
Feeding Ecology of the Cuvier’s Gazelle (Gazella cuvieri, Ogilby, 1841) in the Sahara Desert
 ,
,  ,
,  , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
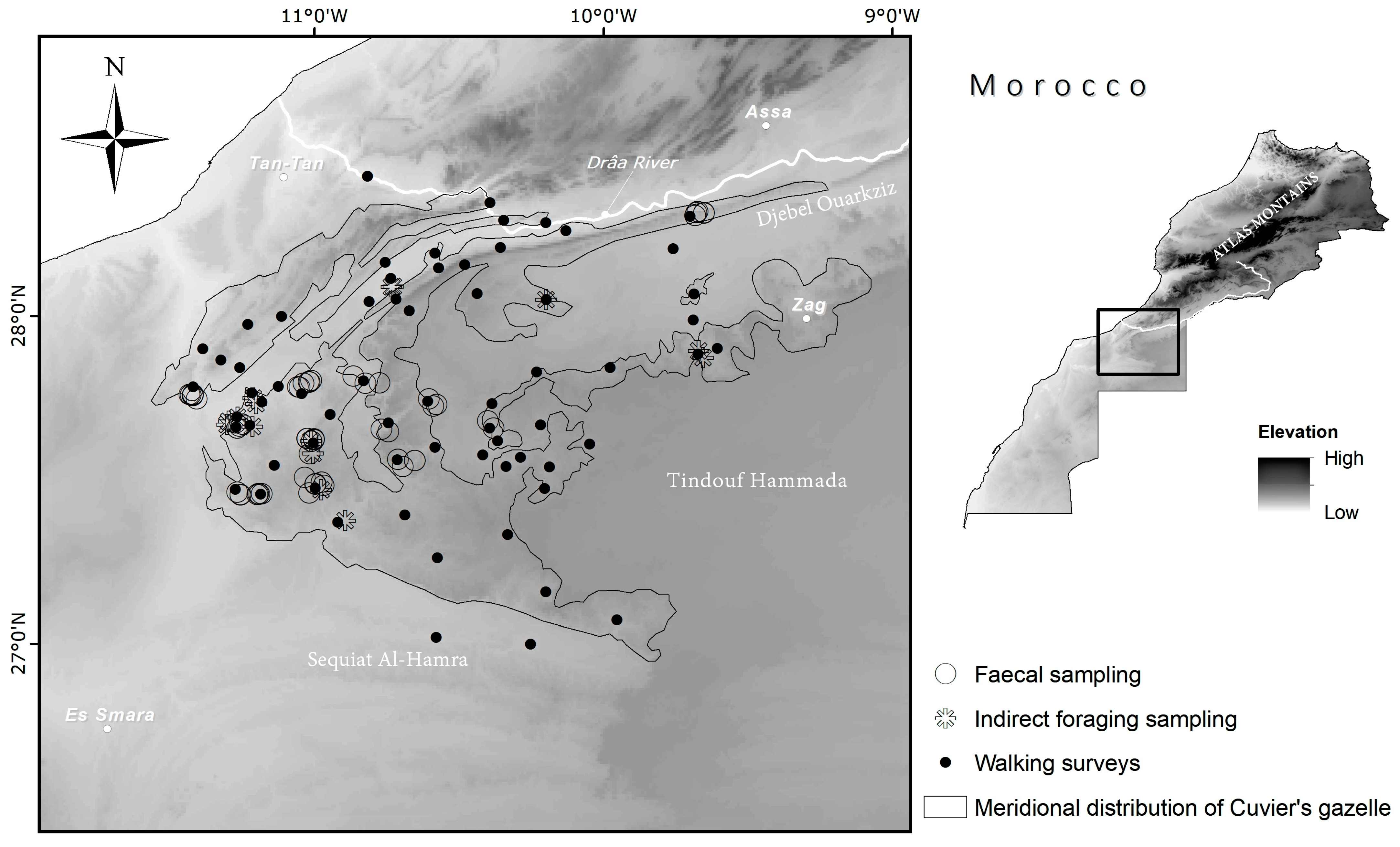
2.1. Study Area
2.2. Data Collection
2.3. Faecal Cuticle Microhistological Analysis of FSs
2.4. Diet Quality
2.5. Environmental Descriptors
2.6. Data Analyses
3. Results
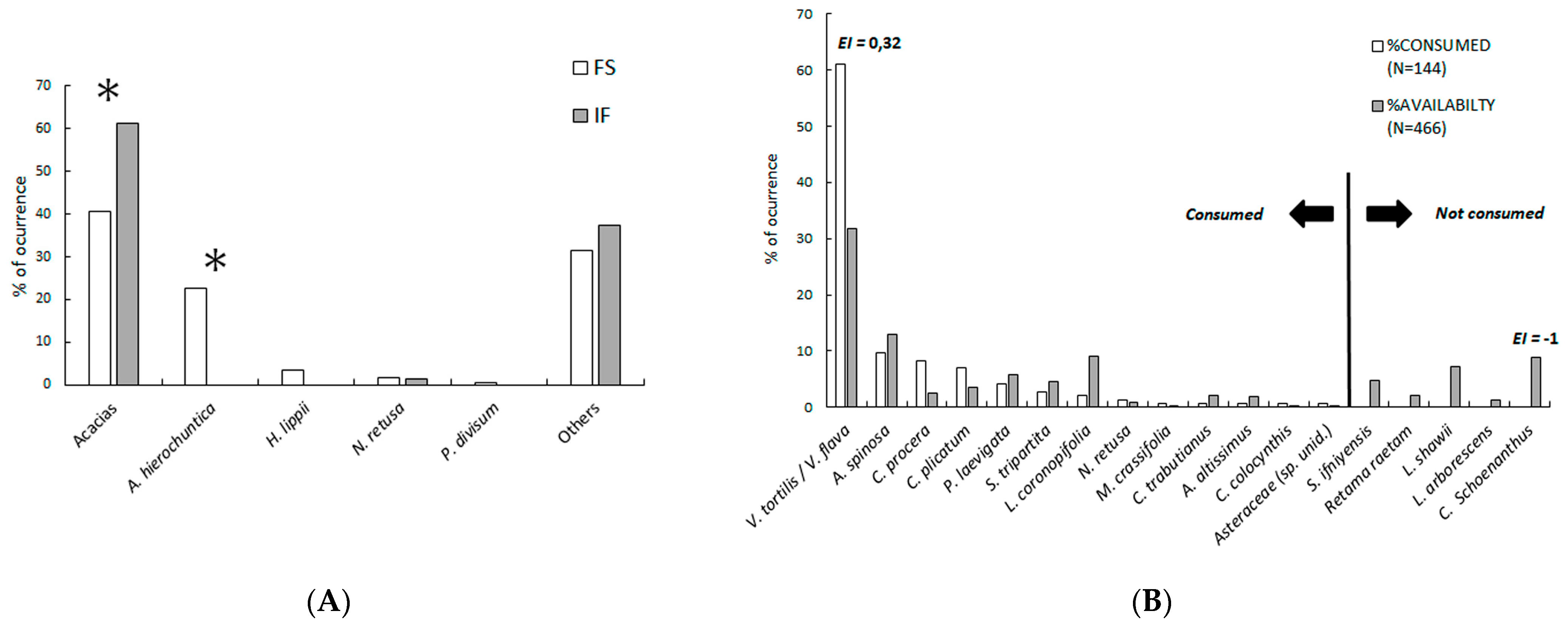
3.1. Diet Composition and Feeding Behaviour
3.2. Effects of Environmental Factors
4. Discussion
4.1. Limitations of the Study
4.2. Feeding Strategy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UNEP. Global Deserts Outlook; UNEP (United Nations Environmental Programme): Nairobi, Kenya, 2006; pp. 4–6. [Google Scholar]
- Le Houérou, H.N. The desert and arid zones of Northern Africa. In Ecosystems of the World. Hot Deserts and Arid Shrublands; Evenari, M., Noy-Meir, E., Goodall, D.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1986; Volume 12B, pp. 101–147. [Google Scholar]
- Whitford, W.G.; Duval, B.D. Ecology of Desert Systems, 2nd ed.; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- IUCN SSC Antelope Specialist Group. Addax nasomaculatus. In The IUCN Red List of Threatened Species 2016: E.T512A50180603; Available online: https://doi.org/10.2305/IUCN.UK.2016-2.RLTS.T512A50180603.en (accessed on 18 May 2022).
