
Biomolecules, Fatty Acids, Meat Quality, and Growth Performance of Slow-Growing Chickens in an Organic Raising System

 ,
,  ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Birds, Experimental Design, and Diets
2.3. Measurements and Chemical Analyses
2.3.1. Growth Performance and Carcass Composition
2.3.2. Sample Collection
2.3.3. Drip Loss Measurement
2.3.4. Cooking Loss Measurement
2.3.5. Warner-Bratzler Shear Force Measurement
2.3.6. Morphological Analysis
2.3.7. Proximate Analysis
2.3.8. Fatty Acid Profile Measurement
2.3.9. Total Collagen Content Measurement
2.3.10. Nucleotide Content Measurement
2.3.11. Cholesterol Measurement
2.4. Synchrotron Radiation-Based Fourier Transform Infrared (SR-FTIR) Spectroscopy
2.4.1. Sample Preparation
2.4.2. SR-FTIR Spectra Measurement
2.4.3. Relative Integral Area for Each Functional Group
2.4.4. Curve Fitting for the Amide I Band
2.5. Statistical Analyses
3. Results and Discussion
3.1. Growth Performance and Carcass Yield
3.2. Physicochemical Properties of Chicken Meat
3.3. Fatty Acid Profile of Chicken Meat
3.4. Changes in Biochemical Profile and Secondary Protein Structure in Breast Meat
3.5. Correlation Loading Plot of FTIR Spectra with the Biochemical Compounds and Quality of Breast Meat from Different Raising Systems
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Srednicka-Tober, D.; Baranski, M.; Seal, C.; Sanderson, R.; Benbrook, C.; Steinshamn, H.; Gromadzka-Ostrowska, J. Composition differences between organic and conventional meat: A systematic literature review and meta-analysis. Br. J. Nutr. 2016, 115, 994–1011. [Google Scholar] [CrossRef] [PubMed]
- Dalle Zotte, A.; Gleeson, E.; Franco, D.; Cullere, M.; Lorenzo, J.M. Proximate composition, amino acid profile, and oxidative stability of slow-growing indigenous chickens compared with commercial broiler chickens. Foods 2020, 9, 546. [Google Scholar] [CrossRef] [PubMed]
- Abbas, T.E.; Ahmed, M.E. Poultry meat quality and welfare as affected by organic production system. AVS 2015, 3, 1–4. [Google Scholar] [CrossRef]
- Miao, Z.H.; Glatz, P.C.; Ru, Y.J. Free-range poultry production-A review. Asian-Australas. J. Anim. Sci. 2005, 18, 113–132. [Google Scholar]
- Grashorn, M.A.; Serini, C.T. Quality of chicken meat from conventional and organic production. Proc. EPC 2006, 10–14. [Google Scholar]
- Husak, R.L.; Sebranek, J.G.; Bregendahl, K. A survey of commercially available broilers marketed as organic, free-range, and conventional broilers for cooked meat yields, meat composition, and relative value. Poult. Sci. 2008, 87, 2367–2376. [Google Scholar] [CrossRef]
- Kucheruk, M.; Midyk, S.; Zasekin, D.; Ushkalov, V.; Kepple, O. Comparison of fatty acid content of organic and traditionally grown broiler chickens. Food Sci. Technol. 2019, 13, 51–57. [Google Scholar] [CrossRef]
- Hovi, M.; Sundrum, A.; Thamsborg, S.M. Animal health and welfare in organic livestock production in Europe: Current state and future challenges. Livest. Prod. Sci. 2003, 80, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Comert, M.; Sayan, Y.; Kırkpınar, F.; Bayraktar, O.H.; Mert, S. Comparison of carcass characteristics, meat quality, and blood parameters of slow and fast grown female broiler chickens raised in organic or conventional production system. Asian-Australas. J. Anim. Sci. 2016, 29, 987–997. [Google Scholar] [CrossRef] [Green Version]
- Dal Bosco, A.; Mattioli, S.; Cartoni Mancinelli, A.; Cotozzolo, E.; Castellini, C. Extensive rearing systems in poultry production: The right chicken for the right farming system. A review of twenty years of scientific research in Perugia University, Italy. Animals 2021, 11, 1281. [Google Scholar] [CrossRef]
- Kubota, S.; Vandee, A.; Keawnakient, P.; Molee, W.; Yongsawatdikul, J.; Molee, A. Effects of the MC4R, CAPN1, and ADSL genes on body weight and purine content in slow-growing chickens. Poult. Sci. 2019, 98, 4327–4337. [Google Scholar] [CrossRef] [PubMed]
