
Differential Expression of Innate and Adaptive Immune Genes during Acute Physical Exercise in American Quarter Horses
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Sampling
2.2. RNA Extraction and qPCR
2.3. Data Analysis
3. Results
3.1. Normalization
3.2. Basal Differences
3.3. Differentially Expressed Genes
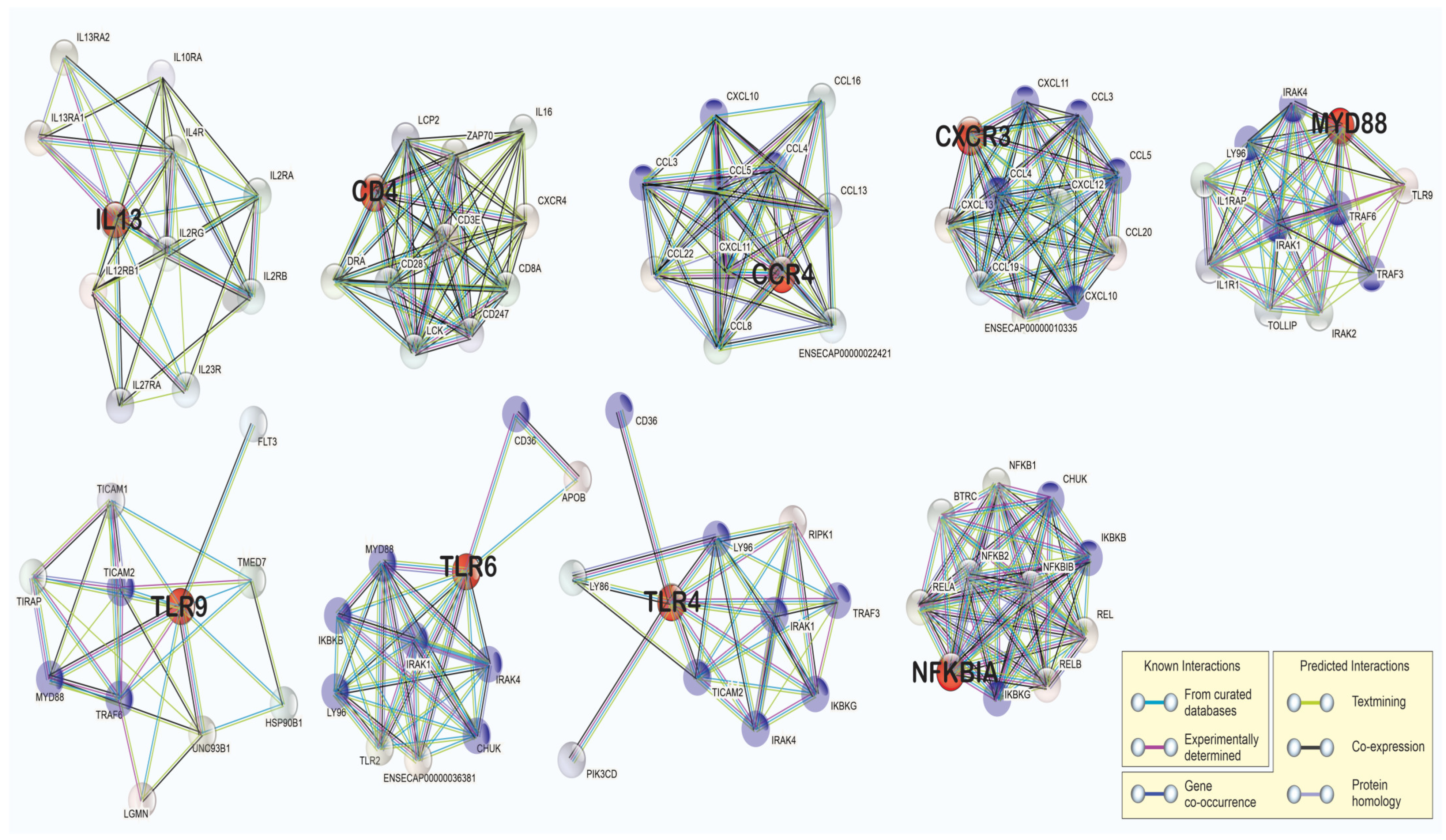
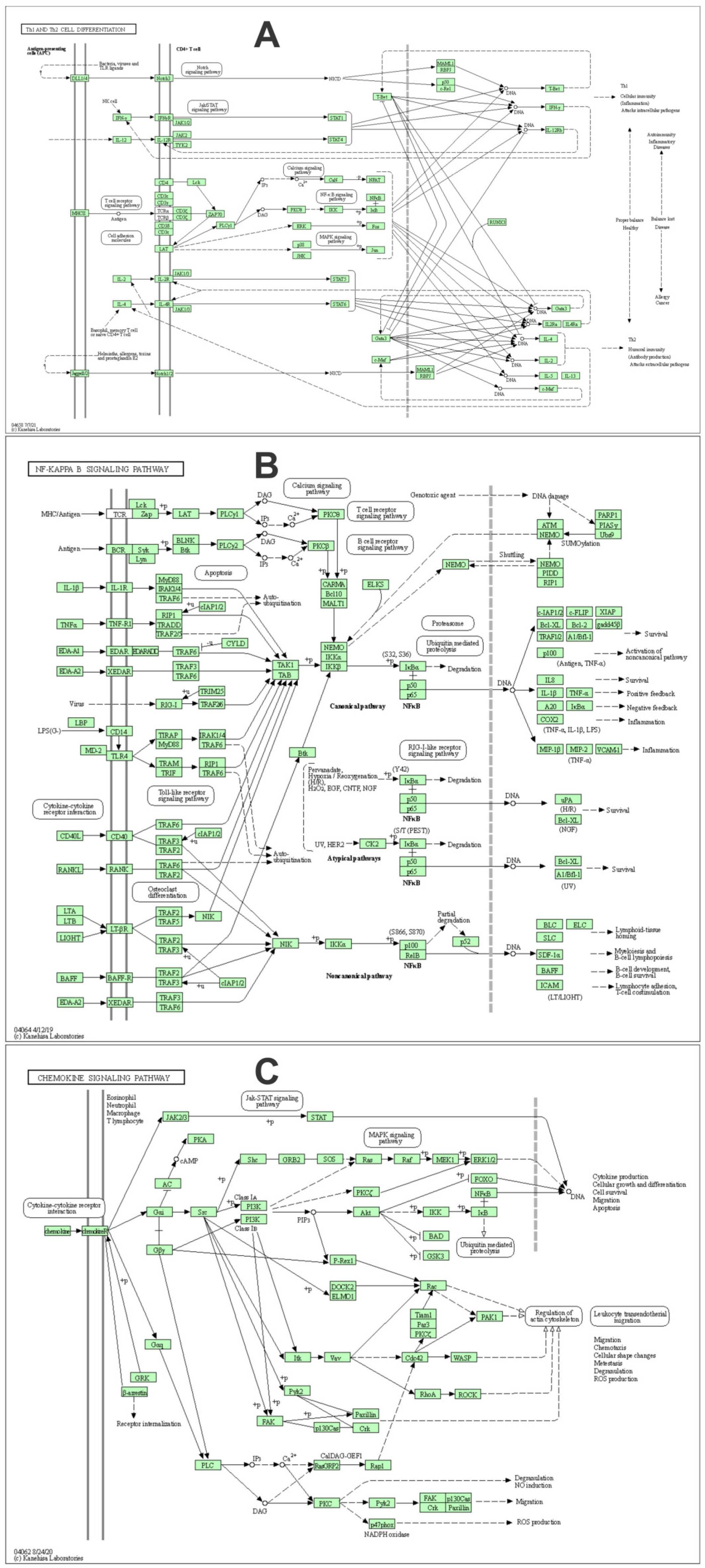
3.4. Pathway Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AQHA. Annual Report of the AQHA. 2021. Available online: https://www.aqha.com/documents/82601/1589238/2021+AQHA+Annual+Report.pdf/94454881-c00d-f587-4482-be16c5acf3a5?t=1649771187977 (accessed on 21 July 2022).
