
The Thyroid Hormone and Immunoglobulin Concentrations in Blood Serum and Thyroid Gland Morphology in Young Hens Fed with Different Diets, Sources, and Levels of Iodine Supply
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Management
2.2. Diets and Feeding Program
2.3. Analytical Methods in Feed Mixtures
2.4. Blood Collection and Analysis
2.5. Histological Examinations
2.6. Statistical Evaluation of Results
3. Results
3.1. Thyroid Hormone Concentration in Hen Blood Serum
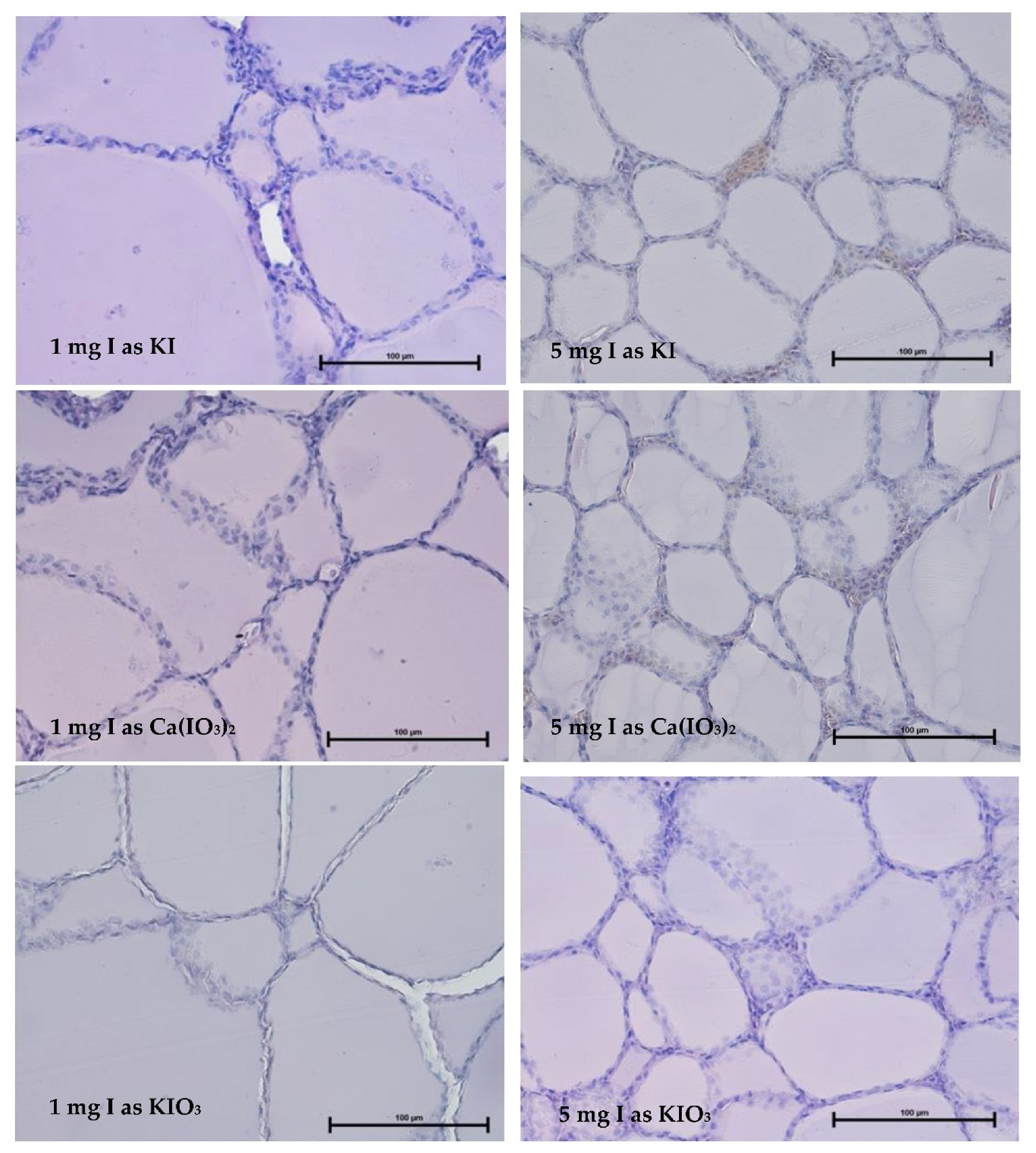
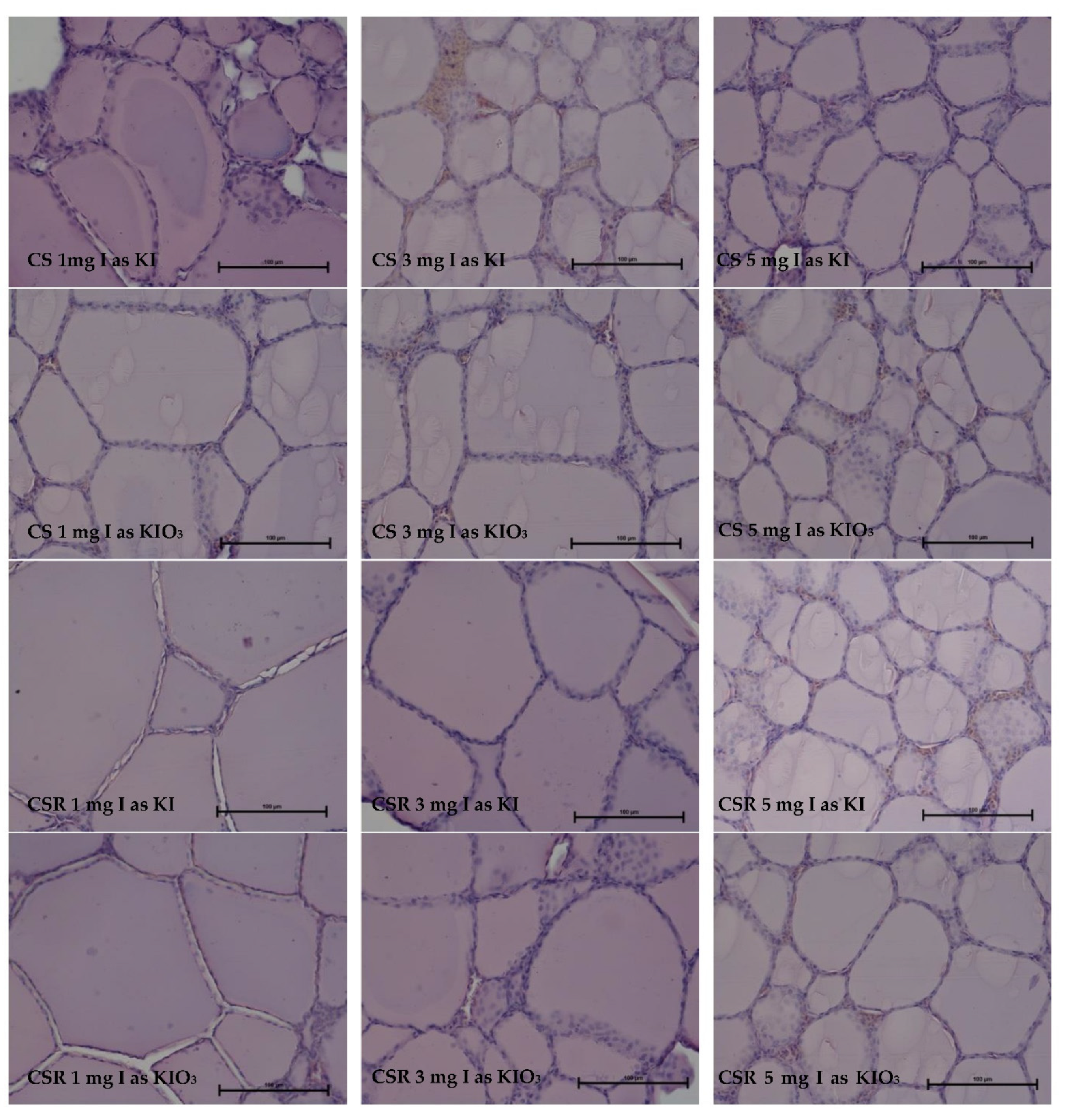
3.2. Thyroid Gland Structure
3.3. Immunoglobulin Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sturkie, P.D. (Ed.) Avian Physiology; Springer: New York, NY, USA, 1986. [Google Scholar]
- Klandorf, H.; Sharp, P.J.; Duncan, I.J.H. Variations in levels of plasma thyroxine and triiodothyronine in juvenile female chickens during 24- and 16-hr lighting cycles. Gen. Comp. Endocrinol. 1978, 36, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Shellabarger, C.J. A comparison of triiodothyronine and thyroxine in the chick goiter-prevention test. Poult. Sci. 1955, 34, 1437–1440. [Google Scholar] [CrossRef]
- Tata, J.R.; Shellabarger, C.J. An explanation for the difference between the responses of mammals and birds to thyroxine and tri-iodothyronine. Biochem. J. 1959, 72, 608–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobek, S.; Jastrzebski, M.; Pietras, M. Age-related changes in oxygen consumption and plasma thyroid hormone concentration in the young chicken. Gen. Comp. Endocrinol. 1976, 31, 169–174. [Google Scholar] [CrossRef]
- Sourgens, H.; Frőmblich-Borges, A.; Kemper, P.H. Iodine and Autoregulation of Thyroid Gland Function: Inhibition of Secretion of Thyroid Hormones in the Rat. 5. Spurenelement-Symposium, Jod. J.Boden, Pflanze, Tier, Mensch; Friedrich-Schiller-Universitat: Jena, Germany, 1986; pp. 53–59. [Google Scholar]
- McDowell, L.R. Iodine. In Minerals in Animal and Human Nutrition; Academic Press Inc.: New York, NY, USA, 1992; pp. 225–245. [Google Scholar]
- Delange, F. Risks and benefits of iodine supplementation. Lancet 1998, 351, 923–924. [Google Scholar] [CrossRef]
- Goldhaber, S.B. Trace element risk assessment: Essentiality vs. toxicity. Regul. Toxicol. Pharmacol. 2003, 38, 232–242. [Google Scholar] [CrossRef]
- Lewis, P.D. Responses of domestic flow to excess iodine: A review. Br. J. Nutr. 2004, 91, 29–39. [Google Scholar] [CrossRef]
- Li, Q.; Mair, C.; Schedle, K.; Hammerl, S.; Schodl, K.; Windisch, W. Effect of iodine source and dose on growth and iodine content in tissue and plasma thyroid hormones in fattening pigs. Eur. J. Nutr. 2012, 51, 685–691. [Google Scholar] [CrossRef]