- Castellini, C.; Mugnai, C.; Dal Bosco, A. Effect of organic production system on broiler carcass and meat quality. Meat Sci. 2002, 60, 219–225. [Google Scholar] [CrossRef]
- Castellini, C.; Mugnai, C.; Dal Bosco, A. Meat quality of three chicken genotypes reared according to the organic system. Ital. J. Food Saf. 2002, 14, 401–412. [Google Scholar]
- Galvez, F.; Dominguez, R.; Maggiolino, R.A.; Pateiro, K.; Carballo, M.; Lorenzo, J.M. Meat quality of commercial chickens reared in different production systems: Industrial, range and organic. Ann. Anim. Sci. 2020, 20, 263–285. [Google Scholar] [CrossRef] [Green Version]
- Sogut, B.; Inci, H.; Sengul, T. Growth performance and carcass traits of broiler reared in conventional and organic conditions. Asian J. Anim. Vet. Adv. 2011, 6, 992–1000. [Google Scholar] [CrossRef]
- Kucukyilmaz, K.; Bozkurt, M.; Catli, A.U.; Herken, E.N.; Cinar, M.; Bintas, E. Chemical composition, fatty acid profile and colour of broiler meat as affected by organic and conventional rearing systems. S. Afr. J. Anim. Sci. 2012, 42, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Jiang, Z.; Lin, Y.; Zhou, G.; Chen, F.; Zheng, C. Effects of different rearing and feeding methods on meat quality and antioxidative properties in Chinese Yellow male broilers. Br. Poult. Sci. 2011, 52, 352–358. [Google Scholar] [CrossRef]
- Spickett, C.M.; Pitt, A.R. Modification of proteins by reactive lipid oxidation products and biochemical effects of lipoxidation. Essays Biochem. 2020, 64, 19–31. [Google Scholar] [CrossRef]
- Yang, Y.; Wen, J.; Fang, G.Y.; Li, Z.R.; Dong, Z.Y. The effects of raising system on the lipid metabolism and meat quality traits of slow-growing chickens. J. Appl. Anim. Res. 2015, 43, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Luo, C.; Wang, J.; Guo, F. Effects of different raising systems on growth performance, carcass, and meat quality of medium-growing chickens. J. Appl. Anim. Res. 2017, 45, 326–330. [Google Scholar] [CrossRef]
- Mato, A.; Rodriguez Vazquez, R.; Lopez Pedrouso, M.D.; Bravo Lopez, M.D.; Franco Ruiz, S.B.; Zapata Babio, J.C. The first evidence of global meat phosphoproteome changes in response to pre-slaughter stress. BMC 2019, 20, 590–605. [Google Scholar] [CrossRef] [Green Version]
- Baker, M.J.; Trevisan, J.; Bassan, P.; Bhargava, R.; Butler, H.J.; Dorling, K.M.; Fielden, P.R.; Fogarty, S.W.; Fullwood, N.J.; Heys, K.A.; et al. Using Fourier transform IR spectroscopy to analyze biological materials. Nat. Protoc. 2014, 9, 1771–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diem, M.; Meo, M.; Atthaus, C.; Miljkovic, M.; Miller, L.; Lasch, P. Comparison of fourier transform infrared (FTIR) spectra of individual cells acquired using synchrotron and conventional sources. Infrared Phys. Technol. 2004, 45, 331–338. [Google Scholar] [CrossRef]
- Chen, Y.; Zou, C.; Mastalerz, M.; Hu, S.; Gasaway, C.; Tao, X. Applications of micro-Fourier transform infrared spectroscopy (FTIR) in the geological sciences-a review. Int. J. Mol. Sci. 2015, 16, 30223–30250. [Google Scholar] [CrossRef]
- Holman, H.Y.N.; Martin, M.C.; Mckinney, W.R. Tracking chemical changes in a live cell: Biomedical applications of SR–FTIR spectromicroscopy. Spectroscopy 2003, 17, 139–159. [Google Scholar] [CrossRef] [Green Version]
- Poompramun, C.W.; Thumanu, M.K.; Molee, A. The significant influence of Residual Feed Intake (RFI) on flavor precursors and biomolecules in slow-growing Korat chicken meat. Anim. Biosci. 2021, 34, 1684–1694. [Google Scholar] [CrossRef]
- National Bureau of Agricultural Commodity and Food Standards. Thai Agricultural Standard Tas 9000 Part 2-2018, 2nd ed.; ACFS Press: Bangkok, Thailand, 2018.