- Belbachir, F.; Pettorelli, N.; Wacher, T.; Belbachir-Bazi, A.; Durant, S.M. Monitoring Rarity: The Critically Endangered Saharan Cheetah as a Flagship Species for a Threatened Ecosystem. PLoS ONE 2015, 10, e0115136. [Google Scholar] [CrossRef]
- Durant, S.M.; Wacher, T.; Bashir, S.; Woodroffe, R.; De Ornellas, P.; Ransom, C.; Newby, J.; Abáigar, T.; Abdelgadir, M.; El Alqamy, H.; et al. Fiddling in Biodiversity Hotspots While Deserts Burn? Collapse of the Sahara’s Megafauna. Divers. Distrib. 2014, 20, 114–122. [Google Scholar] [CrossRef]
- Newby, J.; Wacher, T.; Durant, S.M.; Pettorelli, N.; Gilbert, T. Desert Antelopes on the Brink. In Antelope Conservation: From Diagnosis to Action; Bro-Jørgensen, J., Mallon, D.P., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 253–279. [Google Scholar] [CrossRef]
- Grettenberger, J.F.; Newby, J.E. The Status and Ecology of the Dama Gazelle in the Aïr and Ténéré National Nature Reserve. Biol. Conserv. 1986, 38, 207–216. [Google Scholar] [CrossRef]
- Stabach, J.A.; Rabeil, T.; Turmine, V.; Wacher, T.; Mueller, T.; Leimgruber, P. On the Brink of Extinction-Habitat Selection of Addax and Dorcas Gazelle across the Tin Toumma Desert, Niger. Divers. Distrib. 2017, 23, 581–591. [Google Scholar] [CrossRef]
- Herrera-Sánchez, F.J.; Gil-Sánchez, J.M.; Álvarez, B.; Cancio, I.; de Lucas, J.; Arredondo, Á.; Díaz-Portero, M.Á.; Rodríguez-Siles, J.; Sáez, J.M.; Pérez, J.; et al. Identifying Priority Conservation Areas in a Saharan Environment by Highlighting the Endangered Cuvier’s Gazelle as a Flagship Species. Sci. Rep. 2020, 10, 8241. [Google Scholar] [CrossRef] [PubMed]
- Brito, J.C.; Durant, S.M.; Pettorelli, N.; Newby, J.; Canney, S.; Algadafi, W.; Rabeil, T.; Crochet, P.-A.; Pleguezuelos, J.M.; Wacher, T.; et al. Armed Conflicts and Wildlife Decline: Challenges and Recommendations for Effective Conservation Policy in the Sahara-Sahel. Conserv. Lett. 2018, 11, e12446. [Google Scholar] [CrossRef]
- Brito, J.C.; Del Barrio, G.; Stellmes, M.; Pleguezuelos, J.M.; Saarinen, J. Drivers of Change and Conservation Needs for Vertebrates in Drylands: An Assessment from Global Scale to Sahara-Sahel Wetlands. Eur. Zool. J. 2021, 88, 1103–1129. [Google Scholar] [CrossRef]
- Reynolds, J.F.; Kemp, P.R.; Ogle, K.; Fernández, R.J. Modifying the “pulse-reserve” Paradigm for Deserts of North America: Precipitation Pulses, Soil Water, and Plant Responses. Oecologia 2004, 141, 194–210. [Google Scholar] [CrossRef]
- Clauss, M.; Steuer, P.; Müller, D.W.H.; Codron, D.; Hummel, J. Herbivory and Body Size: Allometries of Diet Quality and Gastrointestinal Physiology, and Implications for Herbivore Ecology and Dinosaur Gigantism. PLoS ONE 2013, 8, e68714. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, R.R.; Stewart, D.R.M. Grazer or browser: A classification based on the stomach-structure and feeding habits of east African ruminants. Mammalia 1972, 36, 226–240. [Google Scholar] [CrossRef]
- Hofmann, R.R. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: A comparative view of their digestive system. Oecologia 1989, 78, 443–457. [Google Scholar] [CrossRef]
- Watson, L.H.; Owen-Smith, N. Diet Composition and Habitat Selection of Eland in Semi-Arid Shrubland. Afr. J. Ecol. 2000, 38, 130–137. [Google Scholar] [CrossRef]
