1. Introduction
Shrimp
Litopenaeus vannamei production accounts for around 70% of China’s shrimp farming production [
1,
2]. This species is popular with farmers because of its fast growth rate, adaptability to changes in water environment, and resistance to high-density culture and disease [
3]. With rapidly developing intensive aquaculture, the quantity, variety, and production of aquafeeds are rapidly increasing, and the demand for fish meal (FM) has also increased significantly [
4,
5]. However, global FM production is unable to meet the growing demand for aquaculture, and the continued rise in FM prices and the imbalance between supply and demand in the market have significant negative effects on the development of the aquaculture industry [
6]. Therefore, the search for FM alternatives has become a research hotspot for the global aquaculture industry [
7].
Rice protein meal (RPM), a high-quality edible protein with high bioavailability, is a low antigenic protein that does not cause allergic reactions [
8]. RPM is extracted by separation from processing byproducts such as broken rice, rice germ, rice bran, and rice husk [
9]. Its crude protein content is generally 60% to 68%, and its digestible protein content is 56% or more, making it more bio-effective and easier to absorb and utilize than maize, wheat, and other proteins [
10]. In addition, rice protein hydrolysates contain a variety of physiologically active small molecule peptides with antioxidant and immunomodulatory activities [
11]. RPM is widely sourced, productive, and rich in nutrients [
12].
Some studies replaced FM with RPM for
L. vannamei feed by evaluating the utilization and nutrient digestibility of RPM by shrimp. They found that FM could be replaced with RPM in shrimp feed [
13,
14]. High levels of vegetable protein are harmful to shrimp health and affect the composition of shrimp gut bacteria [
15,
16]. The normal flora of the gut is important for the digestive function and health of shrimp. When the intestinal flora enters dysbiosis, the immune system of the host may be affected. Under normal conditions, the dominant flora colonizing the gut acts as a stable ecological community that repels foreign bacteria and serves as an important biological barrier for the immunity of the shrimp gut. However, the effect of RPM on the intestinal flora and immunity of shrimp has not been studied. Therefore, an evaluation of the effects of RPM on growth, non-specific immunity, and intestinal flora for
L. vannamei was carried out.
4. Discussion
RPM is a high-energy, high-protein feed material with high amino acid content and good palatability [
8]. At present, effects of RPM replacing FM on growth for shrimps have been reported. A study has shown that it is possible to replace 60% of FM (the base FM content: 30%) in juvenile
L. vannamei (initial weight: 0.65 ± 0.01 g) feed with RPM based on indicators of growth and feed utilization [
13]. In addition, Oujifard [
14] revealed that RPM might be a potential candidate for 50% replacement of FM (the base FM content: 45.7%) in shrimp (Initial weight: 6.99 ± 0.08 g) feed. However, in the present study, RPM replacement for 10% FM (the base FM content: 20%) protein in shrimp (initial weight: 0.54 ± 0.01 g) feed showed no significant effects on WG, SGR, and FCR. When the proportion of RPM replacement increased, the WG of shrimp decreased significantly. This result is consistent with the variation in CP content of the shrimps. The current findings were consistent with the report of Oujifard [
14]. In shrimp feed, a difference in the maximum percentage of RPM replacement was found between the present work and the above studies. The reasons for this discrepancy might be related to the quality of the RPM [
32], the base FM content, the size of the shrimp [
33], and other factors. Nevertheless, all of the above studies indicated that RPM could partially replace FM protein in juvenile
L. vannamei feed.
Digestive enzyme activities reflect the most basic physiological characteristics of an animal’s digestion and its ability to utilize feed nutrients [
34]. The growth of shrimps is related to their digestive enzyme activity. In the present study, replacing small amounts (10%) of FM with RPM increased the AMS and TYS activities. A previous work showed that the enzymatic digestion of rice protein produces a flavor peptide that replaces monosodium glutamate, effectively masking the bitterness and enhancing the viscosity and palatability of the feed [
35]. Therefore, replacing small amounts of FM with RPM could improve digestive enzyme activities, thus ensuring that the shrimp could effectively absorb and digest the nutrients.