- Delange, F. The disorders induced by iodine deficiency. Thyroid. 1994, 4, 107–128. [Google Scholar] [CrossRef] [Green Version]
- Van Middlesworth, L. Iodinated Thyroid Proteins Which Accumulate during Iodine Deficiency. 16. Arbeitstagung “Mengen und Spurenelemente”; Friedrich-Schiller-Universitat: Jena, Germany, 1996; pp. 920–921. [Google Scholar]
- Liu, N.Q.; Xu, Q.; Hou, X.L.; Liu, P.S.; Chai, Z.F.; Zhu, L.; Zhao, Z.Y.; Wang, Z.H.; Li, Y.F. The distribution patterns of trace element in the brain and erythrocytes in a rat experimental model of iodine deficiency. Brain Res. Bull. 2001, 55, 309–312. [Google Scholar] [CrossRef]
- Bagchi, N.; Brown, T.R.; Sundick, R.S. Thyroid cell injury is an initial event in the production of autoimmune thyroiditis by iodine in obese strain chicken. Endocrinology 1995, 136, 5054–5060. [Google Scholar] [CrossRef] [PubMed]
- Lydygard, P.; Whelan, A.; Fanger, M.W. Instant Notes in Immunology; Hames, B.D., Ed.; BIOS Scientific Publisher, Taylor and Francis Group: Abingdon, UK, 2000. [Google Scholar]
- Arrington, L.R.; Santa Cruz, R.A.; Harms, R.H.; Wilson, H.R. Effects of excess dietary iodine upon pullets and laying hens. J. Nutr. 1967, 92, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Bürgi, H.; Schaffner, T.H.; Seiler, J.P. The toxicology of iodate: A review of the literature. Thyroid 2001, 11, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.H. Iodine toxicity and its amelioration. Exp. Biol. Med. 2004, 229, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Ślebodziński, A. Zarys Endokrynologii Zwierząt Domowych; Outline of Endocrinology of Domestical Animals; PWN: Warszawa, Poland, 1979; pp. 174–194. (In Polish) [Google Scholar]
- Kopp, W. Nutrition, evolution and thyroid hormone levels-a link to iodine deficiency disorders? Med. Hypotheses 2004, 62, 871–875. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.T.; Dunn, A.D. The importance of thyroglobulin structure for thyroid hormone biosynthesis. Biochimie 1999, 81, 505–509. [Google Scholar] [CrossRef]
- Miller, E.R.; Ammerman, C.B. Iodine bioavailability. In Bioavailability of Nutrients for Animals; Ammerman, C.B., Baker, D.H., Lewis, A.J., Eds.; Academic Press: San Diego, CA, USA, 2005; pp. 157–167. [Google Scholar]
- Sarlak, S.; Tabeidian, S.A.; Toghyani, M.; Shahraki, A.D.F.; Goli, M.; Habibian, M. Supplementation of two sources and three levels of iodine in the diet of laying hens: Effects on performance, egg quality, serum and egg yolk lipids, antioxidant status, and iodine accumulation in eggs. Ital. J. Anim. Sci. 2020, 19, 974–988. [Google Scholar] [CrossRef]