- Grettenberger, J. Ecology of the Dorcas Gazelle in Northern Niger. Mammalia 1987, 51, 527–536. [Google Scholar] [CrossRef]
- Baharav, D. Food Habits of the Mountain Gazelle in Semi-Arid Habitats of Eastern Lower Galilee, Israel. J. Arid Environ. 1981, 4, 63–69. [Google Scholar] [CrossRef]
- Beudels, R.C.; Devillers, P.; Lafontaine, R.M.; Devillers-Terschuren, M.O. Sahelo-Saharan Antelopes: Status and Perspectives, 2nd ed.; Report on the conservation status of the six Sahelo-Saharan Antelopes, CMS SSA Concerted Action; CMS Technical Series Publication N°10; UNEP/CMS Secretariat: Bonn, Germany, 2005; pp. 39–56. [Google Scholar]
- Gil-Sánchez, J.M.; Herrera-Sánchez, F.J.; Álvarez, B.; Arredondo, Á.; Bautista, J.; Cancio, I.; Castillo, S.; Díaz-Portero, M.Á.; de Lucas, J.; McCain, E.; et al. Evaluating Methods for Surveying the Endangered Cuvier’s Gazelle Gazella cuvieri in Arid Landscapes. Oryx 2017, 51, 648–655. [Google Scholar] [CrossRef]
- IUCN. Conservation Strategy and Action Plan for Cuvier’s Gazelle (Gazella cuvieri) in North Africa 2017–2026; IUCN: Gland, Switzerland; Malaga, Spain, 2018. [Google Scholar]
- IUCN SSC Antelope Specialist Group. Gazella cuvieri. In The IUCN Red List of Threatened Species 2016: E.T8967A50186003.26; Available online: https://doi.org/10.2305/IUCN.UK.2016-2.RLTS.T8967A50186003.en (accessed on 2 February 2022).
- Abaigar, T.; Cano, M.; Sakkouhi, M. Evaluation of Habitat Use of a Semi-Captive Population of Cuvier’s Gazelles Gazella cuvieri Following Release in Boukornine National Park, Tunisia. Acta Theriol. 2005, 50, 405–415. [Google Scholar] [CrossRef]
- Bounaceur, F.; Boualem, A.; Benamor, N.; Fellous, A.; Benkheira, A.; Bissaad, F.Z.; Aulagnier, S. Updated Distribution and Local Abundance of the Endangered Cuvier’s Gazelle (Mammalia, Bovidae) in Algeria. Folia Zool. 2016, 65, 233–238. [Google Scholar] [CrossRef]
- Breslau, B.; Polak, T.; Shalmon, B.; Groner, E. Evidence of Browsing Pressure on the Critically Endangered Acacia Gazelle (Gazella acaciae). J. Arid Environ. 2020, 173, 104019. [Google Scholar] [CrossRef]
- Benamor, N.; Bounaceur, F.; Baha, M.; Aulagnier, S. First Data on the Seasonal Diet of the Vulnerable Gazella cuvieri (Mammalia: Bovidae) in the Djebel Messaâd Forest, Northern Algeria. Folia Zool. 2019, 68, 253. [Google Scholar] [CrossRef] [Green Version]
- Benamor, N.; Bounaceur, F.; Aulagnier, S. Dietary Overlap of Wild Ungulate Cuvier’s Gazelle with Livestock (Sheep and Goats) in Djebel Messaâd Forest, Algeria. Russ. J. Theriol. 2021, 20, 31–43. [Google Scholar] [CrossRef]
- Hanane, S.; Amhaouch, Z. Seasonal Pattern in Browsing Trees and Shrubs by Reintroduced Cuvier’s Gazelles: A Study in Morocco with Management Implications. Eur. J. Wildl. Res. 2021, 67, 53. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. Bioscience 2001, 51, 933. [Google Scholar] [CrossRef]
- Le Houérou, H.N. Climate, Flora and Fauna Changes in the Sahara over the Past 500 Million Years. J. Arid Environ. 1997, 37, 619–647. [Google Scholar] [CrossRef]
- Casañas, G.-T.F. La Macaronesia: Consideraciones geológicas, biogeográficas y paleoecológicas. In Ecología y Cultuzra en Canarias; Fernández-Palacios Martínez, J.M., Ed.; Universidad de La Laguna: Santa Cruz de Tenerife, Spain, 1999; pp. 39–64. [Google Scholar]