Animals grow by increasing the volume of tissue structures and accumulating protein; therefore, protein metabolism is often used as a measure of animal growth. Protein metabolism is mainly carried out in the hepatopancreas of shrimps through transamination and deamination. GPT and GOT are two of the most important transaminases in protein metabolism. GPT mainly plays an amino transfer role in catalyzing the reaction between α-ketoglutarate and aspartic acid to produce glutamic acid and oxaloacetic acid. GOT enables the transfer of amino acids between alanine and glutamate and regulates the synthesis of non-essential amino acids and the proteolytic processes. In the present study, replacing 10% FM with RPM significantly enhanced GPT and GOT activities, indicating that amino acid metabolism was accelerated, protein catabolism was decreased, and anabolism was enhanced. As a result, nitrogen accumulated in the shrimp body. The GPT and GOT activities decreased significantly when the proportion of RPM replacement was increased; the same trend was observed in
Megalobrama amblycephala [
36]. This phenomenon might be attributed to the relatively low lysine content of rice protein, resulting in an imbalance in the amino acid composition of the feed and thus affecting the metabolic homeostasis of amino acids in the liver.
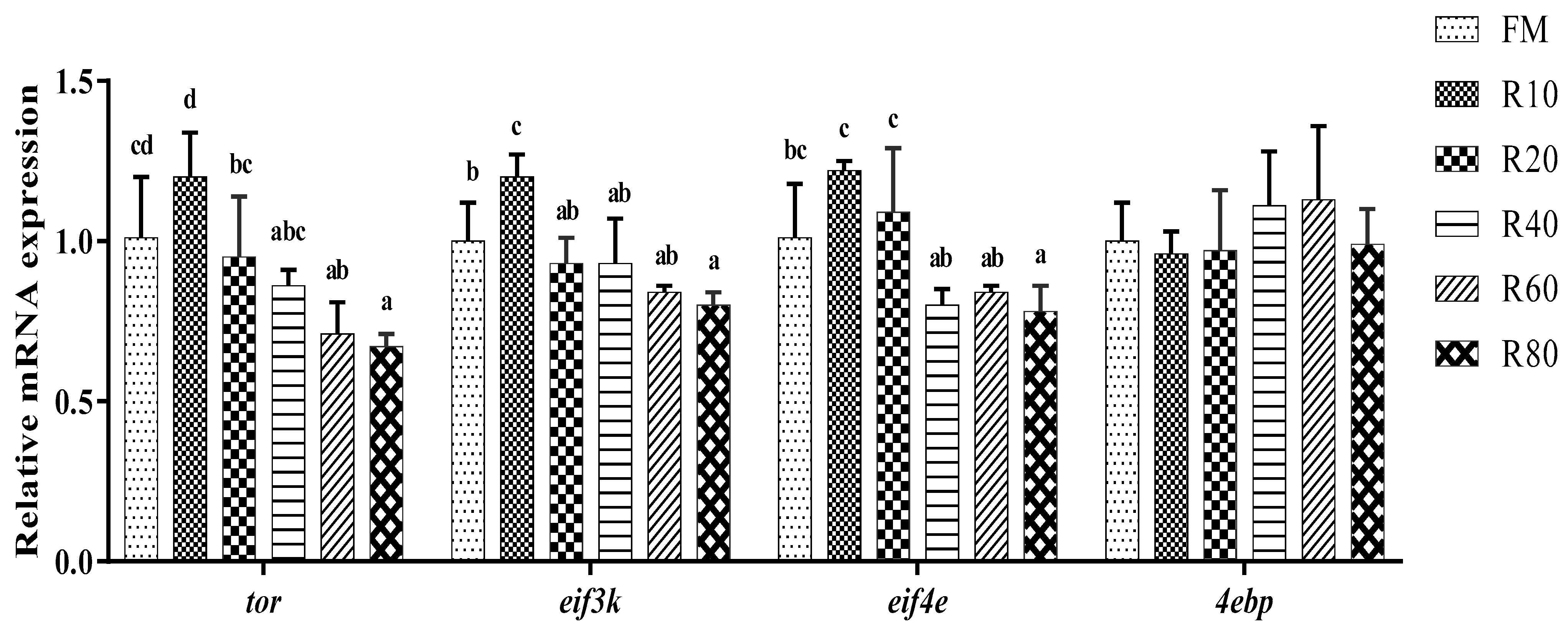
The growth of shrimps is mainly achieved through protein synthesis.
Tor is a highly conserved serine/threonine kinase, and its signaling pathway is vital for organisms to respond effectively to changes in their nutritional environment for regulating growth.