- Wattanachant, S.; Benjakul, S.; Ledward, D.A. Composition, color, and texture of Thai indigenous and broiler chicken muscles. Poult. Sci. 2004, 83, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.P.; Liu, H.H.; Li, Q.Q.; Wang, Y.; Liu, J.Y.; Hu, J.W.; Yan, X.P.; Gou, H.; Li, L.; Wang, J.W. Gene expression patterns, and protein metabolic and histological analyses for muscle development in Peking duck. Poult. Sci. 2014, 93, 3104–3111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of AOAC, 18th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2010. [Google Scholar]
- Jeon, H.J.; Choe, J.H.; Jung, Y.K.; Kruk, Z.A.; Lim, D.G.; Jo, C.R. Comparison of the chemical composition, textural characteristics, and sensory properties of north and south Korean native chickens and commercial broilers. Food Sci. Anim. Resour. 2010, 30, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Metcalfe, L.D.; Schmitz, A.A.; Pelka, J.R. Rapid preparation of fatty acid esters from lipids for gas chromatographic analysis. Anal. Chem. 1966, 38, 514–519. [Google Scholar] [CrossRef]
- Da Silva, C.M.L.; Spinelli, E.; Rodrigues, S.V. Fast and sensitive collagen quantification by alkaline hydrolysis/hydroxyproline assay. Food Chem. 2015, 173, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Bergman, I.; Loxley, R. Two improved and simplified methods for the spectrophotometric determination of hydroxyproline. Anal. Chem. 1963, 35, 1961–1965. [Google Scholar] [CrossRef]
- Kim, Y.B.; Ku, S.K.; Joo, B.J.; Lee, N.H.; Jang, A. Changs in nucleotide compounds, and chemical and sensory qualities of duck meat during aging at 0 °C. Korean J. Food Sci. Anim. Resour. 2012, 32, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Rowea, A.; Macedob, F.A.F.; Visentainer, J.V.; Souzaa, N.E.; Matsushita, M. Muscle composition and fatty acid profile in lambs fattened in drylot or pasture. Meat Sci. 1999, 51, 283–288. [Google Scholar] [CrossRef]
- Helm, D.; Labischinski, H.; Schallehn, G.; Naumann, D. Classification and identification of bacteria by Fourier-transform infrared spectroscopy. Microbiology 1991, 137, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Liyanage, S.; Dassanayake, R.S.; Bouyanfif, A.; Rajakaruna, E.; Ramalingam, L.; Moustaid-Moussa, N.; Abidi, N. Optimization and validation of cryostat temperature conditions for trans-reflectance mode FTIR microspectroscopic imaging of biological tissues. MethodsX 2017, 4, 118–127. [Google Scholar] [CrossRef]
- Candogan, K.E.; Altuntas, G.; Igci, N. Authentication and quality assessment of meat products by Fourier-transform infrared (FTIR) spectroscopy. Food Eng. Rev. 2020, 13, 66–91. [Google Scholar] [CrossRef]
- Wilkosz, N.; Czaja, M.; Seweryn, S.; Skirlinska-Nosek, K.; Szymonski, M.; Lipiec, E.; Sofinska, K. Molecular spectroscopic markers of abnormal protein aggregation. Molecules 2020, 25, 2498. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.; Mauer, L.J. Fourier transform infrared (FT-IR) spectroscopy: A rapid tool for detection and analysis of foodborne pathogenic bacteria. Appl. Microbiol. Biotechnol. 2010, 2, 1582–1594. [Google Scholar]
