
Understanding Estimations of Magnitudes: An fMRI Investigation


Abstract
:1. Introduction
1.1. From Continuous Properties of Physical Stimuli to Numerical Processing
1.2. Cognitive Estimation, the Role of Executive Function and Quantity Estimation
1.3. CET of Continuous Magnitude or Discrete Numerical Information: The Usage of Measurement Units
1.4. The Current Study
2. Materials and Methods
2.1. Participants
2.2. Stimuli and Procedure
2.2.1. Behavioral Pilot: Creating CET Questions for the fMRI
2.2.2. fMRI CET Task
2.2.3. fMRI Acquisition
2.2.4. fMRI Analysis
2.2.5. fMRI Multi-Voxel Pattern Analysis (MVPA)
3. Results
3.1. Behavioral Results
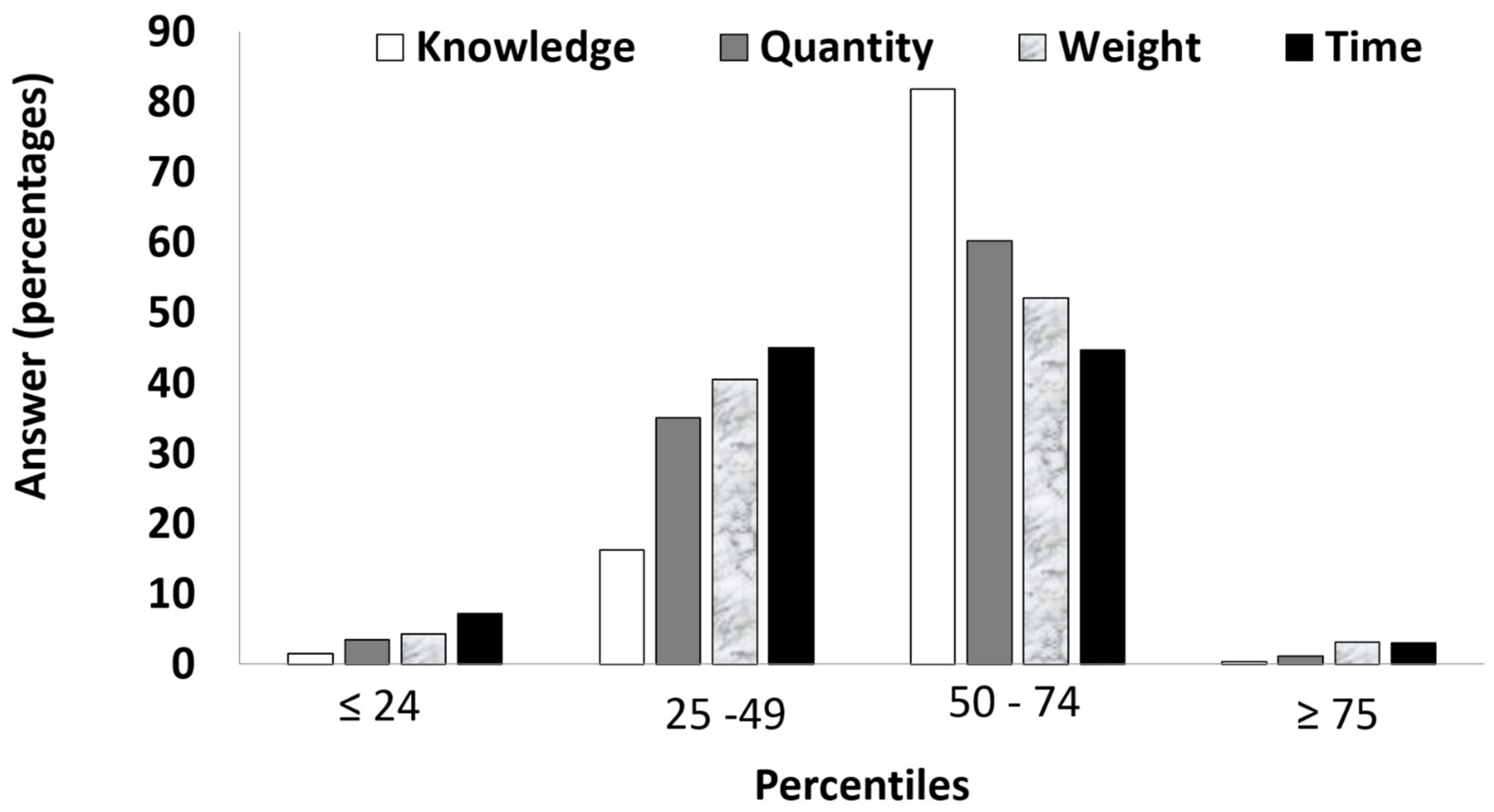
3.1.1. Responses Frequencies
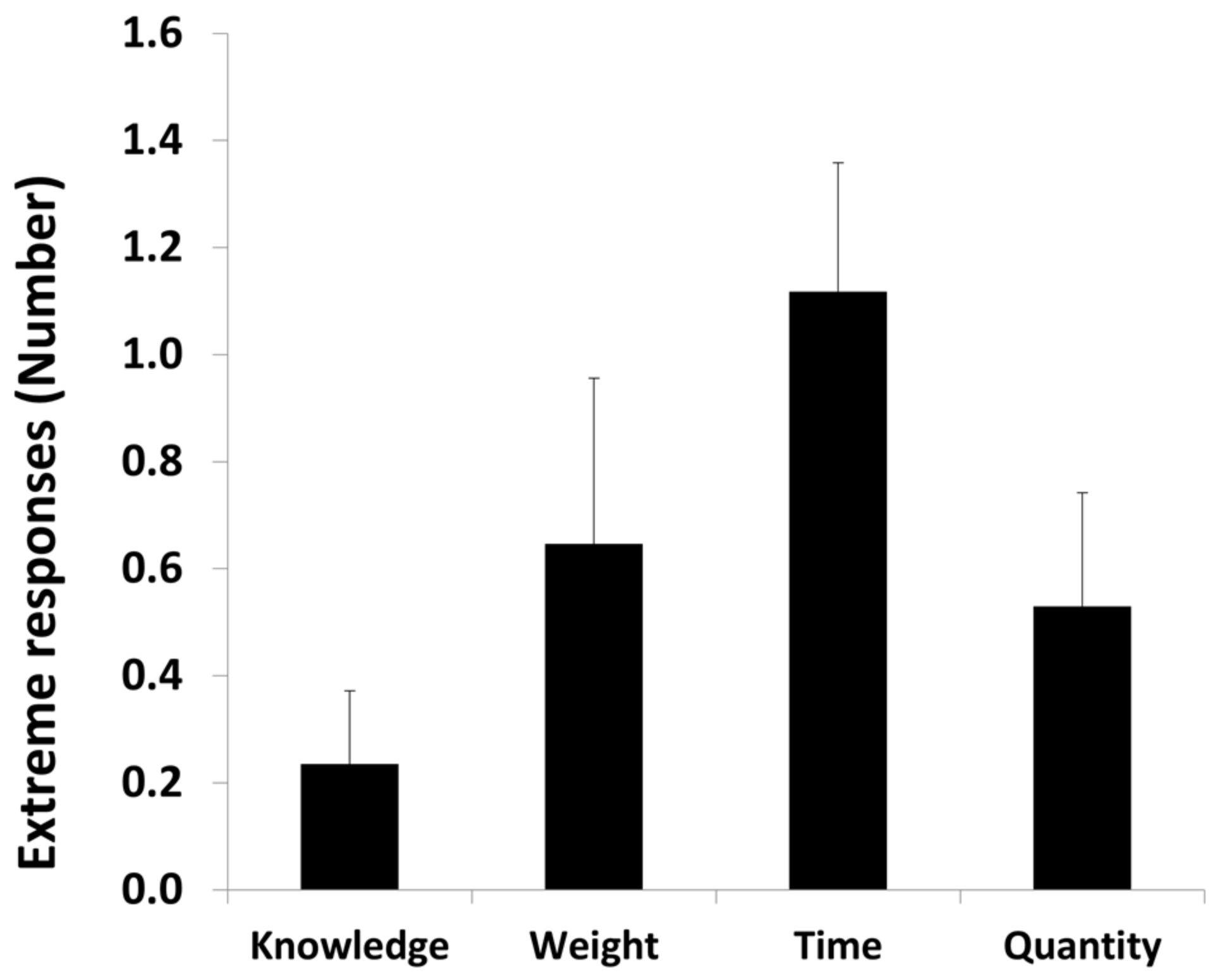
3.1.2. Extreme Responses
3.1.3. RT
3.2. fMRI Results
3.2.1. Whole-Brain Univoxel Analysis
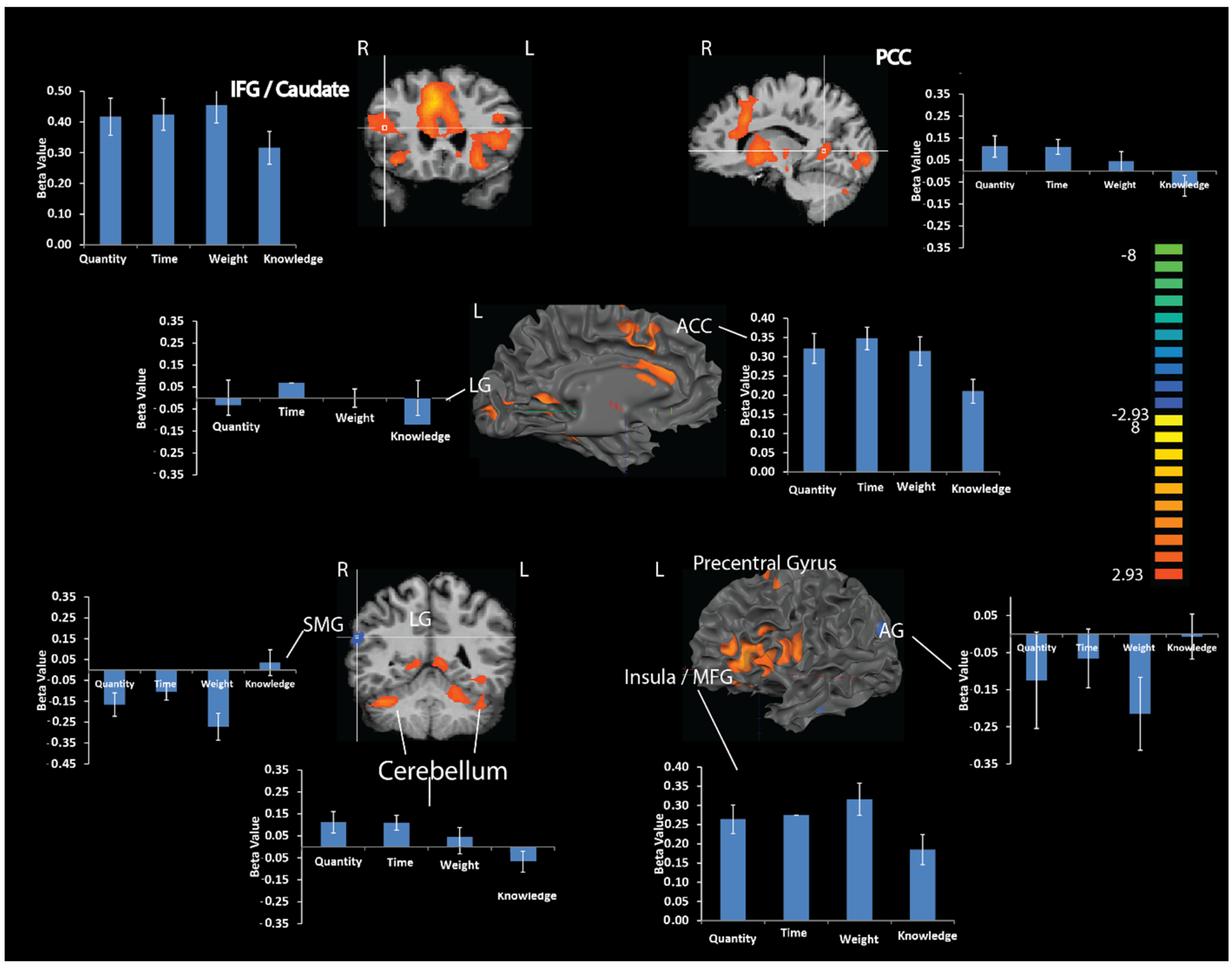
Brain Activations Related to CET
Numerical Estimation vs. Weight
Numerical Estimation vs. Time
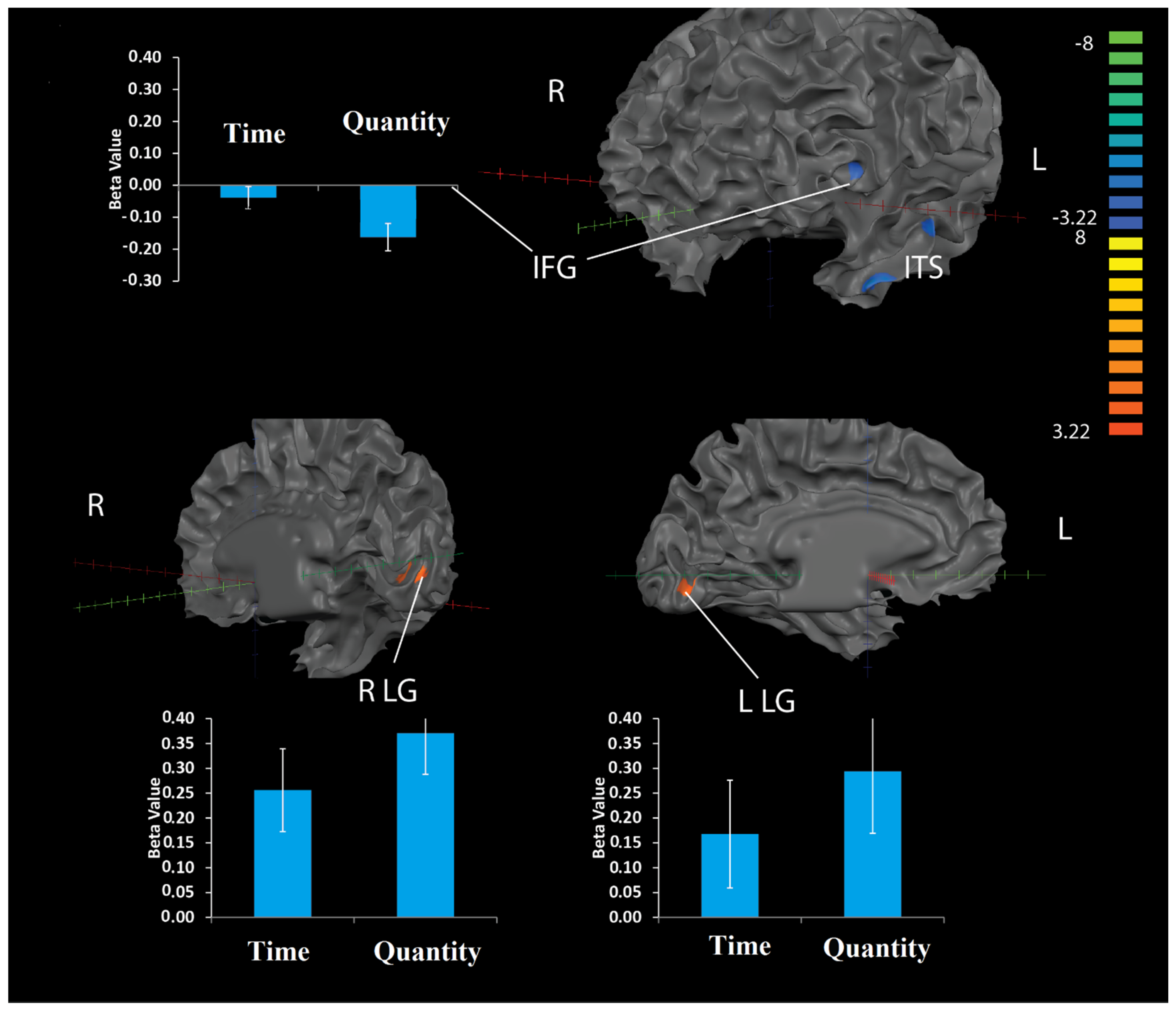
3.2.2. Whole-Brain Multi-Voxel Pattern Analysis (MVPA)
Numerical Estimation vs. Time
Numerical Estimation vs. Weight
4. Discussion
4.1. CET Elicits Activity in the Frontoparietal Network Related to EF and Numerical Cognition
4.2. Distinct Brain Activation to Discrete Numerosity and Continuous Magnitude Estimations
4.3. Predicting Individual Differences in CET
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Halberda, J.; Feigenson, L. Developmental change in the acuity of the “number sense”: The approximate number system in 3-, 4-, 5-, and 6-year-olds and adults. Dev. Psychol. 2008, 44, 1457–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halberda, J.; Ly, R.; Wilmer, J.B.; Naiman, D.Q.; Germine, L. Number sense across the lifespan as revealed by a massive Internet-based sample. Proc. Natl. Acad. Sci. USA 2012, 109, 11116–11120. [Google Scholar] [CrossRef] [Green Version]