Eif4e, a signaling factor downstream of the mammalian target of rapamycin (mTOR) signaling pathway, receives signals from TOR proteins through eIF4E-BP (
eif4e binding protein) and regulates intracellular protein synthesis [
37].
Eif3k acts as a structural protein and is involved in protein interactions [
38]. In this study, the replacement of small amounts (10%) of FM with RPM increased
tor,
eif3k, and
eif4e mRNA expression levels, indicating that RPM activated the mTOR pathway. An activated mTOR inhibits
4ebp1 phosphorylation, leading to the release of
4ebp1 from
eif4e, reduced inhibition of
eif4e-initiated translation by
4ebp1, and increased expression of
eif4e and thereby promoting the translation of key proteins associated with cell growth [
39]. Under the present experimental conditions, the high protein content, rapid energy conversion, high digestibility, and good palatability of RPM allowed it to replace small amounts of FM in shrimp feed without significantly reducing the WGR.
SOD and CAT scavenge excess radicals from the body and decrease any oxidative damage to cells [
40]. MDA is a peroxidation product and its concentration indicates the damage to the liver. GSH-Px assists in maintaining normal immune system function and has antioxidant and detoxifying effects [
41]. T-AOC is a composite indicator of antioxidant capacity [
42]. Rice protein has endogenous antioxidant effects and could effectively reduce oxidative stress damage to the body [
43]. In the present study, the replacement of small amounts (10% and 20%) of FM with RPM significantly increased the SOD, CAT, GSH-Px, and T-AOC activities and reduced the MDA content. A previous work found that RPM was decomposed by a protease to form different bioactive peptides, and the scavenging rate of 1,1-diphenyl-2-picrylhydrazyl free radicals reached 54.5% [
44]. In the present research, the replacement of a small amount (10%) of FM with RPM significantly increased the TYS activity and aided the digestion of RPM by shrimp. A study showed that rice proteins could be involved in the Nrf2-ARE signaling pathway, thus regulating antioxidant enzyme expression [
45,
46]. However, Fu’s study [