- Guo, X.J.; Wang, R.Q. Changes in secondary structure of myofibrillar protein and its relationship with water dynamic changes during storage of battered and deep-fried pork slices. Food Sci. Biotechnol. 2018, 27, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Wasagu, R.S.U.; Abubakar, M.K.; Lawal, M.; Njoku, C.H. Secondary Structures associated with alkaline transition of horse heart ferricytochrome c: An FTIR study. Nig. J. Basic Appl. Sci. 2010, 18, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, S.; Cartoni Mancinelli, A.; Menchetti, L.; Dal Bosco, A.; Angelucci, E.; Castellini, C. How the kinetic behavior of organic chickens affects productive performance and blood and meat oxidative status: A study of six poultry genotypes. Poult. Sci. 2021, 100, 101297. [Google Scholar] [CrossRef]
- Adedeji, O.S.; Amao, S.R.; Oyewumi, S.O.; Alimi, D.H. Growth performance of organically raised broiler chickens at 8 weeks age. Int. J. Appl. Sci. 2014, 3, 140–147. [Google Scholar]
- Santos, A.L.; Sakomura, N.K.; Freitas, E.R.; Fortes, C.M.S.; Carrilho, E.N.M. Comparison of free range broiler chicken strains raised in confined or semi-confined systems. Braz. J. Poult. Sci. 2005, 7, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.H.; Zhang, R.; Gong, A.H.; Gong, D.Q. Effects of different feeding methods on the growth performance of Liyang chicken. China Poult. 2011, 33, 57–58. [Google Scholar]
- Li, L.; Wang, H.H.; Liu, S.Z.; Li, Y.X. Effects of different feeding methods on the production performance and fat deposition of Tibet chicken. Anim. Husb. Feed Sci. 2011, 32, 2–3. [Google Scholar]
- Liu, Z.Y. Effects of different feeding methods on the growth, slaughter performance and meat quality of Guinea fowl. J. Anhui Agric. Univ. 2012, 40, 13397–13417. [Google Scholar]
- Stolze, M.; Piorr, A.; Haring, A.M.; Dabbert, S. Environmental Impacts of Organic Farming in Europe; Haring, A., Ed.; Pages 23–86 in impact of organic farming on the environment and resource use; Universität Hohenheim, Stuttgart-Hohenheim: Ettlingen, Germany, 2000. [Google Scholar]
- Tufarelli, V.; Ragni, M.; Laudadio, V. Feeding forage in poultry: A promising alternative for the future of production systems. Agriculture 2018, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Rothrock, M.J.; Locatelli, A. Importance of farm environment to shape poultry-related microbiomes throughout the farm-to-fork continuum of pasture-raised broiler flocks. Front. Sustain. Food Syst. 2019, 3, 48. [Google Scholar] [CrossRef]
- Fanatico, A.C.; Pillai, P.B.; Emmert, J.L.; Owens, C.M. Meat quality of slow-and fast-growing chicken genotypes fed low-nutrient or standard diets and raised indoors or with outdoor access. Poult. Sci. 2007, 86, 2245–2255. [Google Scholar] [CrossRef] [PubMed]
- Holcman, A.R.; Žlender, V.B.; Stibilj, V. Chemical composition of chicken meat from free-range and extensive indoor rearing. Arch. Geflügelk. 2003, 67, 120–124. [Google Scholar]
- Katemala, S.; Molee, A.; Thumanu, K.; Yongsawatdigul, J. Meat quality and Raman spectroscopic characterization of Korat hybrid chicken obtained from various rearing periods. Poult. Sci. 2021, 100, 1248–1261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Lu, H.; Wang, L.; Yin, M.; Yang, L. Specific expression pattern of IMP metabolism related-genes in chicken muscle between cage and free range conditions. PLoS ONE 2018, 13, e0201736. [Google Scholar] [CrossRef]