- Halberda, J.; Mazzocco, M.M.; Feigenson, L. Individual differences in non-verbal number acuity correlate with maths achievement. Nature 2008, 455, 665–668. [Google Scholar] [CrossRef]
- Ashkenazi, S.; Rosenberg-Lee, M.; Tenison, C.; Menon, V. Weak task-related modulation and stimulus representations during arithmetic problem solving in children with developmental dyscalculia. Dev. Cogn. Neurosci. 2012, 2, S152–S166. [Google Scholar] [CrossRef] [Green Version]
- Cantlon, J.F.; Brannon, E.M.; Carter, E.J.; Pelphrey, K.A. Functional imaging of numerical processing in adults and 4-y-old children. PLoS Biol. 2006, 4, e125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantlon, J.F.; Libertus, M.E.; Pinel, P.; Dehaene, S.; Brannon, E.M.; Pelphrey, K.A. The neural development of an abstract concept of number. J. Cogn. Neurosci. 2009, 21, 2217–2229. [Google Scholar] [CrossRef] [Green Version]
- Kadosh, R.C.; Lammertyn, J.; Izard, V. Are numbers special? An overview of chronometric, neuroimaging, developmental and comparative studies of magnitude representation. Prog. Neurobiol. 2008, 84, 132–147. [Google Scholar] [CrossRef] [Green Version]
- Leibovich, T.; Henik, A. Magnitude processing in non-symbolic stimuli. Front. Psychol. 2013, 4, 375. [Google Scholar] [CrossRef] [Green Version]
- Leibovich, T.; Katzin, N.; Harel, M.; Henik, A. From ‘sense of number’to ‘sense of magnitude’—The role of continuous magnitudes in numerical cognition. Behav. Brain Sci. 2016, 40, 1–62. [Google Scholar]