13] found no significant differences in the SOD activity and MDA content of shrimps fed with partial RPM replacement. We speculate that the processing of rice proteins can influence their physico-chemical and structural properties, as well as their structure, solubility, and hydrolytic capacity [
47,
48,
49]. Therefore, under the present experimental conditions, the replacement of a small amount of FM with RPM significantly increased the antioxidant capacity of the shrimps.
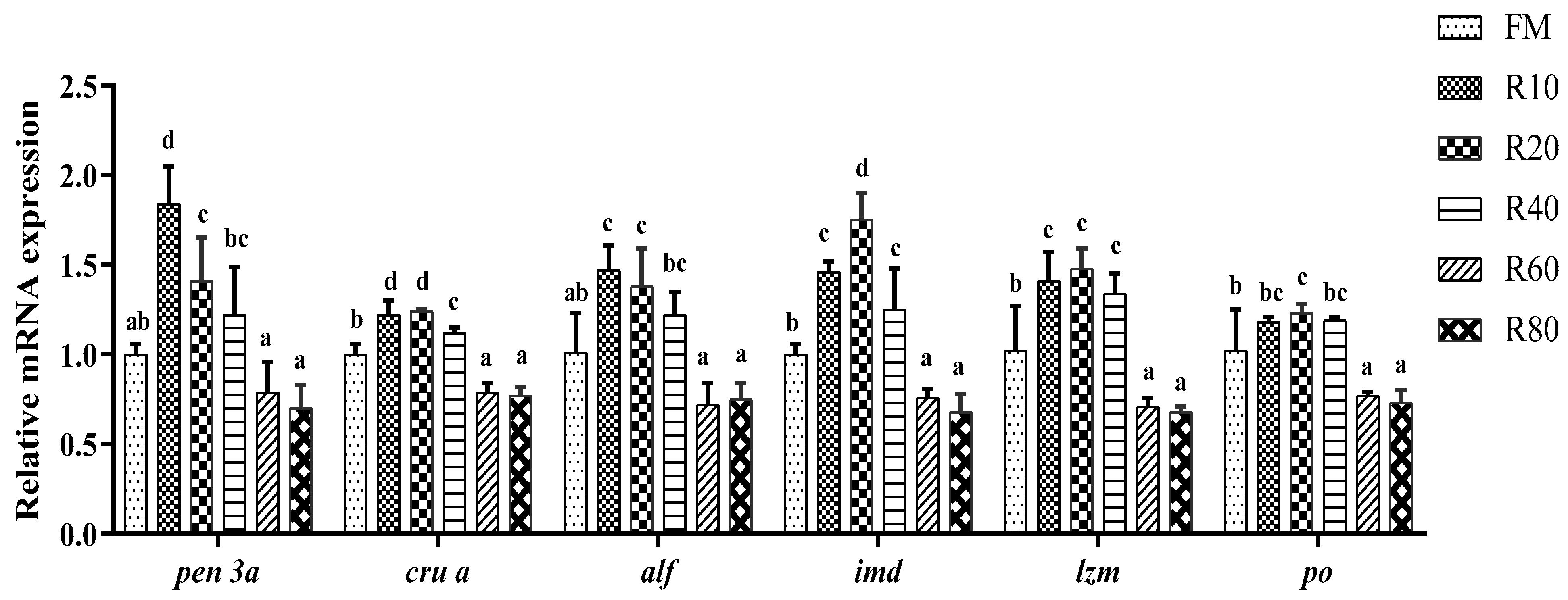
The non-specific immune system is the sole defense of invertebrates against pathogenic invasion [
50]. PO, AKP, and ACP are vital regulatory enzymes and are essential in the non-specific immune response of crustaceans [
51]. LZM has the function of hydrolyzing and digesting invading pathogens and inducing the synthesis and production of related immune factors [
52]. In the present study, replacing 10% of FM with RPM significantly increased PO, AKP, ACP, and LZM activities, in line with Fu’s findings [
13]. Moreover, the replacement of small amounts of FM with RPM increased
lzm and
po mRNA expression. Previously, a study showed that the enzymatic digestion of RPM by trypsin produces a bioactive peptide, oryzatensin, which has immunomodulatory activity [
53].
The intestinal bacterial flora, an important micro-ecosystem, relates to digestion, absorption, and immune function [
54]. Under normal conditions, a balance between the various bacteria in the gut occurs dynamically to maintain the stability of the intestinal environment, thus contributing to the effective suppression of invasion by exogenous pathogenic bacteria and the enhancement of nonspecific immunity [
55]. Richness and diversity indices are important indicators of microbial diversity and complexity, and differences in their values reflect variations in community structure and numbers of species [
56]. Ace and Chao1 indices are commonly used to calculate colony richness, with high values indicating a large number of species in the sample. Meanwhile, the Shannon and Simpson indices are applied to calculate colony diversity, with high values indicating high species diversity [
57]. In the present study, Ace and Chao1 values in the R10 group were significantly higher than those in the control group, and no significant difference in Simpson and Shannon values were observed. This finding indicates that the replacement of a small amount of FM with RPM in the diet increased the species richness of the shrimp gut microbiota without altering its diversity.
