Implication of Polyhistidine, a Novel Apoptosis Inhibitor, in Inhibiting Lipopolysaccharide-Induced Apoptosis in Boar Sperm


 , and
, and 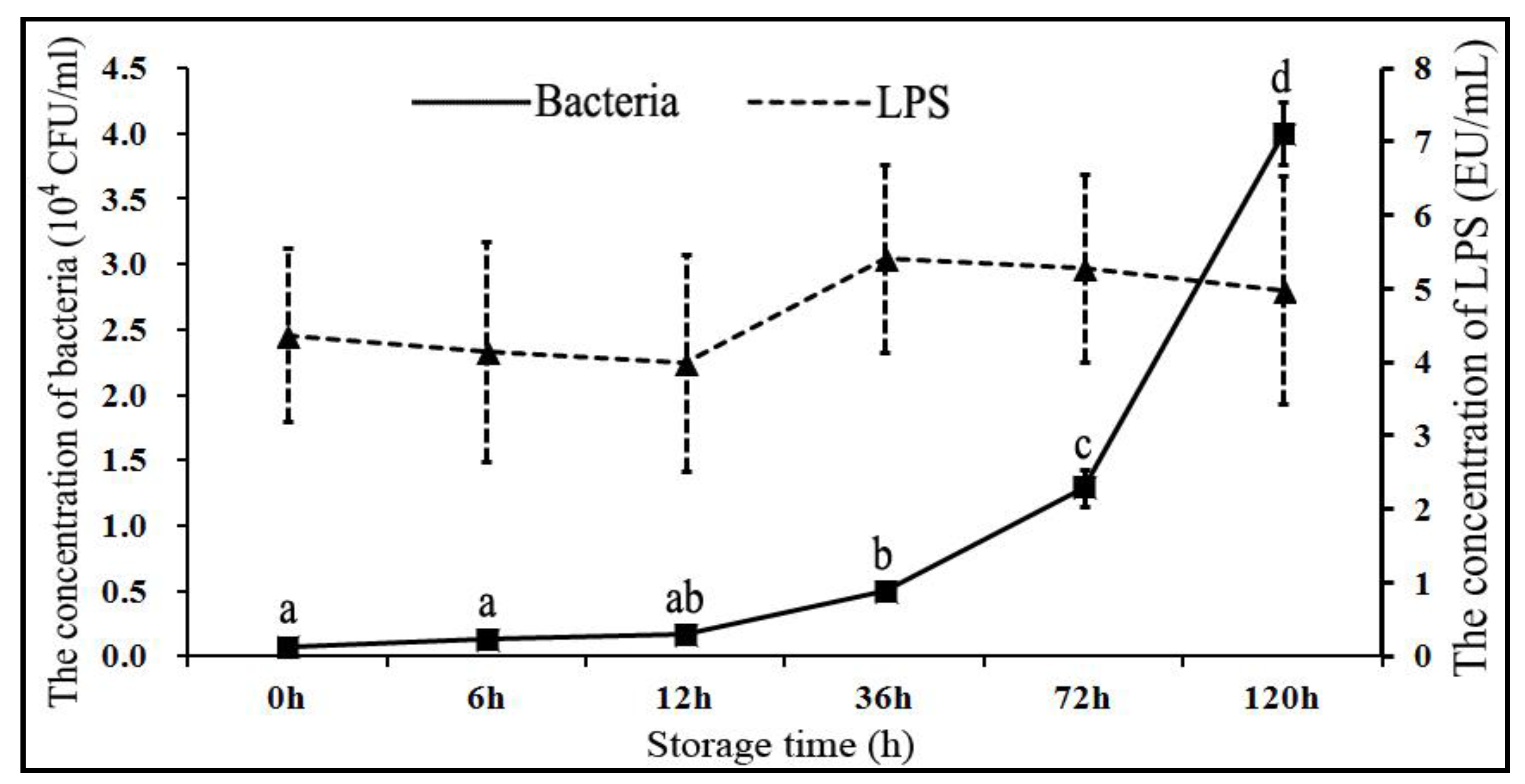{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Extender
2.2. Semen Collection and Preparation
2.3. Experimental Design
2.4. Quantification of Bacterial Colonies
2.5. Detection of LPS in Semen by Limulus Assay
2.6. Semen Quality Assessment
2.7. Detection of Sperm Apoptosis
2.8. Statistical Analysis
3. Results and Discussion
3.1. Changes in Concentration of Bacterial Colonies and LPS Under Liquid Storage Conditions
3.2. Effects of pHis on Boar Sperm Quality