- Henik, A.; Gliksman, Y.; Kallai, A.; Leibovich, T. Size Perception and the Foundation of Numerical Processing. Curr. Dir. Psychol. Sci. 2017, 26, 45–51. [Google Scholar] [CrossRef]
- Henik, A.; Leibovich, T.; Naparstek, S.; Diesendruck, L.; Rubinsten, O. Quantities, amounts, and the numerical core system. Front. Hum. Neurosci. 2011, 5, 186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallistel, C.R.; Gelman, R. Non-verbal numerical cognition: From reals to integers. Trends Cogn. Sci. 2000, 4, 59–65. [Google Scholar] [CrossRef]
- Walsh, V. A theory of magnitude: Common cortical metrics of time, space and quantity. Trends Cogn. Sci. 2003, 7, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Piaget, J. The Child’s Conception of Number; Psychology Press: Oxford, UK, 1965. [Google Scholar]
- Shallice, T.; Evans, M.E. The involvement of the frontal lobes in cognitive estimation. Cortex 1978, 14, 294–303. [Google Scholar] [CrossRef]
- Aron, A.R.; Robbins, T.W.; Poldrack, R.A. Inhibition and the right inferior frontal cortex. Trends Cogn. Sci. 2004, 8, 170–177. [Google Scholar] [CrossRef]
- Duncan, J.; Owen, A.M. Common regions of the human frontal lobe recruited by diverse cognitive demands. Trends Neurosci. 2000, 23, 475–483. [Google Scholar] [CrossRef]
- Casey, B.J.; Forman, S.D.; Franzen, P.; Berkowitz, A.; Braver, T.S.; Nystrom, L.E.; Thomas, K.M.; Noll, D.C. Sensitivity of prefrontal cortex to changes in target probability: A functional MRI study. Hum. Brain Mapp. 2001, 13, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Bisbing, T.A.; Olm, C.A.; McMillan, C.T.; Rascovsky, K.; Baehr, L.; Ternes, K.; Irwin, D.J.; Clark, R.; Grossman, M. Estimating frontal and parietal involvement in cognitive estimation: A study of focal neurodegenerative diseases. Front. Hum. Neurosci. 