- Ozenda, P. Flore et Végétation du Sahara; Centre National de La Recherche Scientifique: Paris, France, 1991. [Google Scholar]
- Aulagnier, S.; Cuzin, F.; Thévenot, M. Mammifères Sauvages du Maroc. Peuplement, Répartition, Ecologie; Société Française Pour l’Etude et La Protection des Mammifères: Paris, France, 2017. [Google Scholar]
- García-Cardenete, L.; Pleguezuelos, J.M.; Brito, J.C.; Jiménez-Cazalla, F.; Pérez-García, M.T.; Santos, X. Water Cisterns as Death Traps for Amphibians and Reptiles in Arid Environments. Environ. Conserv. 2014, 41, 341–349. [Google Scholar] [CrossRef]
- Garnick, S.; Barboza, P.S.; Walker, J.W. Assessment of Animal-Based Methods Used for Estimating and Monitoring Rangeland Herbivore Diet Composition. Rangel. Ecol. Manag. 2018, 71, 449–457. [Google Scholar] [CrossRef]
- Ait Baamrane, M.A.; Shehzad, W.; Ouhammou, A.; Abbad, A.; Naimi, M.; Coissac, E.; Taberlet, P.; Znari, M. Assessment of the Food Habits of the Moroccan Dorcas Gazelle in M’Sabih Talaa, West Central Morocco, Using the TrnL Approach. PLoS ONE 2012, 7, e35643. [Google Scholar] [CrossRef]
- Silva, T.L.; Godinho, R.; Castro, D.; Abáigar, T.; Brito, J.C.; Alves, P.C. Genetic Identification of Endangered North African Ungulates Using Noninvasive Sampling. Mol. Ecol. Resour. 2015, 15, 652–661. [Google Scholar] [CrossRef]
- Schulz, E.; Fraas, S.; Kaiser, T.M.; Cunningham, P.L.; Ismail, K.; Wronski, T. Food Preferences and Tooth Wear in the Sand Gazelle (Gazella marica). Mamm. Biol. 2013, 78, 55–62. [Google Scholar] [CrossRef]
- Wronski, T.; Schulz-Kornas, E. The Farasan Gazelle—A Frugivorous Browser in an Arid Environment? Mamm. Biol. 2015, 80, 87–95. [Google Scholar] [CrossRef]
- Rabeil, T.; Beudels-Jamar, R.; Greth, A. Mission in Kellé and Termit—Tin Toumma, February 2010; Report; Sahara Conservation Fund; Available online: http://www.catsg.org/cheetah/05_library/5_4_reports/Mission_Report_Termit_10-02.pdf (accessed on 12 February 2022).
- Trond, H.; Olav, H. Winter Feeding Strategies of Migrating and Nonmigrating Moose. Can. J. Zool. 1993, 71, 1421–1428. [Google Scholar]
- Routledge, R.G. Moose Winter Diet Selection in Central Ontario. Alces 2015, 40, 95–101. [Google Scholar]
- Wacher, T.; Sassi, M.; Guidara, H.; Tahri, J. A Reconnaissance Survey of the Jabbes-Senghar National Park, Tunisia; Technical Report No. 9; SCF Pan Sahara Wildlife Survey; Sahara Conservation Fund: L’Isle, Switzerland, 2011. [Google Scholar]
- Bartolome, J.; Franch, J.; Plaixats, J.; Seligman, N.G. Diet Selection by Sheep and Goats on Mediterranean Heath-Woodland Range. J. Range Manag. 1998, 51, 383. [Google Scholar] [CrossRef]
- Alipayo, D.; Valdez, R.; Holechek, J.L.; Cardenas, M. Evaluation of Microhistological Analysis for Determining Ruminant Diet Botanical Composition. J. Range Manag. 1992, 45, 148. [Google Scholar] [CrossRef]
- Davis, N.E.; Coulson, G.; Forsyth, D.M. Diets of Native and Introduced Mammalian Herbivores in Shrub-Encroached Grassy Woodland, South-Eastern Australia. Wildl. Res. 2008, 35, 684. [Google Scholar] [CrossRef]
- Espunyes, J.; Bartolomé, J.; Garel, M.; Gálvez-Cerón, A.; Fernández Aguilar, X.; Colom-Cadena, A.; Calleja, J.A.; Gassó, D.; Jarque, L.; Lavín, S.; et al. Seasonal Diet Composition of Pyrenean Chamois Is Mainly Shaped by Primary Production Waves. PLoS ONE 2019, 14, e0210819. [Google Scholar] [CrossRef] [PubMed]