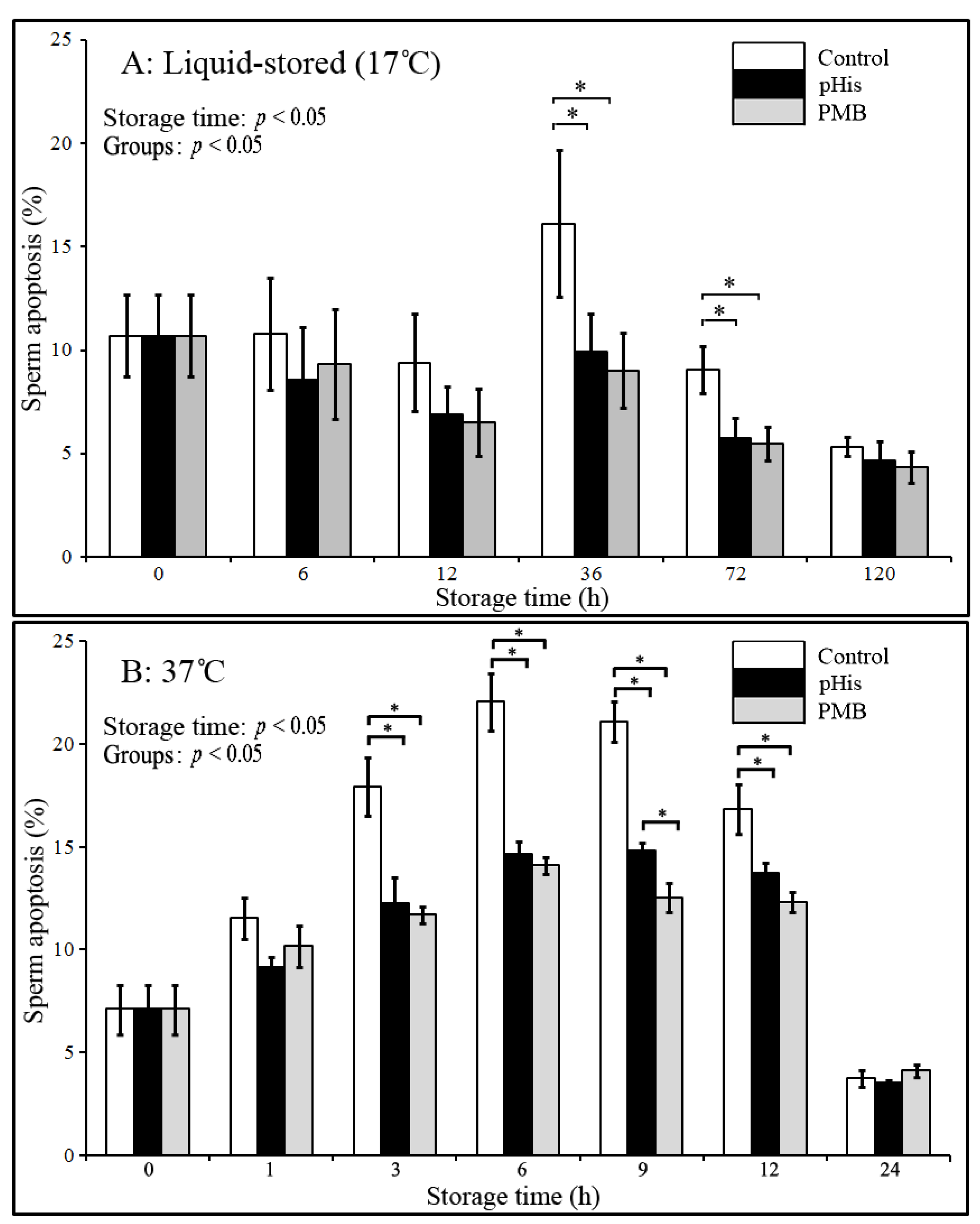
3.3. Implication of Polyhistidine in Inhibiting Sperm Apoptosis
3.4. pHis Inhibited LPS-Induced Sperm Apoptosis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Althouse, G.C.; Pierdon, M.S.; Lu, K.G. Thermotemporal dynamics of contaminant bacteria and antimicrobials in extended porcine semen. Theriogenology 2008, 70, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Sone, M. Investigation on the Control of Bacteria in Boar Semen. Advances in Methods of Fertility Regulation. New York, NY, USA, 1990. Available online: https://www.jstage.jst.go.jp/article/jrd1977/36/5/36_5_23P/_article/-char/en (accessed on 24 September 2019).
