
Features of Fractal Conformity and Bioconsolidation in the Early Myogenesis Gene Expression and Their Relationship to the Genetic Diversity of Chicken Breeds
 ,
,  , , , , ,
, , , , ,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Birds and Raw Data Generation
2.2. Statistical and Mathematical Analyses
3. Results
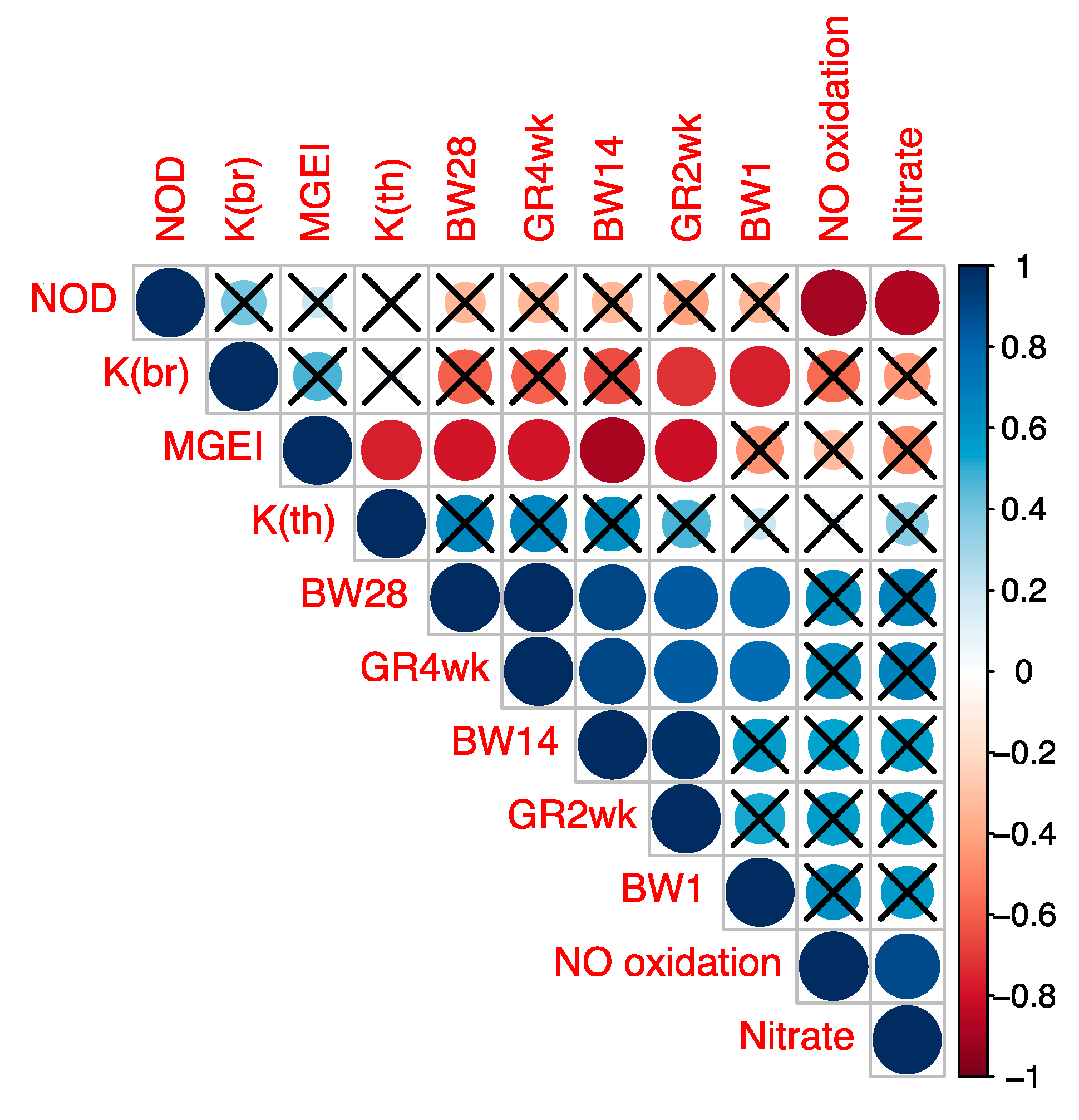
3.1. Relative Differential Gene Expression Data
3.2. Fractal Analysis of Myogenesis Gene Expression Structure in Various Chicken Breeds
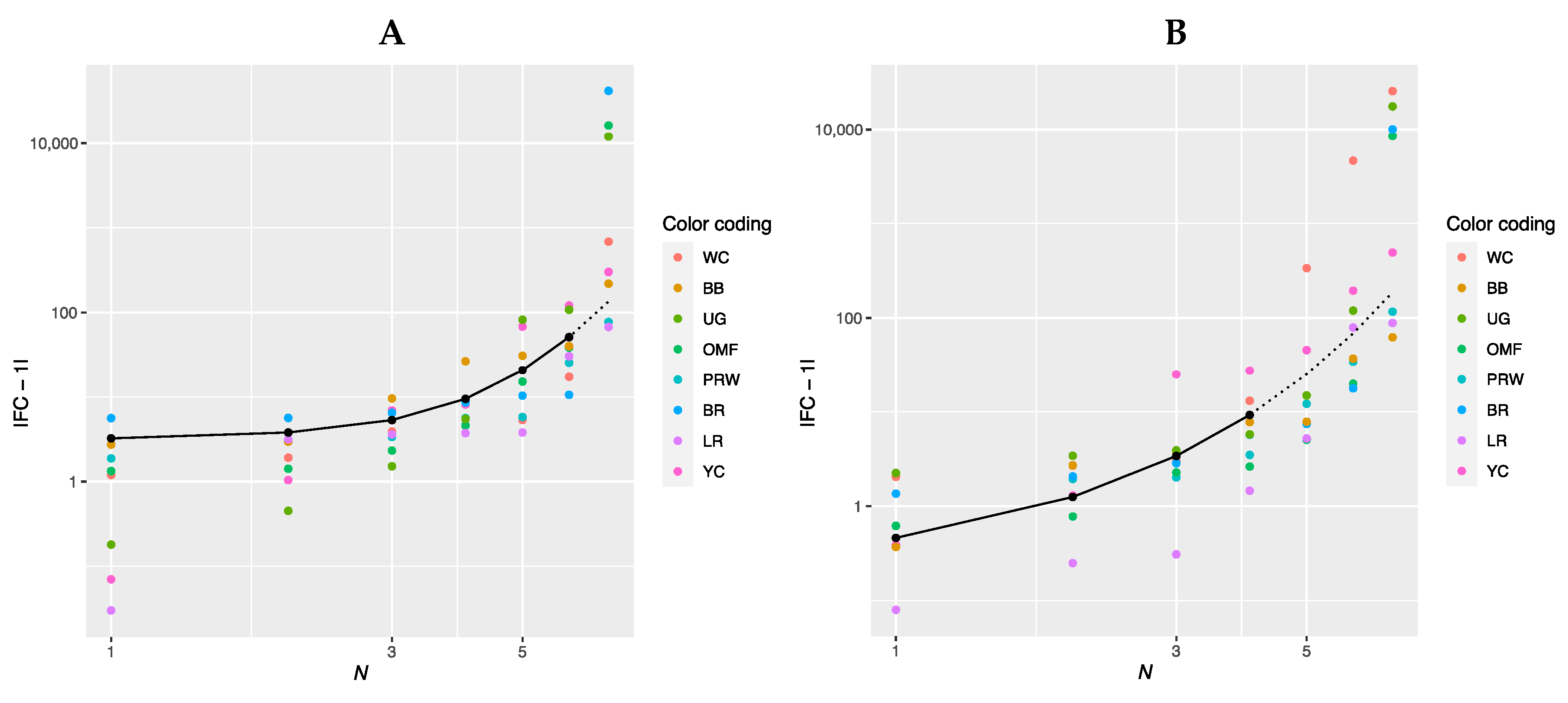
3.2.1. FC Value Transformation and Gene Ranking
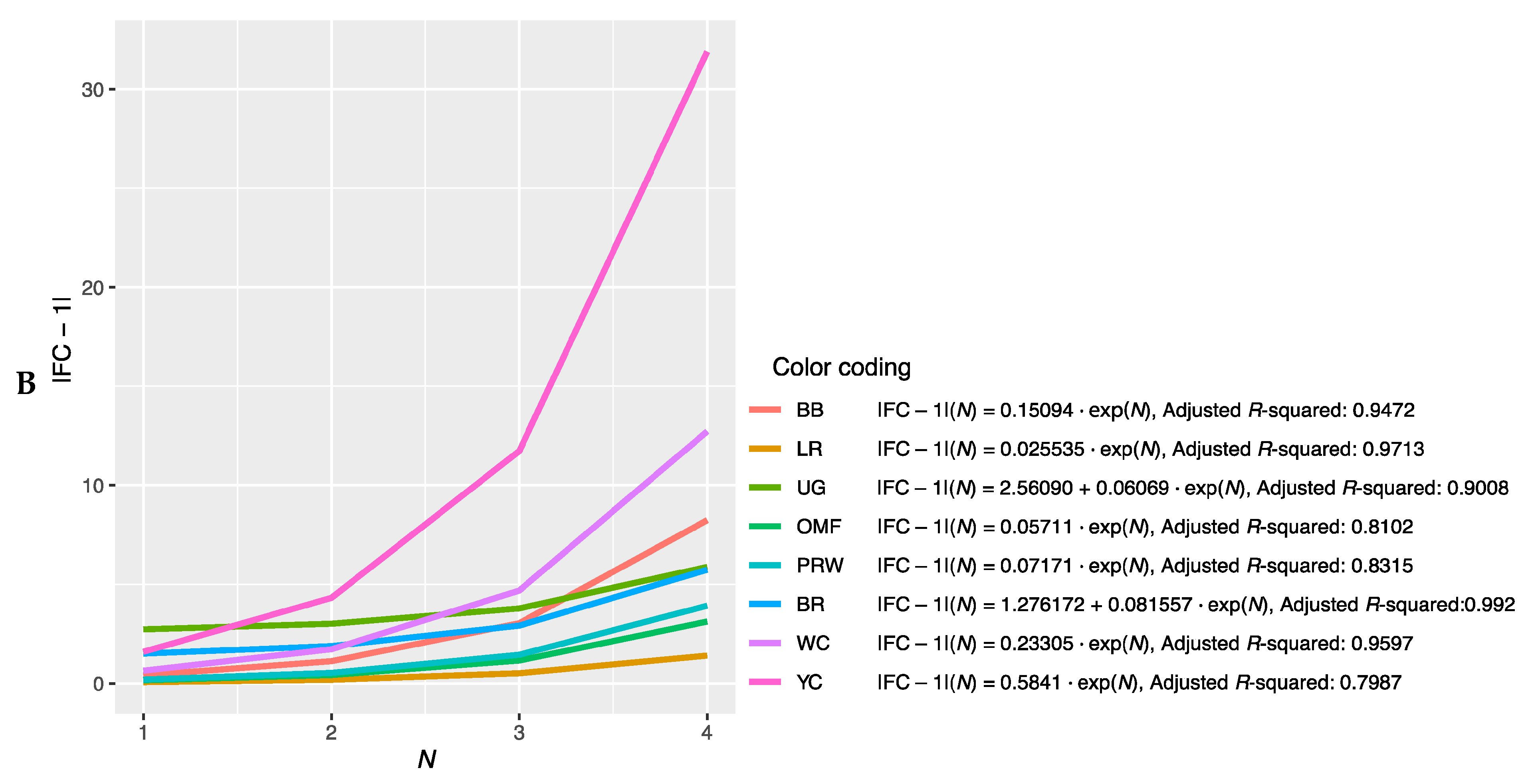
3.2.2. Approximation of the Dependence of FC Level on the Rank of Genes
3.2.3. Determining the Slope of the Function |FC − 1| = f(N)
3.2.4. Deducing the Myogenesis Gene Expression Index
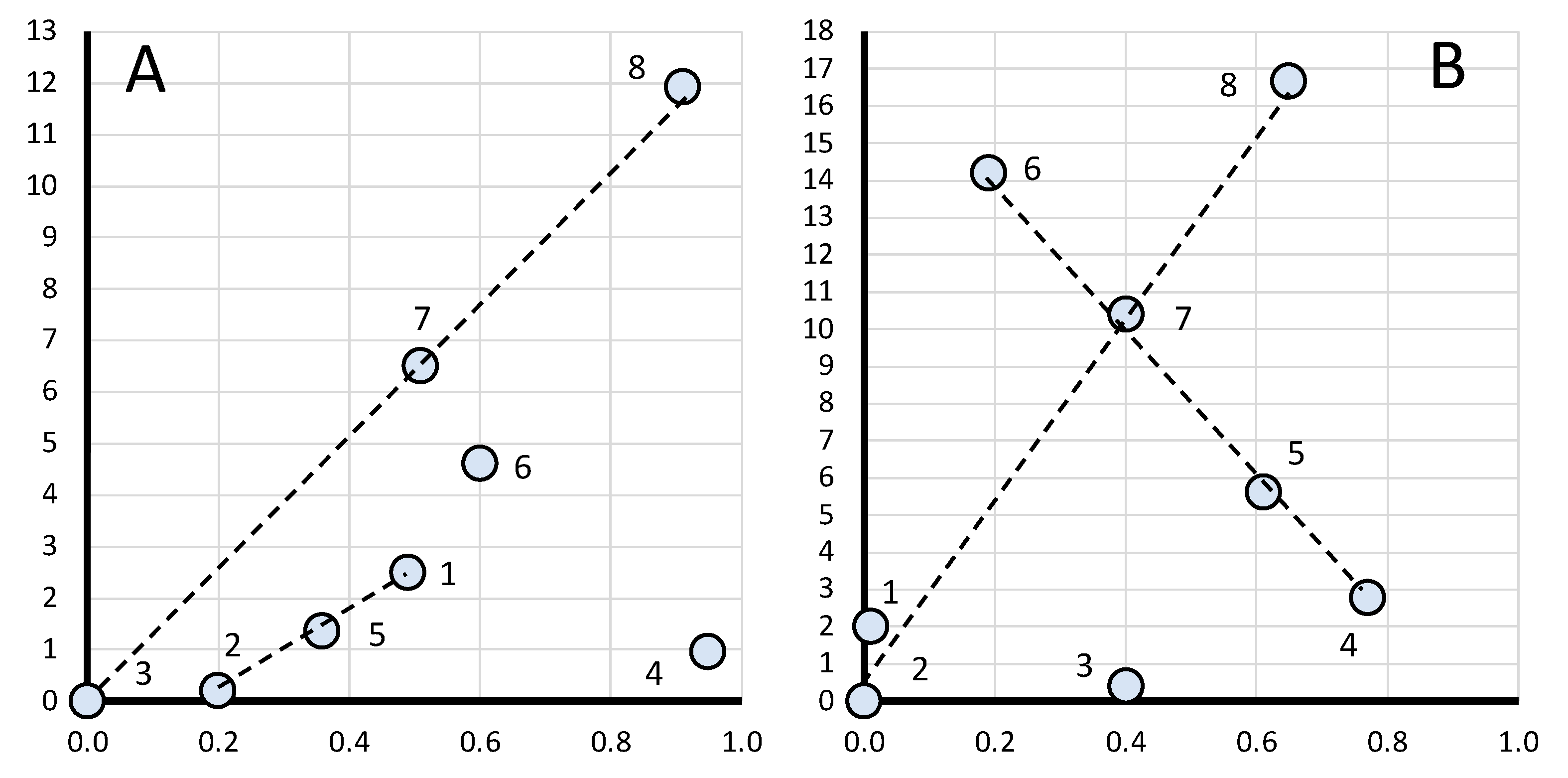
3.2.5. Defining Fractal Dimension D
3.3. Fractal Portraits and Fractal Bioconsolidation Index of Gene Expression
4. Discussion
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morse, D.R.; Lawton, J.H.; Dodson, M.M.; Williamson, M.H. Fractal dimension of vegetation and the distribution of arthropod body lengths. Nature 1985, 314, 731–733. [Google Scholar] [CrossRef]
- Şişli, A.B.; Pahnvar, A.J.; Engin, M.; Engin, E.Z. Obtaining the heart rate information from the speckle images by fractal analysis method. Celal Bayar Üniv. Fen Bilim. Derg. 2020, 16, 47–53. [Google Scholar] [CrossRef]
- Lee, C.Y. The fractal dimension as a measure for characterizing genetic variation of the human genome. Comput. Biol. Chem. 2020, 87, 107278. [Google Scholar] [CrossRef] [PubMed]
- Yakimov, B.N.; Solntsev, L.A.; Rozenberg, G.S.; Iudin, D.I.; Gelashvili, D.B. Scale invariance of biosystems: From embryo to community. Russ. J. Dev. Biol. 2014, 45, 168–176. [Google Scholar] [CrossRef]
- Yakimov, B.N.; Solntsev, L.A.; Rozenberg, G.S.; Iudin, D.I.; Shirokov, A.I.; Lokteva, O.A.; Gelashvili, D.B. Local multifractal analysis of the spatial structure of meadow comminities at small scale. Dokl. Biol. Sci. 2014, 458, 297–301. [Google Scholar] [CrossRef]
- Falconer, K. Fractal Geometry; Wiley: New York, NY, USA, 2003. [Google Scholar]
- Rihan, F.A. Numerical modeling of fractional-order biological systems. Abstr. Appl. Anal. 2013, 2013, 816803. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Lewin, H.A.; Robinson, G.E.; Kress, W.J.; Baker, W.J.; Coddington, J.; Crandall, K.A.; Durbin, R.; Edwards, S.V.; Forest, F.; Gilbert, M.T.P.; et al. Earth BioGenome Project: Sequencing life for the future of life. Proc. Natl. Acad. Sci. USA 2018, 115, 4325–4333. [Google Scholar] [CrossRef]