- Gleeson, M.; Bishop, N.C. Modification of immune responses to exercise by carbohydrate, glutamine and anti-oxidant supplements. Immunol. Cell Biol. 2000, 78, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Toft, A.D. Effects of exercise on lymphocytes and cytokines. Br. J. Sports Med. 2000, 34, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, N.P.; Oliver, S.J. Exercise, immune function and respiratory infection: An update on the influence of training and environmental stress. Immunol. Cell Biol. 2016, 94, 132–139. [Google Scholar] [CrossRef]
- Sakharov, D.A.; Maltseva, D.V.; Riabenko, E.A.; Shkurnikov, M.U.; Northoff, H.; Tonevitsky, A.G.; Grigoriev, A.I. Passing the anaerobic threshold is associated with substantial changes in the gene expression profile in white blood cells. Eur. J. Appl. Physiol. 2012, 112, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Capomaccio, S.; Vitulo, N.; Verini-Supplizi, A.; Barcaccia, G.; Albiero, A.; D’Angelo, M.; Campagna, D.; Valle, G.; Felicetti, M.; Silvestrelli, M.; et al. RNA sequencing of the exercise transcriptome in equine athletes. PLoS ONE 2013, 8, e83504. [Google Scholar] [CrossRef]
- Cappelli, K.; Amadori, M.; Mecocci, S.; Miglio, A.; Antognoni, M.T.; Elisabetta, R. Immune Response in Young Thoroughbred Racehorses under Training. Animals 2020, 10, 1809. [Google Scholar] [CrossRef] [PubMed]
- Büttner, P.; Mosig, S.; Lechtermann, A.; Funke, H.; Mooren, F.C. Exercise affects the gene expression profiles of human white blood cells. J. Appl. Physiol. 2007, 102, 26–36. [Google Scholar] [CrossRef]
- Capomaccio, S.; Cappelli, K.; Spinsanti, G.; Mencarelli, M.; Muscettola, M.; Felicetti, M.; Supplizi, A.V.; Bonifazi, M. Athletic humans and horses: Comparative analysis of interleukin-6 (IL-6) and IL-6 receptor (IL-6R) expression in peripheral blood mononuclear cells in trained and untrained subjects at rest. BMC Physiol. 2011, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Miglio, A.; Cappelli, K.; Capomaccio, S.; Mecocci, S.; Silvestrelli, M.; Antognoni, M.T. Metabolic and biomolecular changes induced by incremental long-term training in young thoroughbred racehorses during first workout season. Animals 2020, 10, 317. [Google Scholar] [CrossRef] [Green Version]
- Henneke, D.R.; Potter, G.D.; Kreider, J.L.; Yeates, B.F. Relationship between condition score, physical measurements and body fat percentage in mares. Equine Vet. J. 1983, 15, 371–372. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Cadegiani, F. Overtraining Syndrome in Athletes, 1st ed.; Springer: Cham, Switzerland, 2020. [Google Scholar]
- Liburt, N.R.; Adams, A.A.; Betancourt, A.; Horohov, D.W.; McKeever, K.H. Exercise-induced increases in inflammatory cytokines in muscle and blood of horses. Equine Vet. J. 2010, 42, 280–288. [Google Scholar] [CrossRef]