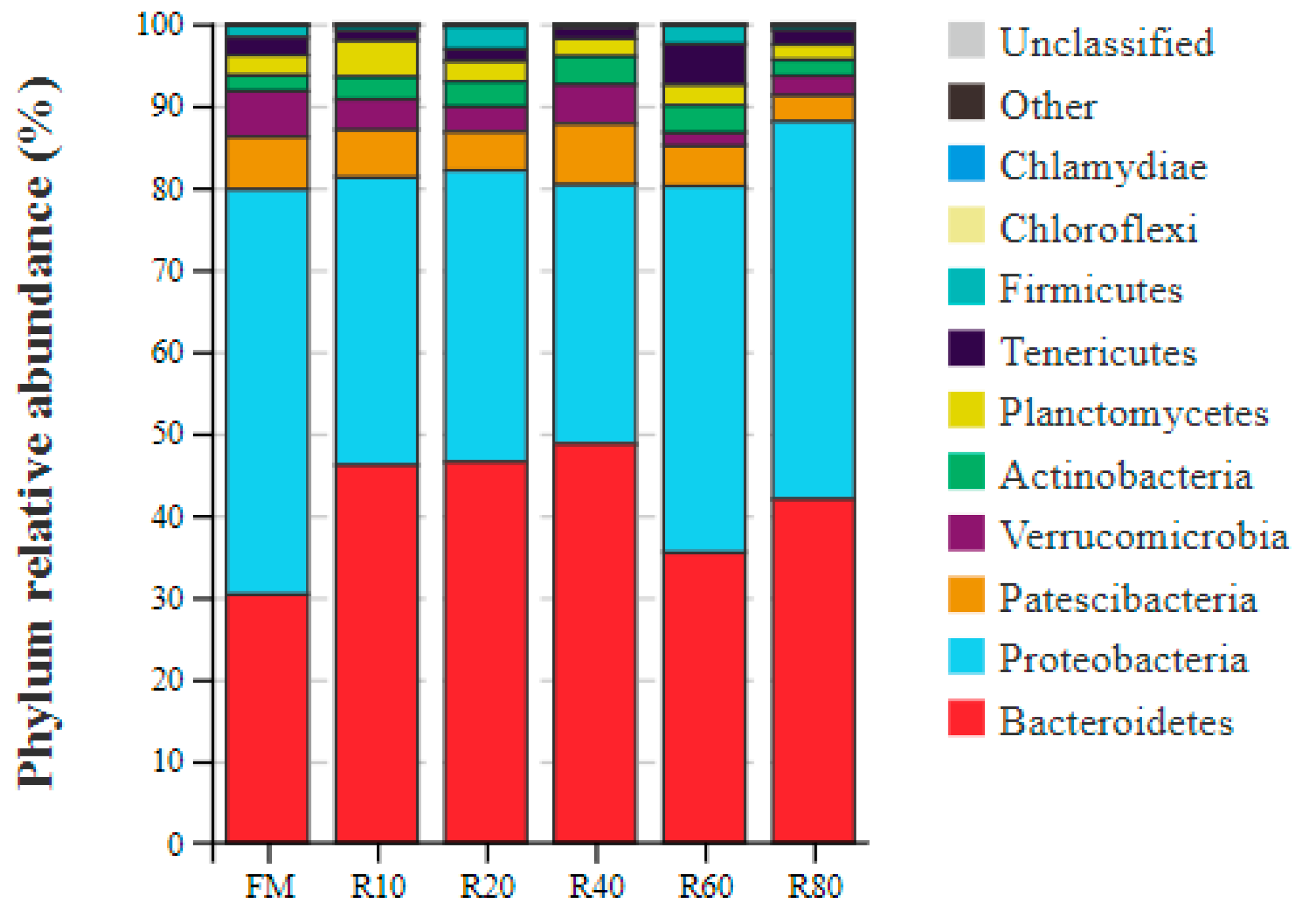
At the phylum level, the main bacterial groups in the gut of shrimp were the phyla Bacteroidetes and Proteobacteria [
16]. Similar conclusions were reached in the present study, with Bacteroidetes and Proteobacteria abundances at over 80%. Bacteroidetes help the host to digest proteins, carbohydrates (especially polysaccharides), and other substances to increase the availability of nutrients; these bacteria are also involved in the metabolism and transport of sugars, provide energy for the host and thus promote its growth [
58]. Proteobacteria are highly correlated with sample spoilage, and their relative proportion in the gut of diseased animals is significantly elevated [
59]. In the present study, the replacement of a small amount (10–40%) of FM with RPM significantly increased Bacteroidetes abundance but reduced that of Proteobacteria. At the genus level,
Vibrio abundance decreased significantly and then increased with increasing RPM replacement amount.
Vibrio is the dominant genus in the sea and one of the main pathogenic bacteria for mariculture animals [
60]. Therefore, the replacement of small amounts (10–40%) of FM with RPM could improve the microbiological composition of the shrimp gut.
The hepatopancreas is the main immune organ of the shrimp, and the innate immunity is an important defense for invertebrates against disease-causing agents. A range of immune factors, such as
penaiedin 3a,
lzm,
alf, and
crustin, play a vital function in humoral immunity and are closely linked to the strength of resistance against infection in
L. vannamei. In invertebrates, the innate immunodeficiency gene IMD encodes an immunodeficiency protein, IMD. The IMD protein transmits the signal of an invading specific foreign body or pathogenic bacteria from outside the cell to inside the cell and causes a series of cascade responses in the cell to form immune effectors (e.g., antimicrobial peptides) [
61]. In this study, the replacement of a small amount of FM with RPM significantly up-regulated the expression of
pen 3a,
cru a,
alf,
imd, and
lzm.