- Zhang, M.; Peng, W.F.; Hu, X.J.; Zhao, Y.X.; Lv, F.H.; Yang, J. Global genomic diversity and conservation priorities for domestic animals are associated with the economies of their regions of origin. Sci. Rep. 2018, 8, 11677. [Google Scholar] [CrossRef]
- Yang, G.; Ryo, M.; Roy, J.; Hempel, S.; Rillig, M.C. Plant and soil biodiversity have non-substitutable stabilising effects on biomass production. Ecol. Lett. 2021, 24, 1582–1593. [Google Scholar] [CrossRef]
- Yakimov, V.N. Methodology for the Analysis of Scaling Taxonomic, Phylogenetic and Functional Diversity of Biotic Communities. D.Sc. Thesis, Nizhny Novgorod State University named after N.I. Lobachevsky, Nizhny Novgorod, Russia, 2015. [Google Scholar]
- Larkina, T.A.; Barkova, O.Y.; Peglivanyan, G.K.; Mitrofanova, O.V.; Dementieva, N.V.; Stanishevskaya, O.I.; Vakhrameev, A.B.; Makarova, A.V.; Shcherbakov, Y.S.; Pozovnikova, M.V.; et al. Evolutionary subdivision of domestic chickens: Implications for local breeds as assessed by phenotype and genotype in comparison to commercial and fancy breeds. Agriculture 2021, 11, 914. [Google Scholar] [CrossRef]
- Ryabokon, Y.O.; Mykytyuk, D.M.; Frolov, V.V.; Katerynych, O.O.; Bondarenko, Y.V.; Mosyakina, T.V.; Gadyuchko, O.T.; Kovalenko, G.T.; Bogatyr, V.P.; Lyuty, Y.S. Catalog of Poultry Breeding Resources of Ukraine; Ryabokon, Y.O., Ed.; Poultry Research Institute: Kharkiv, Ukraine, 2005. [Google Scholar]
- Khvostyk, V.P.; Bondarenko, Y.V. Hereditary load in chicken populations of the domestic gene pool. Visnyk Sumsʹkoho Natsionalʹnoho Ahrarnoho Universytetu [Bull. Sumy Natl. Agrar. Univ.] 2016, 7, 112–114. [Google Scholar]
- Khvostyk, V.P.; Bondarenko, Y.V. Informational and statistical parameters of body weight of chickens of the domestic gene pool. Visnyk Sumsʹkoho Natsionalʹnoho Ahrarnoho Universytetu Seriya Tvarynnytstvo [Bull. Sumy Natl. Agrar. Univ. Ser. Livest.] 2017, 5, 161–165. [Google Scholar]
- Tagirov, M.T.; Tereshchenko, L.V.; Tereshchenko, A.V. Substantiation of the possibility of using primary germ cells as material for the preservation of poultry genetic resources. Ptakhivnytstvo 2006, 58, 464–473. [Google Scholar]
- Feder, J. Fractals; Plenum: New York, NY, USA, 1988. [Google Scholar] [CrossRef]
- Morozov, A.D. Introduction to the Theory of Fractals, 2nd ed.; Institute of Computer Science: Moscow, Russia; Izhevsk, Russia, 2002. [Google Scholar]