- Capomaccio, S.; Cappelli, K.; Barrey, E.; Felicetti, M.; Silvestrelli, M.; Verini-Supplizi, A. Microarray analysis after strenuous exercise in peripheral blood mononuclear cells of endurance horses (aged 5–13 years). Anim. Genet. 2010, 41, 166–175. [Google Scholar] [CrossRef]
- Beyeen, A.D.; Adzemovic, M.Z.; Öckinger, J.; Stridh, P.; Becanovic, K.; Laaksonen, H.; Lassmann, H.; Harris, R.A.; Hillert, J.; Alfredsson, L.; et al. IL-22RA2 associates with multiple sclerosis and macrophage effector mechanisms in experimental neuroinflammation. J. Immunol. 2010, 185, 6883–6890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappelli, K.; Felicetti, M.; Capomaccio, S.; Nocelli, C.; Silvestrelli, M.; Verini-Supplizi, A. Effect of training status on immune defence related gene expression in Thoroughbred: Are genes ready for the sprint? Vet. J. 2013, 195, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Ropka-Molik, K.; Stefaniuk-Szmukier, M.; Żukowski, K.; Piórkowska, K.; Gurgul, A.; Bugno-Poniewierska, M. Transcriptome profiling of Arabian horse blood during training regimens. BMC Genet. 2017, 18, 31. [Google Scholar] [CrossRef] [Green Version]
- Page, A.E.; Stewart, J.C.; Fielding, C.L.; Horohov, D.W. The effect of a 160-kilometer competitive endurance ride on inflammatory marker mRNA expression in horses. J. Equine Vet. Sci. 2019, 79, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, N.H.; Stanya, K.J.; Hyde, A.L.; Chalom, M.M.; Alexander, R.K.; Liou, Y.-H.; Starost, K.A.; Gangl, M.R.; Jacobi, D.; Liu, S.; et al. Interleukin-13 drives metabolic conditioning of muscle to endurance exercise. Science 2020, 368, eaat3987. [Google Scholar] [CrossRef]
- Zhu, M.; Min, S.; Mao, X.; Zhou, Y.; Zhang, Y.; Li, W.; Li, L.; Wu, L.; Cong, X.; Yu, G. Interleukin-13 promotes cellular senescence through inducing mitochondrial dysfunction in IgG4-related sialadenitis. Int. J. Oral Sci. 2022, 14, 29. [Google Scholar] [CrossRef]
- Anderson, C.A.; Patel, P.; Viney, J.M.; Phillips, R.M.; Solari, R.; Pease, J.E. A degradatory fate for CCR4 suggests a primary role in Th2 inflammation. J. Leukoc. Biol. 2020, 107, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Xiang, L.; Rehm, K.E.; Marshall, G.D., Jr. Effects of strenuous exercise on Th1/Th2 gene expression from human peripheral blood mononuclear cells of marathon participants. Mol. Immunol. 2014, 60, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Gleeson, M.; McFarlin, B.; Flynn, M. Exercise and Toll-like receptors. Exerc. Immunol. Rev. 2006, 12, 34–53. [Google Scholar]
- de Vicente, L.G.; Pinto, A.P.; da Rocha, A.L.; Pauli, J.R.; de Moura, L.P.; Cintra, D.E.; Ropelle, E.R.; da Silva, A.S. Role of TLR4 in physical exercise and cardiovascular diseases. Cytokine 2020, 136, 155273. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.D.; Zeng, K.; Liu, W.L.; Gao, Y.G.; Gong, C.S.; Zhang, C.X.; Chen, Y.Q. Effect of aerobic exercise on miRNA-TLR4 signaling in atherosclerosis. Int. J. Sports Med. 2014, 35, 344–350. [Google Scholar] [CrossRef]