- Sanocka-Maciejewska, D.; Ciupińska, M.; Kurpisz, M. Bacterial infection and semen quality. J. Reprod. Immunol. 2005, 67, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Danowski, K.M. Qualitative and Quantitative Investigation of the Germ Content in Boar Semen and the Antibiotic Sensitivity of the Prevailing Germ Spectrum (under the Aspect of Possible Semen Preservation). M.D. Thesis, Tieraerztliche Hochschule Hannover, Hannover, Germany, 1989. [Google Scholar]
- Gączarzewicz, D.; Udała, J.; Piasecka, M.; Błaszczyk, B.; Stankiewicz, T. Bacterial Contamination of Boar Semen and its Relationship to Sperm Quality Preserved in Commercial Extender Containing Gentamicin Sulfate. Pol. J. Vet. Sci. 2016, 19, 451–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagnall, G. An Investigation of the Bacterial Flora of the Preputial Diverticulum and of the Semen of Boars. M.D. Thesis, Universities of Great Britain and Ireland, Great Britain and Ireland, UK, 1987. [Google Scholar]
- Althouse, G.C.; Lu, K.G. Bacteriospermia in extended porcine semen. Theriogenology 2005, 63, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Auroux, M.R.; Jacques, L.; Mathieu, D.; Auer, J. Is the sperm bacterial ratio a determining factor in impairment of sperm motility: An in-vitro study in man with Escherichia coli. Int. J. Androl. 1991, 14, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Gorga, F.; Galdiero, M.; Buommino, E.; Galdiero, E. Porins and lipopolysaccharide induce apoptosis in human spermatozoa. Clin. Diag. Lab. Immunol. 2001, 8, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Mihara, T.; Okazaki, T.; Shitanaka, M.; Kushino, R.; Ikeda, C.; Negishi, H.; Liu, Z.L.; Richards, J.S.; Shimada, M. Toll-like receptors (TLR) 2 and 4 on human sperm recognize bacterial endotoxins and mediate apoptosis. Hum. Reprod. 2011, 26, 2799–2806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nii, T.; Sonoda, Y.; Isobe, N.; Yoshimura, Y. Effects of lipopolysaccharide on the expression of proinflammatory cytokines and chemokines and the subsequent recruitment of immunocompetent cells in the oviduct of laying and molting hens. Poult. Sci. 2011, 90, 2332–2341. [Google Scholar] [CrossRef]
- Liu, S.J.; Shi, Y.; Liu, C.; Zhang, M.; Zuo, Z.C.; Zeng, C.J.; Zhou, G.B.; Xian, H.; Song, T.Z. The upregulation of pro-inflammatory cytokines in the rabbit uterus under the lipopolysaccaride-induced reversible immunoresponse state. Anim. Reprod. Sci. 2017, 176, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; He, Z.B.; Zhang, M.; Chen, S.Y.; Lan, R.X.; Wang, J.; Lai, S.J. Expression pattern of Toll-like receptors (TLRs) in different organs and effects of lipopolysaccharide on the expression of TLR 2 and 4 in reproductive organs of female rabbit. Dev. Comp. Immunol. 2014, 46, 341–348. [Google Scholar] [CrossRef]
- Okazaki, T.; Fujita, Y.; Mihara, T.; Shitanaka, M.; Negishi, H.; Richards, J.S.; Shimada, M. Expression of the Toll-like receptors system that recognize bacterize bacterial infection, controls fertilization ability in mammalian spermatozoa. Reprod. Immunol. Biol. 2009, 24, 174. [Google Scholar]