- Schroeder, M.R. Fractals, Chaos, Power Laws. Minutes from an Infinite Universe; W.H. Freeman: New York, NY, USA, 1991. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. An equilibrium theory of insular zoogeography. Evolution 1963, 17, 373–387. [Google Scholar] [CrossRef]
- Vorob’ev, N.I.; Nikonov, I.N.; Selina, M.V. Mathematical model of determining the index of fractal structures for estimating efficiency of probiotic fodder additives for microbiotes of the gut intestin. Vet. Zootekhniya I Biotekhnologiya [Vet. Med. Zootech. Biotechnol.] 2020, 4, 22–30. [Google Scholar] [CrossRef]
- Kochish, I.I.; Vorobyov, N.I.; Nikonov, I.N.; Selina, M.V. Fractal Bioconsolidation of Microorganisms in the Intestines of Laying Hens Due to the Use of a Feed Additive from the Mineral Shungite. In Proceedings of the Materials of the 2nd International Scientific and Practical Conference on Molecular Genetic Technologies for Analysis of Gene Expression Related to Animal Productivity and Disease Resistance, Moscow, Russia, 25 December 2020; pp. 59–75. [Google Scholar]
- Titov, V.Y.; Dolgorukova, A.M.; Fisinin, V.I.; Borkhunova, E.N.; Kondratov, G.V.; Slesarenko, N.A.; Kochish, I.I. The role of nitric oxide (NO) in the body growth rate of birds. Worlds Poult. Sci. J. 2018, 74, 675–686. [Google Scholar] [CrossRef]
- Titov, V.Y.; Kochish, I.I.; Nikonov, I.N.; Korenyuga, M.V.; Myasnikova, O.V.; Kuvanov, T.K.; Dolgorukova, A.M. Genetic Markers of Meat Performance in Poultry. In Proceedings of the Materials of the 2nd International Scientific and Practical Conference on Molecular Genetic Technologies for Analysis of Gene Expression Related to Animal Productivity and Disease Resistance, Moscow, Russia, 25 December 2020; pp. 136–150. [Google Scholar]
- Titov, V.; Dolgorukova, A.; Khasanova, L.; Kochish, I.; Korenyuga, M. Nitric oxide (NO) and arginine as factors for increasing poultry meat productivity. KnE Life Sci. 2021, 6, 622–631. [Google Scholar] [CrossRef]
- Titov, V.Y.; Dolgorukova, A.M.; Kochish, I.I. What Gene Expression is Associated with Nitric Oxide Oxidation in the Avian Embryo? In Proceedings of the Materials of the 3rd International Scientific and Practical Conference on Molecular Genetic Technologies for Analysis of Gene Expression Related to Animal Productivity and Disease Resistance, Moscow, Russia, 29 September 2021; pp. 411–421. [Google Scholar]
- Cazzato, D.; Assi, E.; Moscheni, C.; Brunelli, S.; De Palma, C.; Cervia, D.; Perrotta, C.; Clementi, E. Nitric oxide drives embryonic myogenesis in chicken through the upregulation of myogenic differentiation factors. Exp. Cell Res. 2014, 320, 269–280. [Google Scholar] [CrossRef]