- da Rocha, A.L.; Pinto, A.P.; Kohama, E.B.; Pauli, J.R.; de Moura, L.P.; Cintra, D.E.; Ropelle, E.R.; da Silva, A.S. The proinflammatory effects of chronic excessive exercise. Cytokine 2019, 119, 57–61. [Google Scholar] [CrossRef]
- Honda, K.; Yanai, H.; Negishi, H.; Asagiri, M.; Sato, M.; Mizutani, T.; Shimada, N.; Ohba, Y.; Takaoka, A.; Yoshida, N.; et al. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Nature 2005, 434, 772–777. [Google Scholar] [CrossRef]
- O’Brown, Z.K.; Van Nostrand, E.L.; Higgins, J.P.; Kim, S.K. The inflammatory transcription factors NFκB, STAT1 and STAT3 drive age-associated transcriptional changes in the human kidney. PLoS Genet. 2015, 11, e1005734. [Google Scholar] [CrossRef] [Green Version]
- Coto, E.; Díaz-Corte, C.; Tranche, S.; Gómez, J.; Alonso, B.; Iglesias, S.; Reguero, J.R.; López-Larrea, C.; Coto-Segura, P. Gene variants in the NF-KB pathway (NFKB1, NFKBIA, NFKBIZ) and their association with type 2 diabetes and impaired renal function. Hum. Immunol. 2018, 79, 494–498. [Google Scholar] [CrossRef]
- Blocquiaux, S.; Ramaekers, M.; Van Thienen, R.; Nielens, H.; Delecluse, C.; De Bock, K.; Thomis, M. Recurrent training rejuvenates and enhances transcriptome and methylome responses in young and older human muscle. JCSM Rapid Commun. 2022, 5, 10–32. [Google Scholar] [CrossRef]
- Gauthier, M.; Kale, S.L.; Oriss, T.B.; Scholl, K.; Das, S.; Yuan, H.; Hu, S.; Chen, J.; Camiolo, M.; Ray, P.; et al. Dual role for CXCR3 and CCR5 in asthmatic type 1 inflammation. J. Allergy Clin. Immunol. 2022, 149, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.; Gangur, V.; Simons, F.E.R.; HayGlass, K.T. Allergic humans are hypo-responsive to CXCR3 chemokines in a Th1 immunity-promoting loop. FASEB J. 2004, 18, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Tossige-Gomes, R.; Costa, K.B.; Ottone, V.D.O.; Magalhães, F.D.C.; Amorim, F.T.; Rocha-Vieira, E. Lymphocyte redox imbalance and reduced proliferation after a single session of high intensity interval exercise. PLoS ONE 2016, 11, e0153647. [Google Scholar] [CrossRef]
- Zhu, X.; Zhu, J. CD4 T helper cell subsets and related human immunological disorders. Int. J. Mol. Sci. 2020, 21, 8011. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Shao, Z.; Tan, Y.; Shen, Q.; Hou, L.; Yao, B.; Qin, J.; Xu, P.; Mao, C.; Chen, L.-N.; Zhang, H.; et al. Molecular insights into ligand recognition and activation of chemokine receptors CCR2 and CCR3. Cell Discov. 2022, 8, 44. [Google Scholar] [CrossRef]
- Smith, L.L. Overtraining, excessive exercise, and altered immunity. Sports Med. 2003, 33, 347–364. [Google Scholar] [CrossRef]
- Gholamnezhad, Z.; Boskabady, M.H.; Hossein, M.; Sankian, M.; Rad, A.K. Evaluation of immune response after moderate and overtraining exercise in wistar rat. IJBMS 2014, 17, 1. [Google Scholar]
- I Lancaster, G.; Halson, S.; Khan, Q.; Drysdale, P.; Wallace, F.; E Jeukendrup, A.; Drayson, M.; Gleeson, M. Effects of acute exhaustive exercise and chronic exercise training on type 1 and type 2 T lymphocytes. Exerc. Immunol. Rev. 2004, 10, 91–106. [Google Scholar]