2015, 9, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, S.J.; Burgess, J.W. Executive function. Curr. Biol. 2008, 18, R110–R114. [Google Scholar] [CrossRef] [Green Version]
- Mix, K.S.; Levine, S.C.; Newcombe, N.S. Development of quantitative thinking across correlated dimensions. In Continuous Issues in Numerical Cognition; Henik, A., Ed.; Elsevier: Amsterdam, Netherlands, 2016; pp. 1–33. [Google Scholar]
- Silverman, S.; Ashkenazi, S. Deconstructing the Cognitive Estimation Task: A Developmental Examination and Intra-Task Contrast. Sci. Rep. 2016, 6, 39316. [Google Scholar] [CrossRef] [Green Version]
- Ashkenazi, S.; Tsyganova, Y. The Cognitive Estimation Task Is Nonunitary: Evidence for Multiple Magnitude Representation Mechanisms Among Normative and ADHD College Students. J. Numer. Cogn. 2017, 2, 26. [Google Scholar] [CrossRef] [Green Version]
- Kriegeskorte, N.; Goebel, R.; Bandettini, P. Information-based functional brain mapping. Proc. Natl. Acad. Sci. USA 2006, 103, 3863–3868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R.; O’Carroll, R. Cognitive estimation in neurological disorders. Br. J. Clin. Psychol. 1995, 34, 223–228. [Google Scholar] [CrossRef]
- Dehaene, S. Varieties of numerical abilities. Cognition 1992, 44, 1–42. [Google Scholar] [CrossRef]