- Dolgorukova, A.M.; Titov, V.Y.; Kochish, I.I.; Fisinin, V.I.; Nikonov, I.N.; Kosenko, O.V.; Myasnikova, O.V. The embryonic metabolism of nitric oxide and its interrelation with postembryonic development in chicken (Gallus gallus domesticus L.) and quails (Coturnix coturnix L.). Sel’skokhozyaistvennaya Biol. Agric. Biol. 2020, 55, 794–803. [Google Scholar] [CrossRef]
- Ulibarri, J.A.; Mozdziak, P.E.; Schultz, E.; Cook, C.; Best, T.M. Nitric oxide donors, sodium nitroprusside and S-nitroso-N-acetylpencillamine, stimulate myoblast proliferation in vitro. Vitr. Cell Dev. Biol. Anim. 1999, 35, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.E. A role for nitric oxide in muscle repair: Nitric oxide-mediated activation of muscle satellite cells. Mol. Biol. Cell. 2000, 11, 1859–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamler, J.S.; Meissner, G. Physiology of nitric oxide in skeletal muscle. Physiol. Rev. 2001, 81, 209–237. [Google Scholar] [CrossRef]
- Long, J.H.; Lira, V.A.; Soltow, Q.A.; Betters, J.L.; Sellman, J.E.; Criswell, D.S. Arginine supplementation induces myoblast fusion via augmentation of nitric oxide production. J. Muscle Res. Cell Motil. 2006, 27, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Willems, E.; Willemsen, H.; Franssens, L.; Buyse, J.; Decuypere, E.; Everaert, N. In ovo L-arginine supplementation stimulates myoblast differentiation but negatively affects muscle development of broiler chicken after hatching. J. Anim. Physiol. Anim. Nutr. 2016, 100, 167–177. [Google Scholar] [CrossRef]
- Tirone, M.; Conti, V.; Manenti, F.; Nicolosi, P.A.; D’Orlando, C.; Azzoni, E.; Brunelli, S. Nitric oxide donor molsidomine positively modulates myogenic differentiation of embryonic endothelial progenitors. PLoS ONE 2016, 11, e0164893. [Google Scholar] [CrossRef]
- Reddish, J.M.; Wick, M.; St-Pierre, N.R.; Lilburn, M.S. Analysis of myosin isoform transitions during growth and development in diverse chicken genotypes. Poult. Sci. 2005, 84, 1729–1734. [Google Scholar] [CrossRef]
- Tereshchenko, O.V.; Katerinich, O.O.; Pankova, S.M.; Borodai, V.P. Formation of genetic resources of domestic breeds of poultry in the context of food security of the state. Sučasne Ptahìvnictvo 2015, 7–8, 19–21. [Google Scholar]
- Bondarenko, Y.V.; Podstreshny, A.P. Genetic Monitoring of Chicken Populations. In Abstracts of the 2nd International Conference on Molecular Genetic Markers of Animals; Agrarna Nauka: Kiev, Ukraine, 1996; pp. 47–48. [Google Scholar]