- Wang, R.; Chen, P.J. Modulation of NKT cells and Th1/Th2 imbalance after α-GalCer treatment in progressive load-trained rats. Int. J. Biol. Sci. 2009, 5, 338–343. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Chen, P.; Wang, R.; Yu, D.; Zhang, Y.; Xiao, W. NADPH oxidase: A target for the modulation of the excessive oxidase damage induced by overtraining in rat neutrophils. Int. J. Biol. Sci. 2011, 7, 881. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
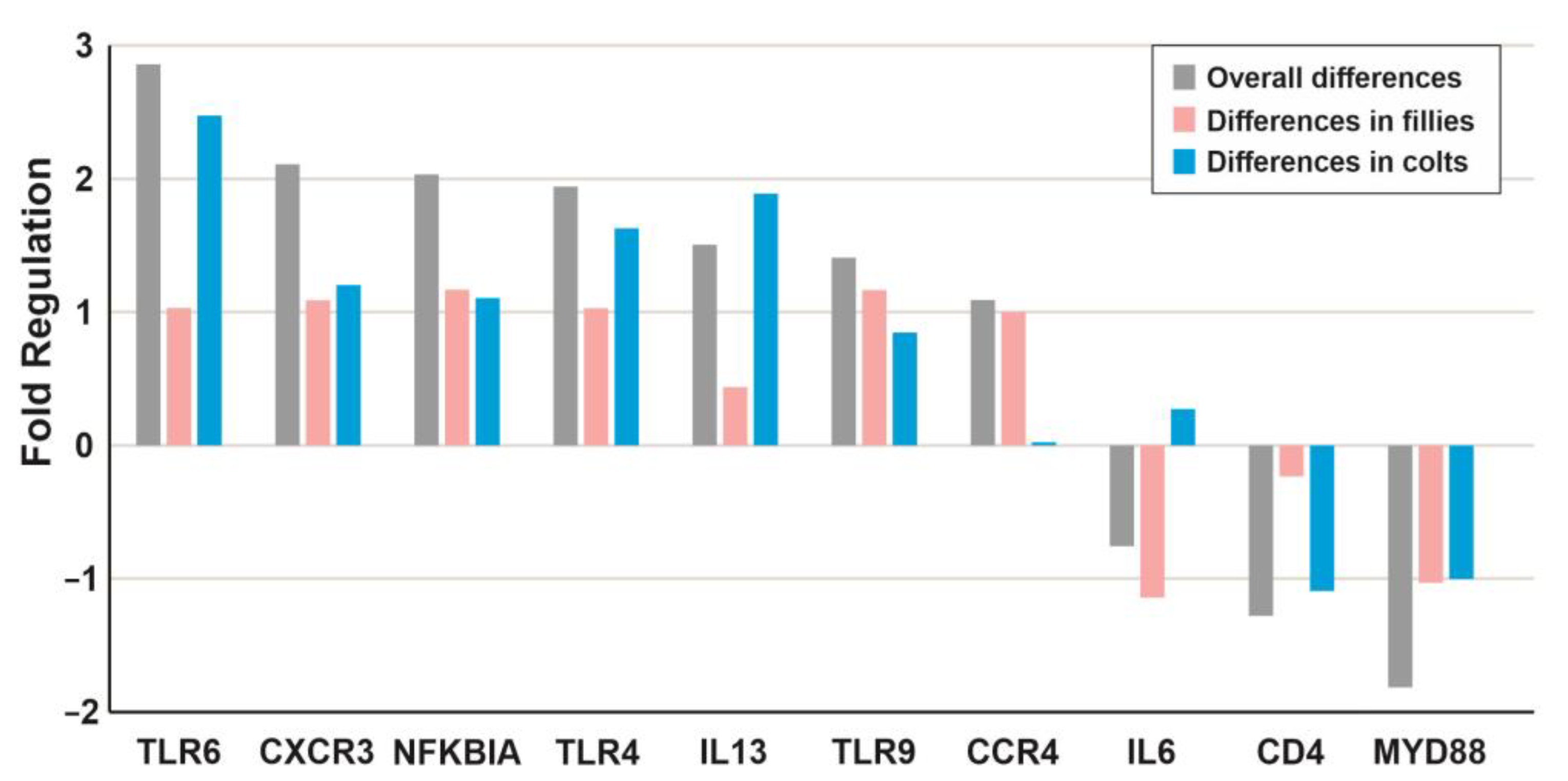
| Symbol | Name | RefSeq | Overall Fold Change | Overall p-Value | Female Fold Change | Female p-Value | Male Fold Change | Male p-Value |
|---|---|---|---|---|---|---|---|---|
| TLR6 | Toll-like receptor 6 | NM_001257142 | 2.88 | 0.003 | 1.04 | 0.044 | 2.49 | 0.045 |
| CXCR3 | C-X-C chemokine receptor type 3-like | XM_001493611 | 2.12 | 0.001 | 1.10 | 0.044 | 1.21 | 0.029 |
| NFKBIA | Nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha | XM_005603476 | 2.04 | 0.000 | 1.17 | 0.008 | 1.11 | 0.019 |
| TLR4 | Toll-like receptor 4 | NM_001099769 | 1.95 | 0.001 | 1.03 | 0.036 | 1.64 | 0.030 |
| IL13 | Interleukin 13 | NM_001143791 | 1.51 | 0.032 | 0.44 | 0.407 | 1.90 | 0.048 |
| TLR9 | Toll-like receptor 9 | NM_001081790 | 1.41 | 0.038 | 1.17 | 0.029 | 0.85 | 0.440 |
| CCR4 | Chemokine (C-C motif) receptor 4 | XM_005600863 | 1.09 | 0.034 | 1.00 | 0.021 | 0.02 | 0.612 |
| IL6 | Interleukin 6 (interferon, beta 2) | NM_001082496 | −0.76 | 0.159 | −1.14 | 0.026 | 0.26 | 0.547 |
| CD4 | T-cell surface glycoprotein CD4-like | XM_001497051 | −1.28 | 0.034 | −0.23 | 0.517 | −1.10 | 0.003 |
| MYD88 | Myeloid differentiation primary response protein MyD88-like | XM_001488549 | −1.81 | 0.000 | −1.03 | 0.007 | −1.01 | 0.006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson, J.; De Donato, M.; Appelbaum, B.; Garcia, C.T.; Peters, S. Differential Expression of Innate and Adaptive Immune Genes during Acute Physical Exercise in American Quarter Horses. Animals 2023, 13, 308. https://doi.org/10.3390/ani13020308
Wilson J, De Donato M, Appelbaum B, Garcia CT, Peters S. Differential Expression of Innate and Adaptive Immune Genes during Acute Physical Exercise in American Quarter Horses. Animals. 2023; 13(2):308. https://doi.org/10.3390/ani13020308
Chicago/Turabian StyleWilson, Judith, Marcos De Donato, Brooke Appelbaum, Carly Turner Garcia, and Sunday Peters. 2023. "Differential Expression of Innate and Adaptive Immune Genes during Acute Physical Exercise in American Quarter Horses" Animals 13, no. 2: 308. https://doi.org/10.3390/ani13020308
APA StyleWilson, J., De Donato, M., Appelbaum, B., Garcia, C. T., & Peters, S. (2023). Differential Expression of Innate and Adaptive Immune Genes during Acute Physical Exercise in American Quarter Horses. Animals, 13(2), 308. https://doi.org/10.3390/ani13020308