- Röttger, A.S.; Halle, I.; Wagner, H.; Breves, G.; Dänicke, S.; Flachowsky, G. The effects of iodine level and source on iodine carry-over in eggs and body tissues of laying hens. Arch. Anim. 2012, 66, 385–401. [Google Scholar] [CrossRef]
- Preston, R.C. Serum inorganic iodine dynamics in cattle following a single oral dose of several iodine sources. Faseb J. 2009, 8, A43. [Google Scholar]
- Leeson, S.; Summers, J.D. Commercial Poultry Nutrition, 3rd ed.; Nottingham University Press: Nottingham, UK, 2009; pp. 72–73. [Google Scholar]
- Bakhshalinejad, R.; Hassanabadi, A.; Nassiri-Moghaddam, H.; Zarghi, H. The effects of dietary calcium iodate on productive performance, egg quality and iodine accumulation in eggs of laying hens. J. Anim. Physiol. Anim. Nutr. 2018, 102, 746–754. [Google Scholar] [CrossRef]
- Słupczyńska, M.; Jamroz, D.; Orda, J.; Wiliczkiewicz, A. Effect of various sources and levels of iodine, as well as the kind of diet, on the performance of young laying hens, iodine accumulation in eggs, egg characteristics, and morphotic and biochemical indices in blood. Poult. Sci. 2014, 93, 2536–2547. [Google Scholar] [CrossRef] [PubMed]
- Smulikowska, S.; Rutkowski, A. Zalecenia Żywieniowe i Wartość Pokarmowa pasz dla Drobiu; Poultry Nutrition Standards. Nutritional Recommendations and Nutritive Value of Feeds in Poultry, 5th ed.; Instytut Fizjologii i Żywienia Zwierząt im. Jana Kielanowskiego, Polskiej Akademii Nauk, Polski Oddział Światowego Stowarzyszenia Wiedzy Drobiarskiej PB WPSA: Poznań, Poland, 2018; p. 37. (In Polish) [Google Scholar]
- EFSA. Opinion of the scientific panel on additives and products or substances used in animal feed on the request from the Commission on the use of iodine in feedingstuffs. EFSA J. 2005, 13, 1–42. [Google Scholar]
- Tripathi, M.K.; Agrawal, I.S.; Sharma, S.D.; Mishra, D.P. Effect of untreated, HCl treated or copper and iodine supplemented high glucosinolate mustard (Brassica juncea) meal on nutrient utilization, liver enzymes, thyroid hormones and growth of calves. Anim. Feed Sci. Technol. 2001, 92, 73–85. [Google Scholar] [CrossRef]
- Lichovnikova, M.; Zeman, L. The effects of a higher amount of iodine supplement on the efficiency of laying hens fed extruded rapeseed and on eggshell quality. Czech J. Anim. Sci. 2004, 49, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Haldimann, M.; Alt, A.; Blanc, A.; Blondeau, K. Iodine content of food groups. J. Food Compos. Anal. 2005, 18, 461–471. [Google Scholar] [CrossRef]