- Romanov, M.N.; Weigend, S.; Bondarenko, Y.V.; Podstreshny, A.P.; Kutnyuk, P.I.; Sakhatsky, N.I. Studies on Poultry Germplasm Diversity and Conservation in Ukraine. In Proceedings of the Poultry Genetics Symposium, Mariensee, Germany, 6–8 October 1999; p. 140. [Google Scholar]
- Tixier-Boichard, M.; Coquerelle, G.; Vilela-Lamego, C.; Weigend, S.; Barre-Dirrie, A.; Groenen, M.; Crooijmans, R.; Vignal, A.; Hillel, J.; Freidlin, P.; et al. Contribution of Data on History, Management and Phenotype to the Description of the Diversity between Chicken Populations Sampled within the AVIANDIV Project. In Proceedings of the Poultry Genetics Symposium, Mariensee, Germany, 6–8 October 1999; pp. 15–21. [Google Scholar]
- Tkachik, T.E.; Kutnyuk, P.I.; Bondarenko, Y.V. Genetic load in land poultry populations. Ptakhivnytstvo 2005, 57, 94–98. [Google Scholar]
- Weigend, S.; Romanov, M.N.; Rath, D. Methodologies to Identify, Evaluate and Conserve Poultry Genetic Resources. In Proceedings of the XXII World’s Poultry Congress & Exhibition: Participant List & Full Text CD + Book of Abstracts, Istanbul, Turkey, 8–13 June 2004; p. 84. [Google Scholar]
- Narushin, V.G.; Laptev, G.Y.; Yildirim, E.A.; Ilina, L.A.; Filippova, V.A.; Kochish, I.I.; Gorfunkel, E.P.; Dubrovin, A.V.; Novikova, N.I.; Novikova, O.B.; et al. Modelling effects of phytobiotic administration on coherent responses to Salmonella infection in laying hens. Ital. J. Anim. Sci. 2020, 19, 282–287. [Google Scholar] [CrossRef]
- Vorobiev, N.I.; Kochish, I.I.; Titov, V.Y.; Nikonov, I.N.; Korenyuga, M.V.; Myasnikova, O.V.; Kuvanov, T.K.; Dolgorukova, A.M. Dependence of the Dynamics of Chick Growth in Egg- and Meat-type Breeds on Fractal Bioconsolidation Index of Myogenesis Gene Expression. In Proceedings of the Materials of the 3rd International Scientific and Practical Conference on Molecular Genetic Technologies for Analysis of Gene Expression Related to Animal Productivity and Disease Resistance, Moscow, Russia, 29 September 2021; pp. 206–215. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Titov, V.Y. The enzymatic technologies open new possibilities for studying nitric oxide (NO) metabolism in living systems. Curr. Enzym. Inhib. 2011, 7, 56–70. [Google Scholar] [CrossRef]
- Titov, V.Y.; Kosenko, O.V.; Starkova, E.S.; Kondratov, G.V.; Borkhunova, E.N.; Petrov, V.A.; Osipov, A.N. Enzymatic sensor detects some forms of nitric oxide donors undetectable by other methods in living tissues. Bull. Exp. Biol. Med. 2016, 162, 107–110. [Google Scholar] [CrossRef] [PubMed]