- Dehaene, S.; Piazza, M.; Pinel, P.; Cohen, L. Three parietal circuits for number processing. Cogn. Neuropsychol. 2003, 20, 487–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabner, R.H.; Ansari, D.; Reishofer, G.; Stern, E.; Ebner, F.; Neuper, C. Individual differences in mathematical competence predict parietal brain activation during mental calculation. NeuroImage 2007, 38, 346–356. [Google Scholar] [CrossRef]
- Ischebeck, A.; Zamarian, L.; Egger, K.; Schocke, M.; Delazer, M. Imaging early practice effects in arithmetic. NeuroImage 2007, 36, 993–1003. [Google Scholar] [CrossRef]
- Rickard, T.C.; Romero, S.G.; Basso, G.; Wharton, C.; Flitman, S.; Grafman, J. The calculating brain: An fMRI study. Neuropsychologia 2000, 38, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.S.; Chang, T.T.; Majid, A.; Caspers, S.; Eickhoff, S.B.; Menon, V. Functional heterogeneity of inferior parietal cortex during mathematical cognition assessed with cytoarchitectonic probability maps. Cereb. Cortex 2009, 19, 2930–2945. [Google Scholar] [CrossRef] [Green Version]
- Koelsch, S.; Schulze, K.; Sammler, D.; Fritz, T.; Muller, K.; Gruber, O. Functional architecture of verbal and tonal working memory: An FMRI study. Hum. Brain Mapp. 2009, 30, 859–873. [Google Scholar] [CrossRef] [Green Version]
- Newman, R.L.; Joanisse, M.F. Modulation of brain regions involved in word recognition by homophonous stimuli: An fMRI study. Brain Res. 2011, 1367, 250–264. [Google Scholar] [CrossRef]
- Simon, O.; Mangin, J.F.; Cohen, L.; Le Bihan, D.; Dehaene, S. Topographical layout of hand, eye, calculation, and language-related areas in the human parietal lobe. Neuron 2002, 33, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Todd, J.; Fougnie, D.; Marois, R. Visual Short-Term Memory Load Suppresses Temporo-Parietal Junction Activity and