- World’s Poultry Science Association. European Table of Energy Values for Poultry Feedstuffs, 3rd ed.; WPSA: Poznań, Poland, 1989; pp. 11–28. [Google Scholar]
- AOAC. Official Methods of Analysis., 18th ed.; Association of Official Analytical Chemists (AOAC): Arlington, VA, USA, 2007. [Google Scholar]
- Moore, S.; Stein, N.H. Discussion of classic methods of acid hydrolysis. Methods Enzymol. 1963, 6, 819–823. [Google Scholar]
- Gąsior, R.; Szczypuła, M. Validation of method for determination of iodine in food and biological material. Rocz. Nauk. Zoot. 2010, 37, 63–73. (In Polish) [Google Scholar]
- Knapp, G.; Maichin, B.; Fecher, P.; Hasse, S.; Schramel, P. Iodine determination in biological materials: Options for sample preparation and final determination. J. Anal. Chem. 1998, 362, 508–513. [Google Scholar]
- Zhang, W.; Mnastsakanov, A.; Hower, R.; Cantor, H.; Wang, Y. Urinary iodine assays and ionophore based potentiometric iodide sensors. Front. Biosci. 2005, 10, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Gabe, M. Techniques Histologiques; Masson, P., Ed.; Masson: Paris, France, 1968; p. 1113. [Google Scholar]
- StatSoft Inc. Statistica Data Analysis Software System; Version 9; StatSoft Inc.: Tulsa, OK, USA, 2012. [Google Scholar]
- Mehner, A.; Hartfiel, W. Handbuch der Geflügelphysiologie. Teil 2; Karger Verlag: Basel, Switzerland; München, Germany, 1983; pp. 344–345. [Google Scholar]
- Eshratkhah, B.; Asad Zadeh, S.; Forouzan, V.; Pour Parsa, A.A.; Ghiasi Ghalehkandi, J. Comparative study on the determination of serum thyroid hormones by two methods of immunoassay in broiler breeder poultry. Comp. Clin. Path. 2011, 20, 337–340. [Google Scholar] [CrossRef]
- Stojević, Z.; Milinković-Tur, S.; Ćurčija, K. Changes in thyroid hormones concentrations in chicken blood plasma during fattening. Veterinarski Arhiv. 2000, 70, 31–37. [Google Scholar]
- Moayyedian, H.; Asasi, K.; Nazifi, S.; Hassanzadeh, M.; Ansari-Lari, M. Relationship between venous blood gas parameters, thyroid hormone levels and ascites syndrome in broiler chickens exposed to cold temperature. Iran. J. Vet. Res. 2011, 12, 31–38. [Google Scholar]
- Ma, H.T.; Tang, X.; Tian, C.Y.; Zou, S.X.; Huang, G.Q.; Chen, W.H. Effects of dehydroepiandrosterone on growth performance, lipid metabolic hormones and parameters in broilers. Vet. Medic. 2008, 53, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Eshratkhah, B.; Ali Pourparsa, A.; Rostami, S.; Abbasi Navid, M.R. Relationship between the variations of blood thyroid hormones concentrations, their percentages and some biochemical parameters during different ages of layer breeder. Glob. Vet. 2011, 6, 459–467. [Google Scholar]