- Leek, J.T.; Scharpf, R.B.; Bravo, H.C.; Simcha, D.; Langmead, B.; Johnson, W.E.; Geman, D.; Baggerly, K.; Irizarry, R.A. Tackling the widespread and critical impact of batch effects in high-throughput data. Nat. Rev. Genet. 2010, 11, 733–739. [Google Scholar] [CrossRef] [PubMed]
- RStudio Team. RStudio Builds: 2022.07.1+554; RStudio 2022.07 “Spotted Wakerobin”, built on 23 July 2022; Posit Software, PBC: Boston, MA, USA; Available online: https://dailies.rstudio.com/version/2022.07.1+554/ (accessed on 29 December 2022).
- RDocumentation. lm: Fitting Linear Models. stats (version 3.6.2). RDocumentation. Available online: https://www.rdocumentation.org/packages/stats/versions/3.6.2/topics/lm (accessed on 29 December 2022).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.L.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D. ggplot2: Create Elegant Data Visualisations Using the Grammar of Graphics, Version 3.3.5; The Comprehensive R Archive Network (CRAN); Vienna University of Economics and Business, Institute for Statistics and Mathematics: Vienna, Austria, 2021; Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 29 December 2022).
- Pedersen, T.L. ggplot2. Version 3.3.5. RDocumentation. 2021. Available online: https://www.rdocumentation.org/packages/ggplot2/versions/3.3.5 (accessed on 29 December 2022).
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix. Version 0.90. 2021. Available online: https://github.com/taiyun/corrplot (accessed on 29 December 2022).
- Lv, X.; Zhang, M.; Li, X.; Ye, R.; Wang, X. Transcriptome profiles reveal the crucial roles of auxin and cytokinin in the “shoot branching” of Cremastra appendiculata. Int. J. Mol. Sci. 2018, 19, 3354. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Song, J.; Qiao, M.; Zhao, X.; Li, R.; Jiao, J.; Sun, Q. Long noncoding RNA expression profile and functional analysis in psoriasis. Mol. Med. Rep. 2019, 19, 3421–3430. [Google Scholar] [CrossRef]
- Sheluhin, O.I.; Magomedova, D.I. Analysis of methods for calculating the fractal dimension of color and grayscale images. H&ES Res. 2017, 9, 6–16. [Google Scholar]
- Coman, S.; Boldisor, C. Special Issue “Fractional Order Modeling in Interdisciplinary Applications” Fractal Fract., EISSN 2504–3110, Published by MDPI (Basel, Switzerland). 2022. Available online: https://www.mdpi.com/journal/fractalfract/special_issues/fractional_order_modeling (accessed on 29 December 2022).