- Althouse, G.C.; Kuster, C.E.; Clark, S.G.; Weisiger, R.M. Field investigations of bacterial contaminants and their effects on extended porcine semen. Theriogenology 2000, 53, 1167–1176. [Google Scholar] [CrossRef]
- Ginsburg, I. The role of bacteiolysis in the pathophysiology of inflammation, infection and post-infectious sequelae: Review article. APMIS 2002, 110, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Anzar, M.; He, L.; Buhr, M.M.; Kroetsch, T.G.; Pauls, K.P. Sperm apoptosis in fresh and cryopreserved bull semen detected by flow cytometry and its relationship with fertility. Biol. Reprod. 2002, 66, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Eley, A.; Hosseinzadeh, S.; Hakimi, H.; Geary, I.; Pacey, A.A. Apoptosis of ejaculated human sperm is induced by co-incubation with Chlamydia trachomatis lipopolysaccharide. Hum. Reprod. 2005, 20, 2601–2607. [Google Scholar] [CrossRef] [Green Version]
- Bussalleu, E.; Yeste, M.; Sepulvda, L.; Torner, E.; Pinart, E.; Bonet, S. Effects of different concentrations of enterotoxigenic and verotoxigenic E. coli on boar sperm quality. Anim. Reprod. Sci. 2011, 127, 176–182. [Google Scholar] [CrossRef]
- Kiranjeet, K.; Vijay, P. Sperm impairment by sperm agglutinating factor isolated from Escherichia coli: Receptor specific interactions. BioMed. Rese. Inter. 2013. [Google Scholar] [CrossRef]
- Beutler, B. TLR4: Central component of the sole mammalian LPS sensor. Curr. Opin. Immunol. 2000, 12, 20–26. [Google Scholar] [CrossRef]
- Kim, H.M.; Park, B.S.; Kim, J.I.; Kim, S.E.; Lee, J.D.; Oh, S.C.; Enkhbayar, P.; Matsushima, N.; Lee, H.Y.; Yoo, O.J.; et al. Crystal structure of the TLR4-MD-2 complex with bound endotoxin antagonist Eritoran. Cell 2007, 130, 906–917. [Google Scholar] [CrossRef]
- Van-der, L.I.M.; Kersten, G.; Fretz, M.M.; Beuvery, C.; Coos, V.J.; Junginger, H.E. Chitosan microparticles for mucosal vaccination against diphtheria: Oral and nasal efficacy studies in mice. Vaccine 2003, 21, 1400–1408. [Google Scholar] [CrossRef]
- Weidemann, B.; Brade, H.; Rietschel, E.T.; Dziarski, R.; Bazil, V.; Kusumoto, S.; Flad, H.D.; Ulmer, A.J. Soluble peptidoglycan-induced monokine production can be blocked by anti-CD14 monoclonal antibodies and by lipid A partial structures. Infect. Immun. 1994, 62, 4709–4715. [Google Scholar] [PubMed]
- Verbon, A.; Dekkers, P.E.; Ten, H.T.; Hack, C.E.; Pribble, J.P.; Turner, T.; Souza, S.; Axtelle, T.; Hoek, F.J.; Van-Deventer, S.J.; et al. IC14, an anti-CD14 antibody, inhibits endotoxin-mediated symptoms and inflammatory responses in humans. J. Immunol. 2001, 166, 3599–3605. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.E.; Park, Z.Y.; Kim, N.D.; Lee, J.Y. Sulforaphane inhibits the engagement of LPS with TLR4/MD2 complex by preferential binding to Cys133 in MD2. Biochem. Biophys. Res. Commun. 2013, 434, 600–605. [Google Scholar] [CrossRef] [PubMed]
- Le-Roy, D.; Di-Padova, F.; Tees, R.; Lengacher, S.; Landmann, R.; Glauser, M.P.; Calandra, T.; Heumann, D. Monoclonal antibodies to murine lipopolysaccharide (LPS)-binding protein (LBP) protect mice from lethal endotoxemia by blocking either the binding of LPS to LBP or the presentation of LPS/LBP complexes to CD14. J. Immunol. 1999, 162, 7454–7460. [Google Scholar] [PubMed]
