, M.M.P.B.; Piacenza, S.E. Insights Into Sea Turtle Population Composition Obtained With Stereo-Video Cameras in Situ Across Nearshore Habitats in the Northeastern Gulf of Mexico. Front. Mar. Sci. 2021, 8, 746500. [Google Scholar] [CrossRef]
- Broadbent, H.; Grasty, S.; Hardy, R.; Lamont, M.; Hart, K.; Lembke, C.; Brizzolara, J.; Murawski, S. West Florida Shelf Pipeline Serves as Sea Turtle Benthic Habitat Based on in Situ Towed Camera Observations. Aquat. Biol. 2019, 29, 17–31. [Google Scholar] [CrossRef]
- Wildermann, N.E.; Gredzens, C.; Avens, L.; Barrios-Garrido, H.A.; Bell, I.; Blumenthal, J.; Bolten, A.B.; McNeill, J.B.; Casale, P.; Domenico, M.D.; et al. Informing Research Priorities for Immature Sea Turtles through Expert Elicitation. Endanger. Species Res. 2018, 37, 55–76. [Google Scholar] [CrossRef] [Green Version]
- Bjorndal, K.A.; Jackson, J. Roles of Sea Turtles in Marine Ecosystems: Reconstructing the Past. In The Biology of Sea Turtles; CRC Press: Boca Raton, FL, USA, 2003; Volume II, p. 455. [Google Scholar]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [Green Version]
- McClenachan, L.; Jackson, J.B.; Newman, M.J. Conservation Implications of Historic Sea Turtle Nesting Beach Loss. Front. Ecol. Environ. 2006, 4, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Wabnitz, C.C.C.; Balazs, G.; Beavers, S.; Bjorndal, K.A.; Bolten, A.B.; Christensen, V.; Hargrove, S.; Pauly, D. Ecosystem Structure and Processes at Kaloko Honokōhau, Focusing on the Role of Herbivores, Including the Green Sea Turtle Chelonia mydas, in Reef Resilience. Mar. Ecol. Prog. Ser. 2010, 420, 27–44. [Google Scholar] [CrossRef]
- Heithaus, M.R.; Alcoverro, T.; Arthur, R.; Burkholder, D.A.; Coates, K.A.; Christianen, M.J.A.; Kelkar, N.; Manuel, S.A.; Wirsing, A.J.; Kenworthy, W.J.; et al. Seagrasses in the Age of Sea Turtle Conservation and Shark Overfishing. Front. Mar. Sci. 2014, 1, 28. [Google Scholar] [CrossRef] [Green Version]
- Wallace, B.P.; Lewison, R.L.; McDonald, S.L.; McDonald, R.K.; Kot, C.Y.; Kelez, S.; Bjorkland, R.K.; Finkbeiner, E.M.; Helmbrecht, S.; Crowder, L.B. Global Patterns of Marine Turtle Bycatch: Global Patterns of Marine Turtle Bycatch. Conserv. Lett. 2010, 3, 131–142. [Google Scholar] [CrossRef]
- Finkbeiner, E.M.; Wallace, B.P.; Moore, J.E.; Lewison, R.L.; Crowder, L.B.; Read, A.J. Cumulative Estimates of Sea Turtle Bycatch and Mortality in USA Fisheries between 1990 and 2007. Biol. Conserv. 2011, 144, 2719–2727. [Google Scholar] [CrossRef]
- Lewison, R.; Crowder, L.; Read, A.; Freeman, S. Understanding Impacts of Fisheries Bycatch on Marine Megafauna. Trends Ecol. Evol. 2004, 19, 598–604. [Google Scholar] [CrossRef]
- Lewison, R.L.; Crowder, L.B.; Wallace, B.P.; Moore, J.E.; Cox, T.; Zydelis, R.; McDonald, S.; DiMatteo, A.; Dunn, D.C.; Kot, C.Y.; et al. Global Patterns of Marine Mammal, Seabird, and Sea Turtle Bycatch Reveal Taxa-Specific and Cumulative Megafauna Hotspots. Proc. Natl. Acad. Sci. USA 2014, 111, 5271–5276. [Google Scholar] [CrossRef] [Green Version]
- Wallace, B.P.; Avens, L.; Braun-McNeill, J.; McClellan, C.M. The Diet Composition of Immature Loggerheads: Insights on Trophic Niche, Growth Rates, and Fisheries Interactions. J. Exp. Mar. Biol. Ecol. 2009, 373, 50–57. [Google Scholar] [CrossRef]