- He, M.L.; Borger, C.; Schuster, M.; Hollwich, W.; Krupova, V.; Kursa, J.; Bonabeze, E.; Ranzland, D.; Rambeck, W.W. Carryover of Iodine from Algae to the Products of Pigs and Layng Hens. 6th Symp.; Pig and Poultry Nutrition: Wittenberg, Germany, 2000; pp. 251–253. [Google Scholar]
- Travniček, J.; Kroupova, V.; Krotochvil, P.; Krabacova, J. The effect of excessive iodine intake on the histology of the thyroid gland in layer. Vet. Med. 1999, 6, 177–182. [Google Scholar]
- Behroozlak, M.; Daneshyar, M.; Farhomand, P. The effect of dietary iodine and its consumption duration on performance, carcass characteristics, meat iodine, thyroid hormones and some blood indices in broiler chickens. J. Anim. Physiol. Anim. Nutr. 2020, 104, 876–885. [Google Scholar] [CrossRef]
- Ahmadi, M. The Effect of Different Levels of Rapeseed Meal with and without Enzyme on the Performance and the Serum Level of Triiodothyronine (T3), Thyroxine (T4) and Thyroid Stimulating Hormone (TSH) in Broiler Chickens. Iran. J. Appl. Anim. Sci. 2016, 6, 203–209. [Google Scholar]
- Maroufyan, E.; Kermanshahi, H. Effect of Different Levels of Rapeseed Meal Supplemented with Calcium Iodate on Performance, Some Carcass Traits, and Thyroid Hormones of Broiler Chickens. Int. J. Poult. Sci. 2006, 5, 1073–1078. [Google Scholar]
- Reinecke, E.P.; Turner, C.W. Seasonal rhythm in the thyroid hormone secretion of the chick. Poult. Sci. 1945, 24, 499–503. [Google Scholar] [CrossRef]
- French, E.I.; Hodges, R.D. Fine structural studies on the thyroid gland of the normal domestic fowl. Cell Tissue. Res. 1977, 178, 397–410. [Google Scholar] [CrossRef]
- Breit, S.; König, H.E.; Stöger, E. The morphology of the thyroid gland in poultry with special regard to seasonal variations. Anat. Histol. Embryol. 1998, 27, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Takashi, Y. Thyroid hormone and seasonal regulation of reproduction. Front. Neuroendocr. 2013, 34, 157–166. [Google Scholar]
- Niemiec, T. The Effect of Invertebrate Extracts on Development and Health of Model Animals; SGGW: Warszawa, Poland, 2013. (In Polish) [Google Scholar]



{kind=link}
{kind=link}
| Experiment I | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Type of iodine supplement | |||||||||||
| potassium iodide KI | calcium iodate Ca(IO3)2 | potassium iodate KIO3 | |||||||||
| Level of I-supplement mg/kg | |||||||||||
| 1 | 5 | 1 | 5 | 1 | 5 | ||||||
| Experiment II | |||||||||||
| Treatments—Diets | |||||||||||
| CS * | CSR ** | ||||||||||