- Wilson, D.F.; Matschinsky, F.M. Metabolic homeostasis: Oxidative phosphorylation and the metabolic requirements of higher plants and animals. J. Appl. Physiol. 2018, 125, 1183–1192. [Google Scholar] [CrossRef]
- Peifer, S.; Barduhn, T.; Zimmet, S.; Volmer, D.A.; Heinzle, E.; Schneider, K. Metabolic engineering of the purine biosynthetic pathway in Corynebacterium glutamicum results in increased intracellular pool sizes of IMP and hypoxanthine. Microb. Cell Factories 2012, 11. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.F.; Olesen, J.L.; Hansen, M.; Dossing, S.; Crameri, R.M.; Welling, R.J.; Langberg, H.; Flyvbjerg, A.; Kjaer, M.; Babraj, J.A.; et al. Coordinated collagen and muscle protein synthesis in human patella tendon and quadriceps muscle after exercise. J. Physiol. 2005, 567, 1021–1033. [Google Scholar] [CrossRef]
- Mikulski, D.; Celej, J.; Jankowski, J.; Majewska, T.; Mikulska, M. Growth performance, carcass traits and meat quality of slower-growing and fast-growing chickens raised with and without outdoor access. Asian-Australas. J. Anim. Sci. 2011, 24, 1407–1416. [Google Scholar] [CrossRef]
- Tumova, E.; Teimouri, A. Chicken muscle fibres characteristics and meat quality. A Review. Sci. Agric. Bohem. 2009, 40, 253–258. [Google Scholar]
- Scheffler, T.L.; Park, S.; Gerrard, D.E. Lessons to learn about postmortem metabolism using the AMPKγ3 R200Q mutation in the pig. Meat Sci. 2011, 89, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Kim, C.W.; Yang, M.R.; No, G.R.; Kim, S.W.; Kim, I.S. Pork quality traits according to postmortem pH and temperature in berkshire. Korean J. Food Sci. An. 2016, 36, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowker, B.; Zhuang, H. Relationship between water-holding capacity and protein denaturation in broiler breast meat. Poult. Sci. 2015, 94, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Cavitt, L.C.; Youm, G.W.; Meullenet, J.F.; Owens, C.M.; Xiong, R. Prediction of poultry meat tenderness using Razor Blade Shear, Allo-Kramer Shear and Sarcomere Length. J. Food Sci. 2004, 69, 11–15. [Google Scholar] [CrossRef]
- Vestergaard, M.; Oksbjerg, N.; Henckel, P. Influence of feeding intensity, grazing and finishing feeding on muscle fibre characteristics and meat colour of semitendinosus, longissimus dorsi and supraspinatus muscles of young bulls. Meat Sci. 2000, 54, 177–185. [Google Scholar] [CrossRef]
- Van Wessel, T.; de Haan, A.; van der Laarse, W.J.; Jaspers, R.T. The muscle fiber type-fiber size paradox: Hypertrophy or oxidative metabolism? Eur. J. Appl. Physiol. 2010, 110, 665–694. [Google Scholar] [CrossRef] [Green Version]
- Poltowicz, K.; Doktor, J. Effect of free-range raising on performance, carcass attributes and meat quality of broiler chicken. Anim. Sci. Pap. Rep. 2011, 29, 139–149. [Google Scholar]
- Komprda, T.; Zelenka, J.; Tieffova, P.; Stohandlova, M.; Foltyn, J. Effect of the growth intensity on cholesterol and fatty acids content in broiler chicken tissues. Arch. Fur Geflugelkd. 1999, 63, 36–43. [Google Scholar]
- Dewhurst, R.J.; Nigel, S. Forages, fat, fitness and flavour. Iger Innov. 1998, 2, 36–39. [Google Scholar]
- Boschetti, E.; Bordoni, A.; Meluzzi, A.; Castellini, C.; Dal Bosco, A.; Sirri, F. Fatty acid composition of chicken breast meat is dependent on genotype-related variation of FADS1 and FADS2 gene expression and desaturating activity. Animal 2016, 10, 700–708. [Google Scholar] [CrossRef]