- Cooke, S.J.; Hinch, S.G.; Wikelski, M.; Andrews, R.D.; Kuchel, L.J.; Wolcott, T.G.; Butler, P.J. Biotelemetry: A Mechanistic Approach to Ecology. Trends Ecol. Evol. 2004, 19, 334–343. [Google Scholar] [CrossRef]
- Metz, T.L.; Landry, A.M. An Assessment of Green Turtle (Chelonia mydas) Stocks Along the Texas Coast, with Emphasis on the Lower Laguna Madre. Chelonian Conserv. Biol 2013, 12, 293–302. [Google Scholar] [CrossRef]
- Coleman, A.T.; Pulis, E.E.; Pitchford, J.L.; Crocker, K.; Heaton, A.J.; Carron, A.M.; Hatchett, W.; Shannon, D.; Austin, F.; Dalton, M.; et al. Population Ecology and Rehabilitation of Incidentally Captured Kemp’s Ridley Sea Turtles (Lepidochelys kempii) in the Mississippi Sound, USA. Herpetol. Conserv. Biol. 2016, 12, 253–264. [Google Scholar]
- Cook, M.; Dunch, V.S.; Coleman, A.T. An Interview-Based Approach to Assess Angler Practices and Sea Turtle Captures on Mississippi Fishing Piers. Front. Mar. Sci. 2020, 7, 655. [Google Scholar] [CrossRef]
- Reimer, J.; Siegfried, T.R.; Roberto, E.; Piacenza, S.E. Environmental Context of Nearby Habitat, and Influence on Recreational Bycatch of Sea Turtles at Fishing Piers in the Eastern Gulf of Mexico. unpublished manuscript.
- Chaloupka, M.; Kamezaki, N.; Limpus, C. Is Climate Change Affecting the Population Dynamics of the Endangered Pacific Loggerhead Sea Turtle? J. Exp. Mar. Biol. Ecol. 2008, 356, 136–143. [Google Scholar] [CrossRef]
- Mazaris, A.D.; Schofield, G.; Gkazinou, C.; Almpanidou, V.; Hays, G.C. Global Sea Turtle Conservation Successes. Sci. Adv. 2017, 3, e1600730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putman, N.F.; Hawkins, J.; Gallaway, B.J. Managing Fisheries in a World with More Sea Turtles. Proc. R. Soc. B 2020, 287, 20200220. [Google Scholar] [CrossRef] [PubMed]
- Schofield, G.; Katselidis, K.A.; Pantis, J.D.; Dimopoulos, P.; Hays, G.C. Female–Female Aggression: Structure of Interaction and Outcome in Loggerhead Sea Turtles. Mar. Ecol. Prog. Ser. 2007, 336, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Goetze, J.S.; Januchowski-Hartley, F.A.; Claudet, J.; Langlois, T.J.; Wilson, S.K.; Jupiter, S.D. Fish Wariness Is a More Sensitive Indicator to Changes in Fishing Pressure than Abundance, Length or Biomass. Ecol. Appl. 2017, 27, 1178–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolten, A.B. Techniques for Measuring Sea Turtles. In Research and Management Techniques for the Conservation of Sea Turtles; Eckert, S.A., Bjorndal, K.A., Abreu-Grobois, F.A., Donnelly, M., Eds.; IUCN: Grand, Switzerland, 1999; pp. 110–114. [Google Scholar]
- Griffin, L.P.; Brownscombe, J.W.; Gagné, T.O.; Wilson, A.D.M.; Cooke, S.J.; Danylchuk, A.J. Individual-Level Behavioral Responses of Immature Green Turtles to Snorkeler Disturbance. Oecologia 2017, 183, 909–917. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Soft. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R.; Burnham, K.P. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-95364-9. [Google Scholar]
- Johnson, J.B.; Omland, K.S. Model Selection in Ecology and Evolution. Trends Ecol. Evol. 2004, 19, 101–108. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R package version 1.43.17. 2020. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 10 October 2021).