| Type of iodine supplement | |||||||||||
| potassium iodide KI | potassium iodate KIO3 | potassium iodide KI | potassium iodate KIO3 | ||||||||
| Level of I-supplement mg/kg | |||||||||||
| 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 |
| Ingredient, % of Diet | Diets | ||
|---|---|---|---|
| Experiment I | Experiment II | ||
| CS | CSR | ||
| Corn meal | 15.00 | 15.00 | 15.00 |
| Wheat meal | 25.81 | 28.44 | 24.09 |
| Barley meal | 20.00 | 20.00 | 20.00 |
| Soybean meal solvent extracted | 21.50 | 20.18 | 13.23 |
| Rapeseed 00 meal. solvent extracted | - | - | 10.00 |
| Rapeseed oil | 5.20 | 4.28 | 5.71 |
| Dicalcium phosphate | 2.05 | 0.481 | 0.435 |
| Chalk | 7.25 | 8.931 | 8.851 |
| NaCl | 0.37 | 0.015 | 0.013 |
| DL methionine | 0.184 | 0.046 | 0.024 |
| L-Lysine | 0.135 | 0.128 | 0.148 |
| Vitamin and mineral premix 1 | 2.50 | 2.50 | 2.50 |
| Nutritive value of mixtures | |||
| EM, MJ/kg | 11.65 | 11.65 | 11.65 |
| Crude protein, g/kg | 163.50 | 165.00 | 162.00 |
| Crude fiber, g/kg | 33.00 | 36.26 | 28.97 |
| Ca, g/kg | 38.80 | 38.80 | 38.90 |
| P available, g/kg | 4.30 | 4.30 | 4.32 |
| Na, g/kg | 1.75 | 1.70 | 1.71 |
| Methionine, g/kg | 4.32 | 4.30 | 4.31 |
| Lysine, g/kg | 9.08 | 9.00 | 8.97 |
| Nutritive recommendations for Hy-Line laying hens | |||
| EM, MJ/kg | 11.65 | ||
| Crude protein, g/kg | 165.00 | ||
| Crude fiber, g/kg | to 40.00 | ||
| Ca, g/kg | 38.80 | ||
| P available, g/kg | 4.30 | ||
| Na, g/kg | 1.70 | ||
| Methionine, g/kg | 4.30 | ||
| Lysine, g/kg | 9.00 | ||
| Item | Experiment I | Experiment II | ||
|---|---|---|---|---|
| fT3, pmol/L | fT4, pmol/L | fT3, pmol/L | fT4, pmol/L | |
| Treatments | ||||
| Experiment I | ||||
| 1 mg I as KI | 5.14 | 10.71 | ||
| 5 mg I as KI | 5.48 | 11.47 | ||
| 1 mg I as Ca(IO3)2 | 5.05 | 22.69 | ||
| 5 mg I as Ca(IO3)2 | 5.54 | 11.85 | ||
| 1 mg I as KIO3 | 4.64 | 9.92 | ||
| 5 mg I as KIO3 | 6.45 | 13.82 | ||
| Experiment II | ||||
| CS | ||||
| 1 mg I as KI | 8.51 | 11.06 | ||
| 3 mg I as KI | 7.01 | 11.58 | ||
| 5 mg I as KI | 10.14 | 9.52 | ||
| 1 mg I as KIO3 | 7.93 | 8.02 | ||
| 3 mg I as KIO3 | 9.04 | 11.36 | ||
| 5 mg I as KIO3 | 9.66 | 10.04 | ||
| CSR | ||||
| 1 mg I as KI | 11.19 | 8.55 | ||
| 3 mg I as KI | 9.29 | 7.22 | ||
| 5 mg I as KI | 13.29 | 7.28 | ||
| 1 mg I as KIO3 | 10.92 | 10.32 | ||
| 3 mg I as KIO3 | 10.70 | 7.59 | ||
| 5 mg I as KIO3 | 12.02 | 7.77 | ||
| SEM | 0.189 | 2.149 | 0.444 | 0.600 |
| p value | 0.111 | 0.492 | 0.137 | 0.920 |
| Kind of diet | ||||
| CS | 8.71 A | 10.26 | ||
| CSR | 11.24 B | 8.12 | ||
| Iodine source | ||||