- Bou, R.; Guardiola, F.; Tres, A.; Barroeta, A.; Codony, R. Effect of dietary fish oil, α-tocopheryl acetate, and zinc supplementation on the composition and consumer acceptability of chicken meat. Poult. Sci. 2004, 83, 282–292. [Google Scholar] [CrossRef]
- Cortinas, L.; Villaverde, C.; Galobart, J.; Baucells, M.D.; Codony, R.; Barroeta, A.C. Fatty acid content in chicken thigh and breast as affected by dietary polyunsaturation level. Poult. Sci. 2004, 83, 1155–1164. [Google Scholar] [CrossRef]
- Lopez-Ferrer, S.; Baucells, M.; Barroeta, A.; Grashorn, M. n-3 enrichment of chicken meat. 1. Use of very long-chain fatty acids in chicken diets and their influence on meat quality: Fish oil. Poult. Sci. 2001, 80, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.A. How relevant is the ratio of dietary n-6 to n-3 polyunsaturated fatty acids to cardiovascular disease risk? Evidence from the OPTILIP study. Curr. Opin. Lipidol. 2008, 19, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Silva-Buzanello, R.A.; Schuch, A.F.; Gasparin, A.W.; Torquato, A.S.; Scremin, F.R.; Canan, C.; Soares, A.L. Quality parameters of chicken breast meat affected by carcass scalding conditions. Asian-Australas. J. Anim. Sci. 2019, 32, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, A.; Han, D.H.; Hansen, P.A.; Nolte, L.A.; Host, H.H.; Hickner, R.C.; Holloszy, J.O. Effect of endurance exercise training on muscle glycogen supercompensation in rats. J. Appl. Physiol. 1997, 82, 711–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, J.; Rustad, P.I.; Kolnes, A.J.; Lai, Y.C. The role of skeletal muscle glycogen breakdown for regulation of insulin sensitivity by exercise. Front. Physiol. 2011, 2, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simopoulos, A.P. Omega-6/omega-3 essential fatty acid ratio and chronic diseases. Food Rev. Int. 2004, 20, 77–90. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Item | Starter (d 1 to d 21) | Grower (d 22 to d 42) | Finisher (d 43 to d 84) |
|---|---|---|---|
| Full fat soybean meal (37% CP) | 47.70 | 41.00 | 34.50 |
| Broken rice | 48.50 | 55.65 | 62.35 |
| DL-methionine | 0.25 | 0.10 | 0.10 |
| Salt | 0.35 | 0.35 | 0.35 |
| CaCO3 | 1.40 | 1.30 | 1.20 |
| Monocalcium phosphate (21% P) | 1.30 | 1.10 | 1.00 |
| Premix 1 | 0.50 | 0.50 | 0.50 |
| Calculated nutrients (% unless stated otherwise) | |||
| ME (kcal/kg) | 3175 | 3190 | 3195 |
| Crude protein | 21.00 | 19.00 | 17.00 |
| Crude fat | 8.33 | 7.00 | 7.18 |
| Crude fiber | 2.73 | 2.37 | 2.05 |
| Digestible lysine | 1.21 | 1.08 | 0.95 |
| Digestible methionine | 0.59 | 0.43 | 0.41 |
| Digestible met + cys | 0.93 | 0.73 | 0.69 |
| Digestible threonine | 0.79 | 0.72 | 0.64 |
| Calcium | 1.01 | 0.91 | 0.84 |
| Available phosphorus | 0.45 | 0.38 | 0.35 |
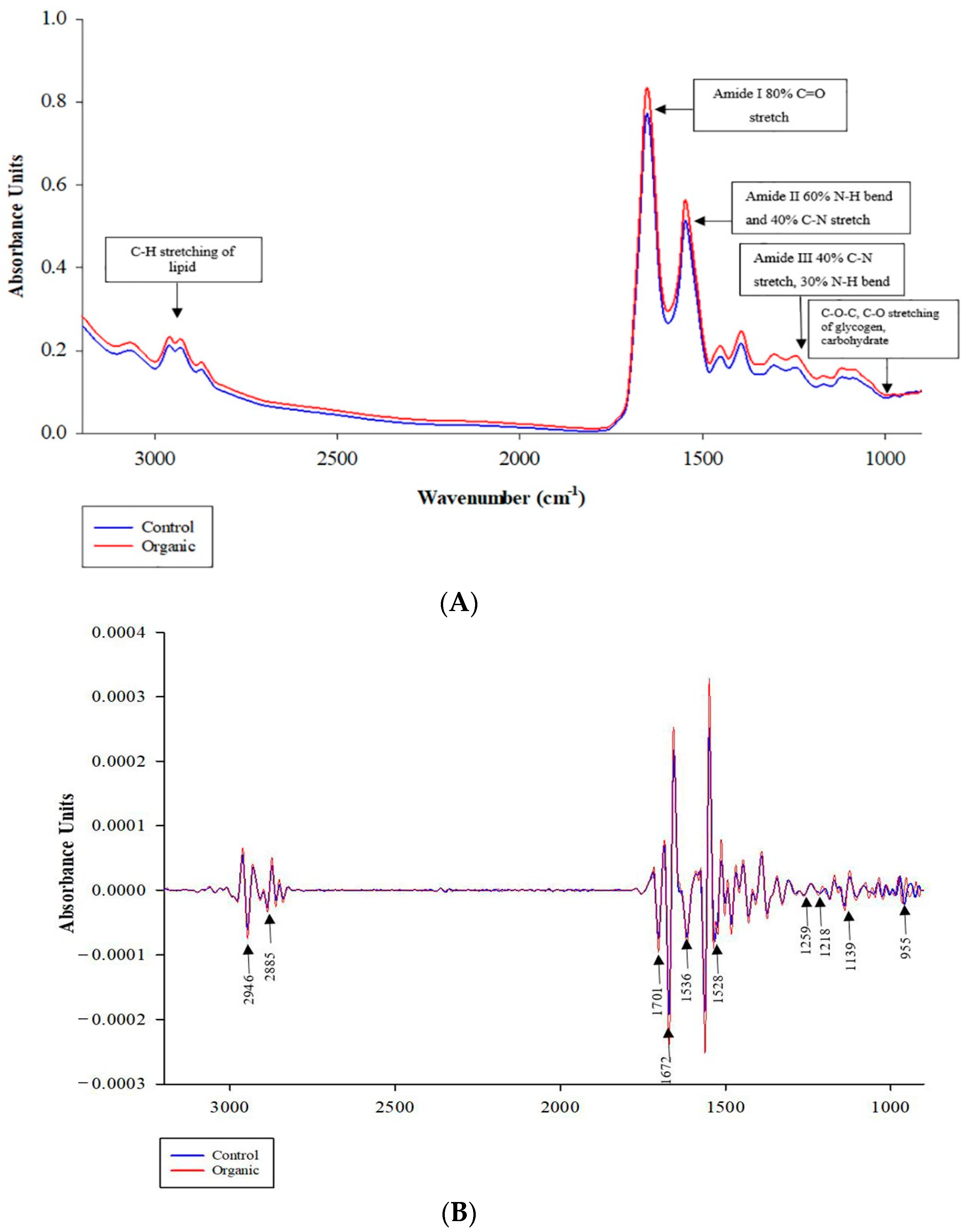
| Wavenumber (cm−1) | Chemical Function | Assignment | References |
|---|---|---|---|
| 2930 to 2910 | CH2 asymmetric stretch | Mainly saturated lipids, proteins | [39] |
| 2970, 2957 to 2953 | CH3 asymmetric stretching | Lipids (mainly), proteins | [40] |
| 2875 to 2870 | CH3 symmetric stretching | Lipids, proteins | |
| 1700 to 1600 | C=O stretching | Amide I band of proteins | [41] |
| 1655, 1650 to 1640 | C=O stretching | Amide I of α-helical structures of proteins | [42,43] |