- Ling, H.; Samarasinghe, S.; Kulasiri, D. Novel recurrent neural network for modelling biological networks: Oscillatory p53 interaction dynamics. Biosystems 2013, 114, 191–205. [Google Scholar] [CrossRef]
- Zhao, X.M.; Li, S. HISP: A hybrid intelligent approach for identifying directed signaling pathways. J. Mol. Cell. Biol. 2017, 9, 453–462. [Google Scholar] [CrossRef]
- Bhalla, U.S.; Iyengar, R. Emergent properties of networks of biological signaling pathways. Science 1999, 283, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Steinacher, A.; Bates, D.G.; Akman, O.E.; Soyer, O.S. Nonlinear dynamics in gene regulation promote robustness and evolvability of gene expression levels. PLoS ONE 2016, 11, e0153295. [Google Scholar] [CrossRef] [PubMed]
- Bulgakov, N.G.; Levich, A.P. Description, origin and using of rank distribution in ecology of communities. Vestn. Mosk. Univ. Seriya 16 Biol. 2005, 1, 18–24. [Google Scholar]
- Motomura, I. On the statistical treatment of assemblages. Zool. Mag. 1932, 44, 379–383. [Google Scholar]
- Bulgakov, N.G.; Levich, A.P.; Maksimov, V.N. Regional ecological control based on biotic and abiotic monitoring data. In Environmental Monitoring. Methods of Biological and Physico-Chemical Monitoring: Textbook; Gelashvili, D.B., Ed.; Part 5; Publishing House of Nizhny Novgorod University: Nizhny Novgorod, Russia, 2003; pp. 93–259. [Google Scholar]
- Drakare, S.; Lennon, J.J.; Hillebrand, H. The imprint of the geographical, evolutionary and ecological context on species-area relationships. Ecol. Lett. 2006, 9, 215–227. [Google Scholar] [CrossRef]
- Weron, R. Estimating long-range dependence: Finite sample properties and confidence intervals. Physica A Stat. Mech. Appl. 2002, 312, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Vanin, A.F. Dinitrosyl iron complexes with thiol-containing ligands as a “working form” of endogenous nitric oxide. Nitric Oxide 2016, 54, 15–29. [Google Scholar] [CrossRef]
- Vanin, A.F.; Borodulin, R.R.; Mikoyan, V.D. Dinitrosyl iron complexes with natural thiol-containing ligands in aqueous solutions: Synthesis and some physico-chemical characteristics (A methodological review). Nitric Oxide 2017, 66, 1–9. [Google Scholar] [CrossRef]
- Tarpey, M.M.; Wink, D.A.; Grisham, M.B. Methods for detection of reactive metabolites of oxygen and nitrogen: In vitro and in vivo considerations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 286, R431–R444. [Google Scholar] [CrossRef]
- Titov, V.Y.; Osipov, A.N. Nitrite and nitroso compounds can serve as specific catalase inhibitors. Redox Rep. 2017, 22, 91–97. [Google Scholar] [CrossRef]
- Titov, V.Y.; Vinnikova, E.Z.; Akimova, N.S.; Fisinin, V.I. Nitric oxide (NO) in bird embryogenesis: Physiological role and ability of practical use. Worlds Poult. Sci. J. 2012, 68, 83–96. [Google Scholar] [CrossRef]
- Severina, I.S.; Bussygina, O.G.; Pyatakova, N.V.; Malenkova, I.V.; Vanin, A.F. Activation of soluble guanylate cyclase by NO donors—S-nitrosothiols, and dinitrosyl-iron complexes with thiol-containing ligands. Nitric Oxide 2003, 8, 155–163. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Code | Type of Breed Utility 1 | Origin |
|---|---|---|---|
| Broiler | BR | Meat | Russia |
| White Cornish | WC | Meat | England/Russia |
| Plymouth Rock White | PRW | Dual purpose (meat-egg type) | USA/Russia |
| Yurlov Crower | YC | Dual purpose (meat-egg type) | Russia |
| Brahma Buff | BB | Dual purpose (egg-meat type) | USA/India |
| Orloff Mille Fleur | OMF | Dual purpose (meat-egg type) | Russia |
| Layer | LR | Egg | The Netherlands |
| Uzbek Game (or Kulangi) | UG | Game | Uzbekistan |
| Muscles | Genes * | Breeds | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Broiler | White Cornish | Plymouth Rock White | Yurlov Crower | Brahma Buff | Orloff Mille Fleur | Layer | Uzbek Game | ||
| Breast | MSTN | 11.55 | 4.89 | 6.59 | 121.9 | 41.07 | 2.41 | 4.72 | 1.18 |
| GHR | 6.63 | 5.62 | 4.35 | 69.1 | 31.78 | 3.32 | 4.79 | 2.51 | |
| MEF2C | 6.59 | 2.91 | 4 | 302.3 | 219.8 | 2.33 | 4.14 | 1.45 | |
| MYOD1 | 11.31 | 2.19 | 2.87 | −7.11 | −25.46 | 16.11 | 4.59 | −81.01 | |
| MYOG | 7.46 | −4.32 | 78.25 | 2.04 | −1.95 | 5.58 | 1.03 | −106.9 | |