| KI | 5.30 | 10.80 | 9.90 | 9.20 |
| Ca(IO3)2 | 5.29 | 20.13 | - | - |
| KIO3 | 5.54 | 11.30 | 10.05 | 9.19 |
| Level of iodine supplement | ||||
| 1 mg | 4.94 a | 15.62 | 9.64 Aa | 9.49 |
| 3 mg | - | - | 9.01 Ab | 9.43 |
| 5 mg | 5.83 b | 12.52 | 11.28 B | 8.65 |
| Source of variation, p value | ||||
| Kind of diet | - | - | 0.006 | 0.181 |
| Iodine source | 0.838 | 0.377 | 0.830 | 0.891 |
| Iodine level | 0.020 | 0.616 | 0.045 | 0.832 |
| Kind of diet × I-source | - | - | 0.874 | 0.576 |
| Kind of diet × I-level | - | - | 0.826 | 0.439 |
| I-source × I-level | 0.208 | 0.335 | 0.372 | 0.880 |
| Kind of diet × I-source × I-level | - | - | 0.744 | 0.769 |
| Item | Experiment I | Experiment II | ||
|---|---|---|---|---|
| Follicle Diameter, µm | Height of Follicle Epithelial Cells, µm | Follicle Diameter, µm | Height of Follicle Epithelial Cells, µm | |
| Experiment I | ||||
| 1 mg I as KI | 71.6 A | 3.87 | ||
| 5 mg I as KI | 54.2 Bc | 4.21 | ||
| 1 mg I as Ca(IO3)2 | 72.3 A | 4.58 | ||
| 5 mg I as Ca(IO3)2 | 53.9 Bc | 5.17 a | ||
| 1 mg I as KIO3 | 75.1 Ab | 3.22 b | ||
| 5 mg I as KIO3 | 59.0 Bc | 3.31 b | ||
| Experiment II | ||||
| CS | ||||
| 1 mg I as KI | 72.5 | 3.87 | ||
| 3 mg I as KI | 65.2 abce | 4.58 ABab | ||
| 5 mg I as KI | 54.2 Ac | 4.21 | ||
| 1 mg I as KIO3 | 87.1 Bde | 3.22 BCbcd | ||
| 3 mg I as KIO3 | 76 | 2.59 Cd | ||
| 5 mg I as KIO3 | 59.1 ACDabc | 3.31 bcd | ||
| CSR | ||||
| 1 mg I as KI | 88.7 Bd | 2.90 BC | ||
| 3 mg I as KI | 81.3 BCDacde | 3.78 cd | ||
| 5 mg I as KI | 67.5 | 5.17 Aa | ||
| 1 mg I as KIO3 | 84.1 BDde | 3.09 BCbcd | ||
| 3 mg I as KIO3 | 78.1 abde | 3.16 BCbcd | ||
| 5 mg I as KIO3 | 57.5 ACb | 4.43 abc | ||
| SEM | 1.640 | 0.166 | 1.534 | 0.111 |
| p value | 0.000 | 0.009 | 0.000 | 0.000 |
| Kind of diet | ||||
| CS | 69.0 a | 3.64 | ||
| CSR | 76.2 b | 3.76 | ||
| Iodine source | ||||
| KI | 62.9 | 4.04 | 71.6 | 4.04 A |
| Ca(IO3)2 | 63.1 | 4.88 A | ||
| KIO3 | 67.1 | 3.26 B | 73.7 | 3.29 B |
| Level of iodine supplement | ||||
| 1 mg | 72.9 A | 3.89 | 83.1 A | 3.43 A |
| 3 mg | 75.2 | 3.51 | ||
| 5 mg | 59.1 B | 4.23 | 67.8 B | 4.24 B |
| Source of variation, p value | ||||
| Kind of diet | 0.011 | 0.528 | ||
| Iodine source | 0.097 | 0.001 | 0.456 | 0.000 |
| Iodine level | 0.000 | 0.300 | 0.005 | 0.000 |
| Kind of diet × I-source | 0.004 | 0.049 | ||
| Kind of diet × I-level | 0.001 | 0.004 | ||
| I-source × I-level | 0.001 | 0.828 | 0.019 | 0.083 |
| Kind of diet × I-source × I-level | 0.000 | 0.487 | ||
| Item | IgA | IgM | IgG |
|---|---|---|---|
| Treatments | |||
| CS | |||
| 1 mg I as KI | 0.397 | 1.398 | 11.99 |