- Lee, H.J.; Huang, Y.W.; Chiou, S.H.; Aronstam, R.S. Polyhistidine facilitates direct membrane translocation of cell-penetrating peptide into cells. Sci. Rep. 2019, 9, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Francesco, P.; Matteo, P.; Valentina, C.; Gaetana, D.; Roberto, C. Exploring the LPS/TLR4 signal pathway with small molecules. Biochem. Soc. Trans. 2010, 38, 1390–1395. [Google Scholar] [Green Version]
- Joshua, A.B.; Joseph, J.F. Purification of proteins using polyhistidine affinity tags. Methods Enzymol. 2000, 326, 245–254. [Google Scholar]
- Evans, M.E.; Feola, D.J.; Rapp, R.P. Polymyxin B sulfate and colistin: old antibiotics for emerging multiresistant gram-negative bacteria. Ann. Pharmacother. 1999, 33, 960–967. [Google Scholar] [CrossRef]
- Srimal, S.; Surolia, N.; Balasubramanian, S.; Surolia, A. Titration calorimetric studies to elucidate the specificity of the interactions of polymyxin B with lipopolysaccharides and lipid A. Biochem. J. 1996, 315, 679–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, L.; Aalbers, J.; Grooten, H. Artificial insemination of swine: fecundity of boar semen stored in Beltsville TS (BTS), modified Modena (MM), or MR-A and inseminated on one, three and four days after collection. Reprod. Domest. Anim. 1988, 23, 49–55. [Google Scholar] [CrossRef]
- Elin, R.J.; Robinson, R.A.; Levine, A.S.; Wolff, S.M. Lack of clinical usefulness of the limulus test in the diagnosis of endotoxemia. N. Engl. J. Med. 1975, 293, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Arrighi, C.D.V.; Lucas, H.; Chardonnens, D.; Agostini, A.D. Removal of spermatozoa with externalized phosphatidylserine from sperm preparation in human assisted medical procreation: Effects on viability, motility and mitochondrial membrane potential. Reprod. Biol. Endocrin. 2009, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Morrell, J.; Wallgren, M. Removal of bacteria from boar ejaculates by Single-layer centrifugation can reduce the use of antibiotics in semen extenders. Anim. Reprod. Sci. 2011, 123, 64–69. [Google Scholar] [CrossRef] [PubMed]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, T.; Shi, Y.; Wang, Y.; Qazi, I.H.; Angel, C.; Zhang, M. Implication of Polyhistidine, a Novel Apoptosis Inhibitor, in Inhibiting Lipopolysaccharide-Induced Apoptosis in Boar Sperm. Animals 2019, 9, 719. https://doi.org/10.3390/ani9100719
Song T, Shi Y, Wang Y, Qazi IH, Angel C, Zhang M. Implication of Polyhistidine, a Novel Apoptosis Inhibitor, in Inhibiting Lipopolysaccharide-Induced Apoptosis in Boar Sperm. Animals. 2019; 9(10):719. https://doi.org/10.3390/ani9100719
Chicago/Turabian StyleSong, Tianzeng, Yi Shi, Yangang Wang, Izhar Hyder Qazi, Christiana Angel, and Ming Zhang. 2019. "Implication of Polyhistidine, a Novel Apoptosis Inhibitor, in Inhibiting Lipopolysaccharide-Induced Apoptosis in Boar Sperm" Animals 9, no. 10: 719. https://doi.org/10.3390/ani9100719
APA StyleSong, T., Shi, Y., Wang, Y., Qazi, I. H., Angel, C., & Zhang, M. (2019). Implication of Polyhistidine, a Novel Apoptosis Inhibitor, in Inhibiting Lipopolysaccharide-Induced Apoptosis in Boar Sperm. Animals, 9(10), 719. https://doi.org/10.3390/ani9100719