- Arnold, T.W. Uninformative Parameters and Model Selection Using Akaike’s Information Criterion. J. Wildl. Manag. 2010, 74, 1175–1178. [Google Scholar] [CrossRef]
- Leroux, S.J. On the Prevalence of Uninformative Parameters in Statistical Models Applying Model Selection in Applied Ecology. PLoS ONE 2019, 14, e0206711. [Google Scholar] [CrossRef] [Green Version]
- Bresette, M.; Witherington, B.; Herren, R.; Bagley, D.; Gorham, J.; Traxler, S.; Crady, C.; Hardy, R. Size-Class Partitioning and Herding in a Foraging Group of Green Turtles Chelonia mydas. Endanger. Species Res. 2010, 9, 105–116. [Google Scholar] [CrossRef]
- Márquez Millán, R. Sea Turtles of the World. An Annotated and Illustrated Catalogue of Sea Turtle Species Known to Date. FAO Species Cat. 1990, 11, 1–81. [Google Scholar]
- Turtle Expert Working Group. Assessment Update for the Kemp’s Ridley and Loggerhead Sea Turtle Populations in the Western North Atlantic. NOAA Tech. Memo. NMFS-SEFSC 2000, 444, 115. [Google Scholar]
- Stankowich, T.; Blumstein, D.T. Fear in Animals: A Meta-Analysis and Review of Risk Assessment. Proc. R. Soc. B 2005, 272, 2627–2634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andradi-Brown, D.A.; Gress, E.; Laverick, J.H.; Monfared, M.A.A.; Rogers, A.D.; Exton, D.A. Wariness of Reef Fish to Passive Diver Presence with Varying Dive Gear Type across a Coral Reef Depth Gradient. J. Mar. Biol. Assoc. 2018, 98, 1733–1743. [Google Scholar] [CrossRef]
- Jerome Montague, J. Influence of Water Level, Hunting Pressure and Habitat Type on Crocodile Abundance in the Fly River Drainage, Papua New Guinea. Biol. Conserv. 1983, 26, 309–339. [Google Scholar] [CrossRef]
- Webb, G.; Messel, H. Wariness in Crocodylus Porosus (Reptilia: Crocodilidae). Wildl. Res. 1979, 6, 227–234. [Google Scholar] [CrossRef]
- Pacheco, L.F. Wariness of Caiman Populations and Its Effect on Abundance Estimates. J. Herpetol. 1996, 30, 123–126. [Google Scholar] [CrossRef]
- Lamont, M.M.; Johnson, D. Variation in Species Composition, Size and Fitness of Two Multi-Species Sea Turtle Assemblages Using Different Neritic Habitats. Front. Mar. Sci. 2021, 7, 608740. [Google Scholar] [CrossRef]
- Roberto, E.; Piacenza, S. Obtaining in-Water Morphometrics and Photo Identification of Sea Turtles in Nearshore Habitats Along the Gulf of Mexico via Stereo-Video Cameras; University of West Florida: Pensacola, FL, USA, 2022; manuscript in preparation. [Google Scholar]
- Gotanda, K.M.; Turgeon, K.; Kramer, D.L. Body Size and Reserve Protection Affect Flight Initiation Distance in Parrotfishes. Behav. Ecol. Sociobiol. 2009, 63, 1563–1572. [Google Scholar] [CrossRef]
- Sazima, I.; Grossman, A.; Sazima, C. Hawksbill Turtles Visit Moustached Barbers: Cleaning Symbiosis between Eretmochelys imbricata and the Shrimp Stenopus Hispidus. Biota Neotrop. 2004, 4, 1–6. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior Category | Behavior Sub-Category | Description |
|---|---|---|
| Movement | Swimming forward | Movement through the water while maintaining the same depth in the water column (Videos S13 and S14) |
| Diving | Steadily increasing depth (Video S5) | |
| Ascent | Steadily decreasing depth (Video S1) | |
| Crawling on benthos | Use of flippers against the seafloor to change position (Video S4) | |