| MYH1 | −41,760.00 | −16.22 | −24.42 | 1.07 | −1.73 | −16,270.00 | −29.45 | −11,990.00 | |
| MYF5 | −7.57 | −685.02 | −4.76 | −5.90 | −8.57 | −37.53 | −66.26 | −4.47 | |
| Thigh | MSTN | 3.86 | 4.03 | 4.5 | 46.53 | 8.86 | −1.28 | 1.25 | 3.25 |
| GHR | 3.07 | 3.05 | 2.95 | 26.1 | 3.72 | 1.62 | 1.31 | 4.92 | |
| MEF2C | 2.36 | −1.69 | −1.02 | 494.56 | 63.39 | 1.78 | 2.46 | −4.79 | |
| MYOD1 | 18.77 | −12.13 | 13.18 | 28.44 | 8.78 | 6.06 | 1.08 | −13.93 | |
| MYOG | 6.73 | −4640.29 | 1.39 | 1.39 | −2.70 | 3.63 | −78.25 | −118.60 | |
| MYH1 | −10,020.00 | −335.46 | −115.36 | 2.3 | 1.37 | −8481.00 | 6.23 | −17,560.00 | |
| MYF5 | −6.45 | −25,531.63 | −33.36 | 195.36 | 38.02 | −18.90 | −87.43 | −2.43 | |
| Breed | Body Weight, g | Growth Rate | E7 Levels of NO Metabolites | Breed Type Category (ANOVA, p < 0.05) | K(br) | K(th) | MGEI | FC (GHR) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 Day | 14 Days | 28 Days | 2 Weeks | 4 Weeks | NOD, µM | NOD Category (ANOVA, p > 0.05) | Nitrate, µM | Nitrate Category (ANOVA, p > 0.05) | NO Oxidation, % | NO Oxidation Category (ANOVA, p > 0.05) | ||||||
| Broiler | 47.5 | 305.0 | 1157.0 | 6.42 | 24.36 | 3.3 | Low | 145.4 | High | 98.1 | High | Meat | 0.012 | 0.082 | 0.147 | 6.63 |
| White Cornish | 49.3 | 291.7 | 1287.5 | 5.92 | 26.12 | 9.5 | Low | 152.2 | High | 96.9 | High | Meat | 0.037 | 0.233 | 0.159 | 5.62 |
| Plymouth Rock White | 44.9 | 265.2 | 1058.4 | 5.91 | 23.57 | 141.8 | High | 0 | Low | 2.6 | Low | Meat | 0.061 | 0.072 | 0.853 | 4.35 |
| Yurlov Crower | 39.0 | 101.3 | 240.0 | 2.60 | 6.15 | 149.6 | High | 0 | Low | 2.0 | Low | No | 0.316 | 0.584 | 0.541 | 69.1 |
| Brahma Buff | 38.2 | 107.5 | 241.7 | 2.81 | 6.33 | 36.0 | Low | 100.0 | High | 74.1 | High | No | 0.119 | 0.151 | 0.788 | 31.78 |
| Orloff Mille Fleur | 35.5 | 93.1 | 167.8 | 2.62 | 4.73 | 131.5 | High | 0 | Low | 2.1 | Low | No | 0.096 | 0.057 | 1.684 | 3.32 |
| Layer | 42.4 | 79.8 | 222.4 | 1.88 | 5.25 | 138.9 | High | 0 | Low | 2.4 | Low | No | 0.070 | 0.026 | 2.735 | 4.79 |
| Uzbek Game | 41.3 | 92.4 | 223.7 | 2.24 | 5.42 | 8.8 | Low | 143.4 | High | 96.9 | High | No | 0.300 | 0.061 | 4.905 | 2.51 |
| Breed | Breast | Thigh | MGEFDI ** | ||
|---|---|---|---|---|---|
| D(br) | N * | D(th) | N | ||
| Broiler | 0.400 | [1:6] | 0.954 | [1:4] | 0.419 |
| White Cornish | 1.289 | [1:6] | 11.211 | [3:7] | 0.115 |
| Plymouth Rock White | 0.717 | [1:4] | 1.509 | [1:4] | 0.475 |
| Yurlov Crower | 4.104 | [1:6] | 3.377 | [1:4] | 1.215 |
| Brahma Buff | 2.968 | [2:7] | 2.122 | [1:4] | 1.399 |
| Orloff Mille Fleur | 2.984 | [2:6] | 1.1325 | [1:4] | 2.634 |
| Layer | 3.109 | [1:6] | 1.859 | [1:4] | 1.673 |
| Uzbek Game | 3.740 | [1:6] | 0.632 | [1:4] | 5.915 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kochish, I.I.; Brazhnik, E.A.; Vorobyov, N.I.; Nikonov, I.N.; Korenyuga, M.V.; Myasnikova, O.V.; Griffin, D.K.; Surai, P.F.; Romanov, M.N. Features of Fractal Conformity and Bioconsolidation in the Early Myogenesis Gene Expression and Their Relationship to the Genetic Diversity of Chicken Breeds. Animals 2023, 13, 521. https://doi.org/10.3390/ani13030521
Kochish II, Brazhnik EA, Vorobyov NI, Nikonov IN, Korenyuga MV, Myasnikova OV, Griffin DK, Surai PF, Romanov MN. Features of Fractal Conformity and Bioconsolidation in the Early Myogenesis Gene Expression and Their Relationship to the Genetic Diversity of Chicken Breeds. Animals. 2023; 13(3):521. https://doi.org/10.3390/ani13030521
Chicago/Turabian StyleKochish, Ivan I., Evgeni A. Brazhnik, Nikolai I. Vorobyov, Ilya N. Nikonov, Maxim V. Korenyuga, Olga V. Myasnikova, Darren K. Griffin, Peter F. Surai, and Michael N. Romanov. 2023. "Features of Fractal Conformity and Bioconsolidation in the Early Myogenesis Gene Expression and Their Relationship to the Genetic Diversity of Chicken Breeds" Animals 13, no. 3: 521. https://doi.org/10.3390/ani13030521
APA StyleKochish, I. I., Brazhnik, E. A., Vorobyov, N. I., Nikonov, I. N., Korenyuga, M. V., Myasnikova, O. V., Griffin, D. K., Surai, P. F., & Romanov, M. N. (2023). Features of Fractal Conformity and Bioconsolidation in the Early Myogenesis Gene Expression and Their Relationship to the Genetic Diversity of Chicken Breeds. Animals, 13(3), 521. https://doi.org/10.3390/ani13030521