- Villamuelas, M.; Fernández, N.; Albanell, E.; Gálvez-Cerón, A.; Bartolomé, J.; Mentaberre, G.; López-Olvera, J.R.; Fernández-Aguilar, X.; Colom-Cadena, A.; López-Martín, J.M.; et al. The Enhanced Vegetation Index (EVI) as a Proxy for Diet Quality and Composition in a Mountain Ungulate. Ecol. Indic. 2016, 61, 658–666. [Google Scholar] [CrossRef]
- Villamuelas, M.; Serrano, E.; Espunyes, J.; Fernández, N.; López-Olvera, J.R.; Garel, M.; Santos, J.; Parra-Aguado, M.Á.; Ramanzin, M.; Fernández-Aguilar, X.; et al. Predicting Herbivore Faecal Nitrogen Using a Multispecies Near-Infrared Reflectance Spectroscopy Calibration. PLoS ONE 2017, 12, e0176635. [Google Scholar] [CrossRef]
- Ramanzin, M.; Párraga Aguado, M.Á.; Ferragina, A.; Sturaro, E.; Semenzato, P.; Serrano, E.; Clauss, M.; Albanell, E.; Cassini, R.; Bittante, G. Methodological Considerations for the Use of Faecal Nitrogen to Assess Diet Quality in Ungulates: The Alpine Ibex as a Case Study. Ecol. Indic. 2017, 82, 399–408. [Google Scholar] [CrossRef]
- Steuer, P.; Südekum, K.-H.; Tütken, T.; Müller, D.W.H.; Kaandorp, J.; Bucher, M.; Clauss, M.; Hummel, J. Does Body Mass Convey a Digestive Advantage for Large Herbivores? Funct. Ecol. 2014, 28, 1127–1134. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Ivlev, V.S. Experimental Ecology of the Feeding of Fishes; Yale University Press: New Haven, CT, USA, 1961. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analyzing Ecological Data; Springer: New York, NY, USA, 2007. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 253–258. [Google Scholar] [CrossRef]
- Schoener, T.W. Theory of Feeding Strategies. Annu. Rev. Ecol. Syst. 1971, 2, 369–404. [Google Scholar] [CrossRef]
- Pyke, G.H.; Pulliam, R.; Charnov, E.L. Optimal Foraging: A Selective Review of Theory and Tests. Q. Rev. Biol. 1977, 52, 137–154. [Google Scholar] [CrossRef]
- Stephens, D.W.; Krebs, J.R. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Futuyma, D.J.; Moreno, G. The evolution of ecological specialization. Annu. Rev. Ecol. Syst. 1988, 19, 207–233. [Google Scholar] [CrossRef]
- Valverde, J.A. Aves del Sahara Español (Estudio Ecológico del Desierto); Consejo Superior de Investigaciones Científicas, Instituto de Estudios Africanos: Madrid, Spain, 1957. [Google Scholar]
- Attum, O.; Ghazali, U.; El Noby, S.K.; Hassan, I.N. The Effects of Precipitation History on the Kilometric Index of Dorcas Gazelles. J. Arid Environ. 2014, 102, 113–116. [Google Scholar] [CrossRef]
- Huffman, B.A. Acacia Gazelle Gazella acaciae. In Handbook of the Mammals of the World, Volume 2: Hoofed Mammals; Wilson, D.E., Mittermeier, R.A., Eds.; Lynx Editions: Barcelona, Spain, 2011; p. 642. [Google Scholar]
- Huffman, B.A. Arabian Desert Gazelle Gazella cora. In Handbook of the Mammals of the World, Volume 2: Hoofed Mammals; Wilson, D.E., Mittermeier, R.A., Eds.; Lynx Editions: Barcelona, Spain, 2011; pp. 642–643. [Google Scholar]
- William, A.; Shamaki, B.U.; Sadiq, A.A.; Abdullahi, A. Phytochemical and Elemental Constituents, Acute Toxicity (LD50) Studies of Aqueous Leaf Extract of Calotropis Procera. World J. Pharm. Sci. 2015, 3, 2321–3086. [Google Scholar]
- Batanouny, K.H. Topography and Plant Life. In Plants in the Deserts of the Middle East. Adaptations of Desert Organisms; Springer: Berlin/Heidelberg, Germany, 2001; pp. 25–29. [Google Scholar] [CrossRef]