| Obligatory | Breathing | Ascent in which the turtle’s head rises above the surface of the water (Videos S2 and S3) |
| Feeding | Includes acts of foraging and visible consumption of organisms (Video S6) | |
| Unique | Intraspecific encounters | Interaction between sea turtles of the same species in which each is aware of the presence of the other (Video S9) |
| Self-cleaning | Active movement to scrape the surface of the sea turtle’s carapace or other body part on another surface (i.e., reef module) (Video S12) | |
| Interspecific encounter with cleaner species | Symbiotic interaction between cleaner species (i.e., fish) and sea turtle to remove epibionts (Videos S15) | |
| Interspecific encounter with >1 species of sea turtle | Two species of sea turtle interacting with each other in proximity (Video S7) | |
| Resting | Resting underneath module | Not actively moving flippers and is remaining below the reef module (Video S11) |
| Resting on top of module | Not actively moving flippers and is remaining atop the reef module (Video S10) |
| Species | Sample Size | MAD Range (m) | Mean ± SD |
|---|---|---|---|
| Green sea turtle | 73 | 0.72–5.99 | 2.10 ± 1.10 |
| Loggerhead sea turtle | 16 | 0.93–3.80 | 2.12 ± 0.99 |
| Kemp’s Ridley sea turtle | 8 | 0.78–3.63 | 2.35 ± 0.99 |
| Model Terms | Model Support | |||||||
|---|---|---|---|---|---|---|---|---|
| Chelonia | Dive Method | Habitat Type | lnSCL (cm) | Location | df | AICc | ΔAICc | Weight |
| Model 1 | - | - | 0.19 (−0.01–0.44) | + | 4 | 38.79 | 0 | 0.48 |
| Model 2 | - | - | - | + | 3 | 39.93 | 1.14 | 0.27 |
| Model 3 | + | - | 0.21 | + | 5 | 40.09 | 1.30 | 0.25 |
| R.I. | 0.25 | - | 0.73 | |||||
| Model Terms | Model Support | |||||||
|---|---|---|---|---|---|---|---|---|
| Caretta | Dive Method | Habitat Type | lnSCL (cm) | Location | df | AICc | ΔAICc | Weight |
| Model 1 | - | - | 1.24 (0.29–2.32) | + | 4 | 20.37 | 0 | 0.38 |
| Model 2 | + | - | - | + | 4 | 20.97 | 0.59 | 0.28 |
| Model 3 | - | - | - | + | 3 | 21.68 | 1.31 | 0.20 |
| Model 4 | + | - | 0.89 | + | 5 | 22.22 | 1.85 | 0.15 |
| R.I. | 0.43 | - | 0.53 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siegfried, T.R.; Reimer, J.; Roberto, E.; Noren, C.; Vidal, A.; Dixon, K.; DuBois, M.; Piacenza, S.E. Size-Mediated Sea Turtle Behavioral Responses at Artificial Habitats in the Northern Gulf of Mexico. Animals 2023, 13, 114. https://doi.org/10.3390/ani13010114
Siegfried TR, Reimer J, Roberto E, Noren C, Vidal A, Dixon K, DuBois M, Piacenza SE. Size-Mediated Sea Turtle Behavioral Responses at Artificial Habitats in the Northern Gulf of Mexico. Animals. 2023; 13(1):114. https://doi.org/10.3390/ani13010114
Chicago/Turabian StyleSiegfried, Tabitha Renee, Jackson Reimer, Emma Roberto, Christopher Noren, Alex Vidal, Kristi Dixon, Morgan DuBois, and Susan E. Piacenza. 2023. "Size-Mediated Sea Turtle Behavioral Responses at Artificial Habitats in the Northern Gulf of Mexico" Animals 13, no. 1: 114. https://doi.org/10.3390/ani13010114
APA StyleSiegfried, T. R., Reimer, J., Roberto, E., Noren, C., Vidal, A., Dixon, K., DuBois, M., & Piacenza, S. E. (2023). Size-Mediated Sea Turtle Behavioral Responses at Artificial Habitats in the Northern Gulf of Mexico. Animals, 13(1), 114. https://doi.org/10.3390/ani13010114