| 3 mg I as KI | 0.284 | 1.103 | 10.22 |
| 5 mg I as KI | 0.367 | 1.326 | 11.08 |
| 1 mg I as KIO3 | 0.280 | 1.236 | 11.81 |
| 3 mg I as KIO3 | 0.403 | 0.946 | 11.67 |
| 5 mg I as KIO3 | 0.284 | 0.985 | 11.57 |
| CSR | |||
| 1 mg I as KI | 0.285 | 1.255 | 10.32 |
| 3 mg I as KI | 0.349 | 1.080 | 13.73 |
| 5 mg I as KI | 0.275 | 1.432 | 9.71 |
| 1 mg I as KIO3 | 0.453 | 1.065 | 12.00 |
| 3 mg I as KIO3 | 0.301 | 1.055 | 9.99 |
| 5 mg I as KIO3 | 0.372 | 1.140 | 11.93 |
| SEM | 0.019 | 0.079 | 0.470 |
| p value | 0.643 | 0.927 | 0.914 |
| Kind of diet | |||
| CS | 0.336 | 1.166 | 11.39 |
| CSR | 0.339 | 1.171 | 11.28 |
| Iodine source | |||
| KI | 0.326 | 1.265 | 11.17 |
| KIO3 | 0.349 | 1.071 | 11.49 |
| Level of iodine supplement | |||
| 1 mg | 0.354 | 1.238 | 11.53 |
| 3 mg | 0.334 | 1.046 | 11.40 |
| 5 mg | 0.324 | 1.221 | 11.07 |
| Source of variation, | |||
| p value | |||
| Kind of diet | 0.942 | 0.757 | 0.811 |
| Iodine source | 0.483 | 0.354 | 0.602 |
| Iodine level | 0.857 | 0.589 | 0.906 |
| Kind of diet × I-source | 0.171 | 0.675 | 0.96 |
| Kind of diet × I-level | 0.935 | 0.410 | 0.672 |
| I-source × I-level | 0.939 | 0.639 | 0.548 |
| Kind of diet × I-source × I-level | 0.066 | 0.887 | 0.254 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Słupczyńska, M.; Jamroz, D.; Orda, J.; Wiliczkiewicz, A.; Kuropka, P.; Król, B. The Thyroid Hormone and Immunoglobulin Concentrations in Blood Serum and Thyroid Gland Morphology in Young Hens Fed with Different Diets, Sources, and Levels of Iodine Supply. Animals 2023, 13, 158. https://doi.org/10.3390/ani13010158
Słupczyńska M, Jamroz D, Orda J, Wiliczkiewicz A, Kuropka P, Król B. The Thyroid Hormone and Immunoglobulin Concentrations in Blood Serum and Thyroid Gland Morphology in Young Hens Fed with Different Diets, Sources, and Levels of Iodine Supply. Animals. 2023; 13(1):158. https://doi.org/10.3390/ani13010158
Chicago/Turabian StyleSłupczyńska, Maja, Dorota Jamroz, Janusz Orda, Andrzej Wiliczkiewicz, Piotr Kuropka, and Barbara Król. 2023. "The Thyroid Hormone and Immunoglobulin Concentrations in Blood Serum and Thyroid Gland Morphology in Young Hens Fed with Different Diets, Sources, and Levels of Iodine Supply" Animals 13, no. 1: 158. https://doi.org/10.3390/ani13010158
APA StyleSłupczyńska, M., Jamroz, D., Orda, J., Wiliczkiewicz, A., Kuropka, P., & Król, B. (2023). The Thyroid Hormone and Immunoglobulin Concentrations in Blood Serum and Thyroid Gland Morphology in Young Hens Fed with Different Diets, Sources, and Levels of Iodine Supply. Animals, 13(1), 158. https://doi.org/10.3390/ani13010158