| 1695 to 1685 | C=O stretching | Antiparallel β-sheet | [41] |
| 1550 to 1520 | C-N stretching + N-H bending coupled in of face | Amide II band of proteins | [43] |
| 1310 to 1240 | C-N stretching + N-H bending coupled in of face | Amide III of proteins | [40,43] |
| 1637 to 1615 | C=O stretching | β-sheet | [40,43,44] |
| 1681 to 1664 1685 to 1675 | C=O stretching | β-turn | [44,45] |
| 1200 to 900 | C-O-C, C-O dominated by ring vibrations of carbohydrates C-O-P, P-O-P | Carbohydrate and Glycogen | [40] |
| Item | CO | OR | p-Value |
|---|---|---|---|
| Final BW (g) | 1480.31 ± 37.94 | 1445.56 ± 42.90 | 0.558 |
| FI (g) | 3634.74 ± 32.22 a | 3074.69 ± 21.37 b | <0.0001 |
| BWG (g) | 1434.60 ± 37.74 | 1399.68 ± 42.80 | 0.554 |
| FCR | 2.54 ± 0.06 a | 2.20 ± 0.07 b | 0.004 |
| Yield (%) | CO | OR | p-Value |
|---|---|---|---|
| Eviscerated carcass 1 | 63.90 ± 0.94 | 63.67 ± 0.84 | 0.941 |
| Pectoralis minor | 3.27 ± 0.12 | 3.05 ± 0.13 | 0.247 |
| Pectoralis major | 8.64 ± 0.20 | 8.49 ± 0.30 | 0.680 |
| Thigh meat | 10.06 ± 0.26 | 10.38 ±0.35 | 0.464 |
| Drumstick meat | 10.02 ± 0.37 | 9.68 ± 0.30 | 0.477 |
| Abdominal fat | 1.47 ± 0.15 a | 1.05 ± 0.11 b | 0.029 |
| Item | Treatment | p-Value | |
|---|---|---|---|
| CO | OR | ||
| Breast meat | |||
| Moisture (%) | 73.74 ± 1.20 | 72.89 ± 0.92 | 0.580 |
| Crude Protein (%) | 23.58 ± 0.12 b | 24.54 ± 0.07 a | <0.0001 |
| Crude fat (%) | 1.99 ± 0.10 | 1.93 ± 0.15 | 0.430 |
| Cholesterol (mg/100 g meat) | 59.04 ± 6.91 | 52.68 ± 4.78 | 0.450 |
| Total collagen (mg/g meat) | 0.85 ± 0.05 b | 1.02 ± 0.05 a | 0.030 |
| IMP (mg/g meat) | 0.14 ± 0.00 | 0.13 ± 0.00 | 0.150 |
| GMP (mg/g meat) | 4.93 ± 0.12 | 5.09 ± 0.09 | 0.150 |
| Thigh meat | |||
| Moisture (%) | 74.04 ± 0.30 | 73.03 ± 0.14 | 0.060 |
| Crude Protein (%) | 13.74 ± 0.04 b | 14.44 ± 0.04 a | <0.0001 |
| Crude fat (%) | 5.64 ± 0.35 | 5.43 ± 0.35 | 0.508 |
| Cholesterol (mg/100 g meat) | 79.89 ± 9.01 | 76.37 ± 5.74 | 0.720 |
| Total collagen (mg/g meat) | 0.73 ± 0.03 b | 1.08 ± 0.06 a | <0.0001 |
| IMP (mg/g meat) | 0.15 ± 0.00 | 0.16 ± 0.00 | 0.220 |
| GMP (mg/g meat) | 3.73 ± 0.22 | 4.18 ± 0.28 | 0.100 |
| Item | CO | OR | p-Value |
|---|---|---|---|
| Ultimate pH | 5.40 ± 0.02 | 5.35 ± 0.02 | 0.079 |
| Drip loss (%) | 11.93 ± 0.66 | 12.27 ± 0.62 | 0.874 |
| Cooking loss (%) | 22.84 ± 0.95 | 23.09 ± 0.55 | 0.570 |
| Shear force (WBS) | 2.17 ± 0.04 b | 2.63 ± 0.10 a | <0.0001 |
| Skin color | |||
| Lightness | 66.96 ± 0.59 | 65.34 ± 2.47 | 0.057 |
| Redness | −0.47 ± 0.19 b | 0.30 ± 0.15 a | 0.004 |
| Yellowness | 7.02 ± 0.37 b | 15.50 ± 0.66 a | <0.0001 |
| Meat color | |||
| Lightness | 52.21 ± 0.65 | 51.37 ± 0.55 | 0.591 |
| Redness | −0.29 ± 0.18 b | 0.35 ± 0.13 a | 0.031 |
| Yellowness | 3.49 ± 0.28 b | 7.27 ± 0.44 a | <0.0001 |
| Muscle diameter (µm) | 22.34 ± 0.51 | 23.88 ± 0.56 | 0.056 |
| Fatty Acid | Breast Meat | Thigh Meat | ||||
|---|---|---|---|---|---|---|
| CO | OR | p-Value | CO | OR | p-Value | |
| C14:0 | 0.90 ± 0.37 | 0.96 ± 0.48 | 0.92 | 0.39 ± 0.01 | 0.38 ± 0.02 | 0.79 |
| C16:0 | 18.27 ± 0.22 | 17.54 ± 0.30 | 0.06 | 16.81 ± 0.46 | 16.99 ± 0.48 | 0.79 |
| C16:1 | 1.53 ± 0.16 | 1.16 ± 0.25 | 0.23 | 2.01 ± 0.24 | 2.09 ± 0.25 | 0.82 |
| C18:0 | 8.28 ± 0.13 | 8.63 ± 0.26 | 0.24 | 7.61 ± 0.36 | 7.42 ± 0.21 | 0.66 |