- Batanouny, K.H. Adaptation of Desert Plants. In Plants in the Deserts of the Middle East. Adaptations of Desert Organisms; Springer: Berlin/Heidelberg, Germany, 2001; pp. 39–44. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Category | Method 1 |
|---|---|---|---|
| Vachellia flava | Leguminosae | Tree | DO, IF, FS |
| V. tortilis subsp. raddiana | Leguminosae | Tree | IF, FS |
| Anastatica hierochuntica | Brassicaceae | Forbs | FS |
| Argania spinosa | Sapotaceae | Tree | IF |
| Asparagus altissimus | Asparagaceae | Shrub | DO, IF |
| Asteraceae (sp. unidentified) | Asteraceae | Forbs | IF |
| Calotropis procera | Apocynaceae | Tree | IF |
| Citrullus colocynthis | Cucurbitaceae | Forbs | IF |
| Convolvulus trabutianus | Convolvulaceae | Shrub | IF |
| Cullen plicatum | Leguminosae | Shrub | IF |
| Helianthemum lippii | Cistaceae | Shrub | FS |
| Lavandula coronopifolia | Lamiaceae | Shrub | IF |
| Lycium shawii | Solanaceae | Shrub | DO |
| Maerua crassifolia | Capparaceae | Tree | IF |
| Nitraria retusa | Nitrariaceae | Shrub | IF, FS |
| Pennisetum divisum | Poaceae | Grass | FS |
| Periploca laevigata | Apocynaceae | Shrub | DO, IF |
| Searsia tripartita | Anacardiaceae | Shrub | IF |
| Faecal Items 1 | Mean ± SD | Range | |
|---|---|---|---|
| Diet composition | Anastatica hierochuntica | 22.77 ± 23.80 | 0–80.00 |
| Acacias | 41.01 ± 16.42 | 9.00–82.00 | |
| Others | 31.73 ± 13.9 | 6.00–71.00 | |
| Helianthemum lippii | 3.61 ± 8.18 | 0–40.00 | |
| Nitraria retusa | 1.63 ± 4.72 | 0–33.00 | |
| Pennisetum divisum | 0.04 ± 2.91 | 0–22.00 | |
| Diet quality | FNc | 0.18 ± 0.06 | 0.0.79–0.034 |
| NDF | 37.71 ± 4.63 | 27.80–50.70 | |
| ADF | 26.30 ± 3.02 | 19.8–34.3 | |
| ADL | 11.72 ± 2.20 | 7.8–16.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera-Sánchez, F.J.; López, O.; Rodríguez-Siles, J.; Díaz-Portero, M.Á.; Arredondo, Á.; Sáez, J.M.; Álvarez, B.; Cancio, I.; de Lucas, J.; Pérez, J.; et al. Feeding Ecology of the Cuvier’s Gazelle (Gazella cuvieri, Ogilby, 1841) in the Sahara Desert. Animals 2023, 13, 567. https://doi.org/10.3390/ani13040567
Herrera-Sánchez FJ, López O, Rodríguez-Siles J, Díaz-Portero MÁ, Arredondo Á, Sáez JM, Álvarez B, Cancio I, de Lucas J, Pérez J, et al. Feeding Ecology of the Cuvier’s Gazelle (Gazella cuvieri, Ogilby, 1841) in the Sahara Desert. Animals. 2023; 13(4):567. https://doi.org/10.3390/ani13040567
Chicago/Turabian StyleHerrera-Sánchez, F. Javier, Omar López, Javier Rodríguez-Siles, Miguel Ángel Díaz-Portero, Ángel Arredondo, Juan Manuel Sáez, Begoña Álvarez, Inmaculada Cancio, Jesús de Lucas, Joaquín Pérez, and et al. 2023. "Feeding Ecology of the Cuvier’s Gazelle (Gazella cuvieri, Ogilby, 1841) in the Sahara Desert" Animals 13, no. 4: 567. https://doi.org/10.3390/ani13040567
APA StyleHerrera-Sánchez, F. J., López, O., Rodríguez-Siles, J., Díaz-Portero, M. Á., Arredondo, Á., Sáez, J. M., Álvarez, B., Cancio, I., de Lucas, J., Pérez, J., Valenzuela, G., Martínez-Valderrama, J., Sánchez-Cerdá, M., Qninba, A., Virgós, E., Calleja, J. A., Bartolomé, J., Albanell, E., Serrano, E., ... Gil-Sánchez, J. M. (2023). Feeding Ecology of the Cuvier’s Gazelle (Gazella cuvieri, Ogilby, 1841) in the Sahara Desert. Animals, 13(4), 567. https://doi.org/10.3390/ani13040567