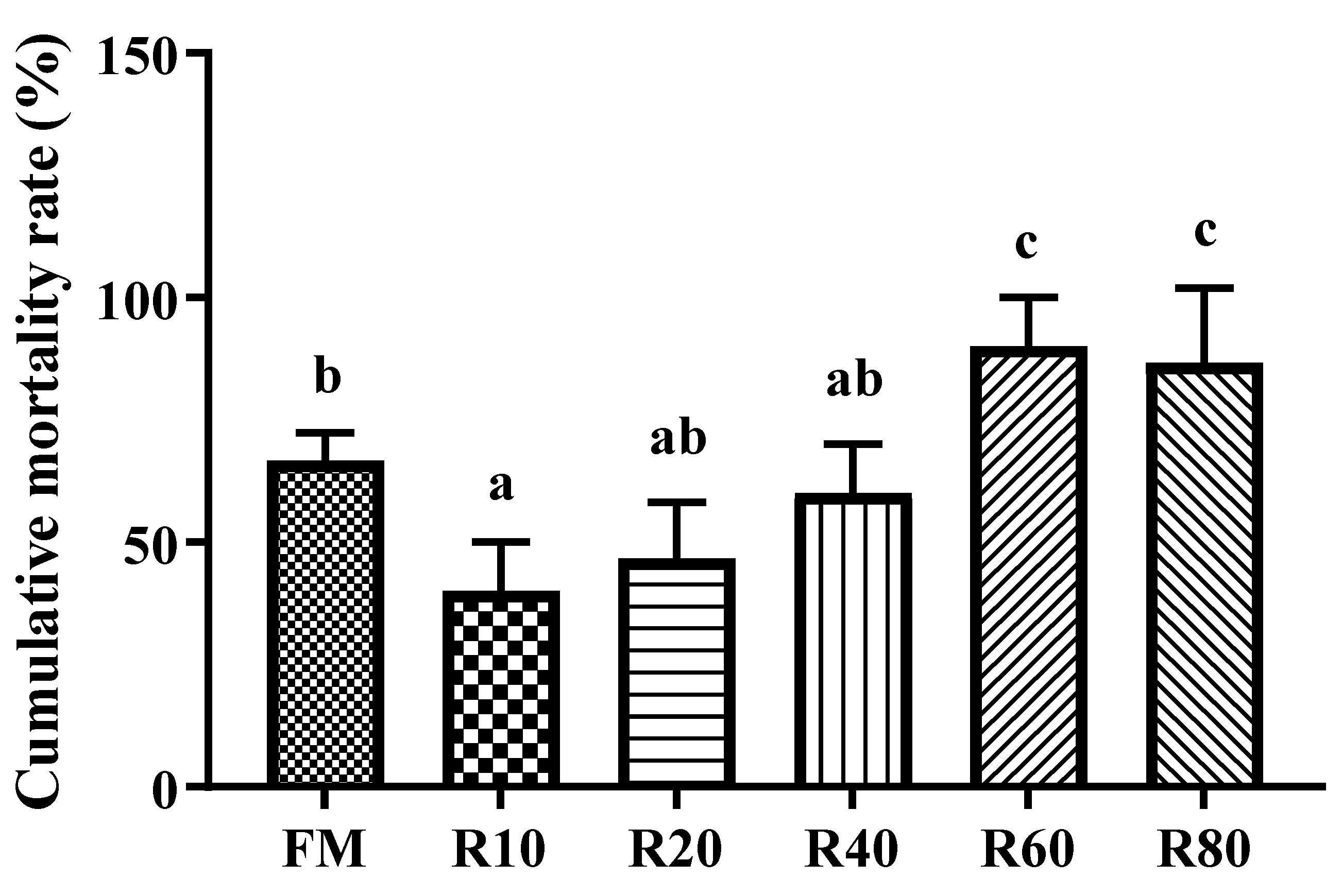
V. parahaemolyticus is a Gram-negative bacterium that is highly pathogenic to shrimps such as
L. vannamei,
Fenneropenaeus chinensis, and
Marsupenaeus japonicus [
62]. In this work, an attack experiment using
V. parahaemolyticus showed the lowest cumulative mortality in the 10% substitution group. Meanwhile, a 60% substitution rate resulted in a significant increase in cumulative mortality in shrimps. Takahashi [
53] showed that the hydrolysis of rice proteins by trypsin yields the immunocompetent nine-band peptide Gly-Tyr-Pro-Met-Tyr-Pro-Leu-Pro-Arg, which enhances the phagocytosis of leukocytes. Therefore, replacing FM with moderate amounts of RPM could significantly improve the immune regulation and resistance to diseases of shrimp.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}