| C18:1n-9 | 24.56 ± 0.89 | 25.78 ± 1.50 | 0.65 | 26.66 ± 0.75 | 28.67 ± 0.83 | 0.07 |
| C18:2n-6 | 31.09 ± 0.20 | 28.91 ± 0.11 | 0.11 | 37.45 ± 1.11 | 34.90 ± 0.99 | 0.49 |
| C18:3n-6 | 0.13 ± 0.08 | 0.11 ± 0.09 | 0.87 | 0.26 ± 0.01 | 0.22 ± 0.02 | 0.30 |
| C18:3n-3 | 2.12 ± 0.15 | 2.25 ± 0.21 | 0.62 | 3.51 ± 0.16 | 3.77 ± 0.14 | 0.24 |
| C20:2n-6 | 0.30 ± 0.03 | 0.35 ± 0.04 | 0.30 | 0.28 ±0.02 | 0.28 ± 0.04 | 0.90 |
| C20:3n-6 | 0.53 ± 0.04 | 0.63 ± 0.05 | 0.13 | 0.24 ± 0.02 | 0.27 ± 0.01 | 0.33 |
| C20:4n-6 | 9.58 ± 0.86 | 10.50 ± 0.91 | 0.21 | 3.98 ± 0.44 | 3.15 ± 0.45 | 0.20 |
| C20:5n-3 | 0.13 ± 0.05 | 0.16 ± 0.09 | 0.72 | 0.16 ± 0.11 | 0.03 ± 0.02 | 0.29 |
| C22:6n-3 | 1.11 ± 0.07 b | 1.75 ± 0.19 a | 0.01 | 0.41 ± 0.05 | 0.67 ± 0.13 | 0.09 |
| SFA | 27.73 ± 0.50 | 27.13 ± 0.58 | 0.32 | 24.80 ± 0.59 | 24.79 ± 0.52 | 0.40 |
| MUFA | 27.29 ± 1.00 | 28.82 ± 1.62 | 0.29 | 28.89 ± 0.89 | 31.08 ± 0.96 | 0.07 |
| PUFA | 44.98 ± 0.83 | 44.04 ± 0.85 | 0.31 | 46.31 ±1.14 | 44.13 ± 1.36 | 0.27 |
| Total n-6 | 40.79 ± 0.77 | 39.87 ± 0.83 | 0.18 | 41.94 ± 1.02 | 38.59 ± 0.94 | 0.19 |
| Total n-3 | 3.36 ± 0.14 b | 4.17 ± 0.18 a | < 0.001 | 4.09 ± 0.15 b | 5.27 ± 0.18 a | 0.01 |
| n-6/n-3 | 12.14 ± 0.54 a | 9.57 ± 0.40 b | < 0.001 | 10.26 ± 0.21 a | 7.33 ± 0.63 b | 0.02 |
| Biomolecule (Wavenumber) | CO | OR | p-Value |
|---|---|---|---|
| C-H stretching of lipid | 1.13 ± 0.005 a | 1.10 ± 0.005 b | 0.017 |
| Amide I 80% C=O stretch | 11.38 ± 0.007 b | 11.97 ± 0.006 a | <0.0001 |
| Amide II 60% N-H bend and 40% C-N stretch | 0.33 ± 0.001 b | 0.58 ± 0.001 a | <0.0001 |
| Amide III 40% C-N stretch, 30% N-H bend | 0.74 ± 0.004 | 0.78 ± 0.006 | 0.061 |
| C-O-C, C-O stretching of glycogen, carbohydrate | 0.29 ± 0.002 b | 0.38 ± 0.002 a | <0.0001 |
| Items | % Curve Fitting ± SD | p-Value | |
|---|---|---|---|
| CO | OR | ||
| α-helix (1655 cm−1) | 54.40 ± 5.59 | 52.23 ± 5.94 | 0.163 |
| β-sheet (1622, 1627, 1630 cm−1) | 26.77 ± 2.28 | 25.44 ± 2.71 | 0.066 |
| Antiparallel β-sheet (1695 to 1685 cm−1) | 5.25 ± 6.52 | 5.08 ± 7.06 | 0.918 |
| β-turn (1670, 1678, 1680 cm−1) | 18.82 ± 5.85 | 22.32 ± 7.41 | 0.369 |
| α-helix/β-sheet | 2.05 ± 0.28 | 2.07 ± 0.26 | 0.750 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molee, W.; Khosinklang, W.; Tongduang, P.; Thumanu, K.; Yongsawatdigul, J.; Molee, A. Biomolecules, Fatty Acids, Meat Quality, and Growth Performance of Slow-Growing Chickens in an Organic Raising System. Animals 2022, 12, 570. https://doi.org/10.3390/ani12050570
Molee W, Khosinklang W, Tongduang P, Thumanu K, Yongsawatdigul J, Molee A. Biomolecules, Fatty Acids, Meat Quality, and Growth Performance of Slow-Growing Chickens in an Organic Raising System. Animals. 2022; 12(5):570. https://doi.org/10.3390/ani12050570
Chicago/Turabian StyleMolee, Wittawat, Wichuta Khosinklang, Pramkamon Tongduang, Kanjana Thumanu, Jirawat Yongsawatdigul, and Amonrat Molee. 2022. "Biomolecules, Fatty Acids, Meat Quality, and Growth Performance of Slow-Growing Chickens in an Organic Raising System" Animals 12, no. 5: 570. https://doi.org/10.3390/ani12050570
APA StyleMolee, W., Khosinklang, W., Tongduang, P., Thumanu, K., Yongsawatdigul, J., & Molee, A. (2022). Biomolecules, Fatty Acids, Meat Quality, and Growth Performance of Slow-Growing Chickens in an Organic Raising System. Animals, 12(5), 570. https://doi.org/10.3390/ani12050570