Induces Inattentional Blindness. Psychol. Sci. 2005, 16, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Kincade, J.M.; Abrams, R.A.; Astafiev, S.V.; Shulman, G.L.; Corbetta, M. An event-related functional magnetic resonance imaging study of voluntary and stimulus-driven orienting of attention. J. Neurosci. 2005, 25, 4593–4604. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.M.; Huettel, S.A. A Nexus Model of the Temporal-Parietal Junction. Trends Cogn. Sci. 2013, 17, 328–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabner, R.H.; Ansari, D.; Koschutnig, K.; Reishofer, G.; Ebner, F. The function of the left angular gyrus in mental arithmetic: Evidence from the associative confusion effect. Hum. Brain Mapp. 2013, 34, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.F.; Stern, E.R.; Gehring, W.J. Neural systems for error monitoring: Recent findings and theoretical perspectives. Neuroscientist 2007, 13, 160–172. [Google Scholar] [CrossRef]
- Devue, C.; Collette, F.; Balteau, E.; Degueldre, C.; Luxen, A.; Maquet, P.; Brédart, S. Here I am: The cortical correlates of visual self-recognition. Brain Res. 2007, 1143, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Peterson, B.S.; Skudlarski, P.; Gatenby, J.C.; Zhang, H.; Anderson, A.W.; Gore, J.C. An fMRI study of Stroop word-color interference: Evidence for cingulate subregions subserving multiple distributed attentional systems. Biol. Psychiatry 1999, 45, 1237–1258. [Google Scholar] [CrossRef]
- Koechlin, E.; Summerfield, C. An information theoretical approach to prefrontal executive function. Trends Cogn. Sci. 2007, 11, 229–235. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
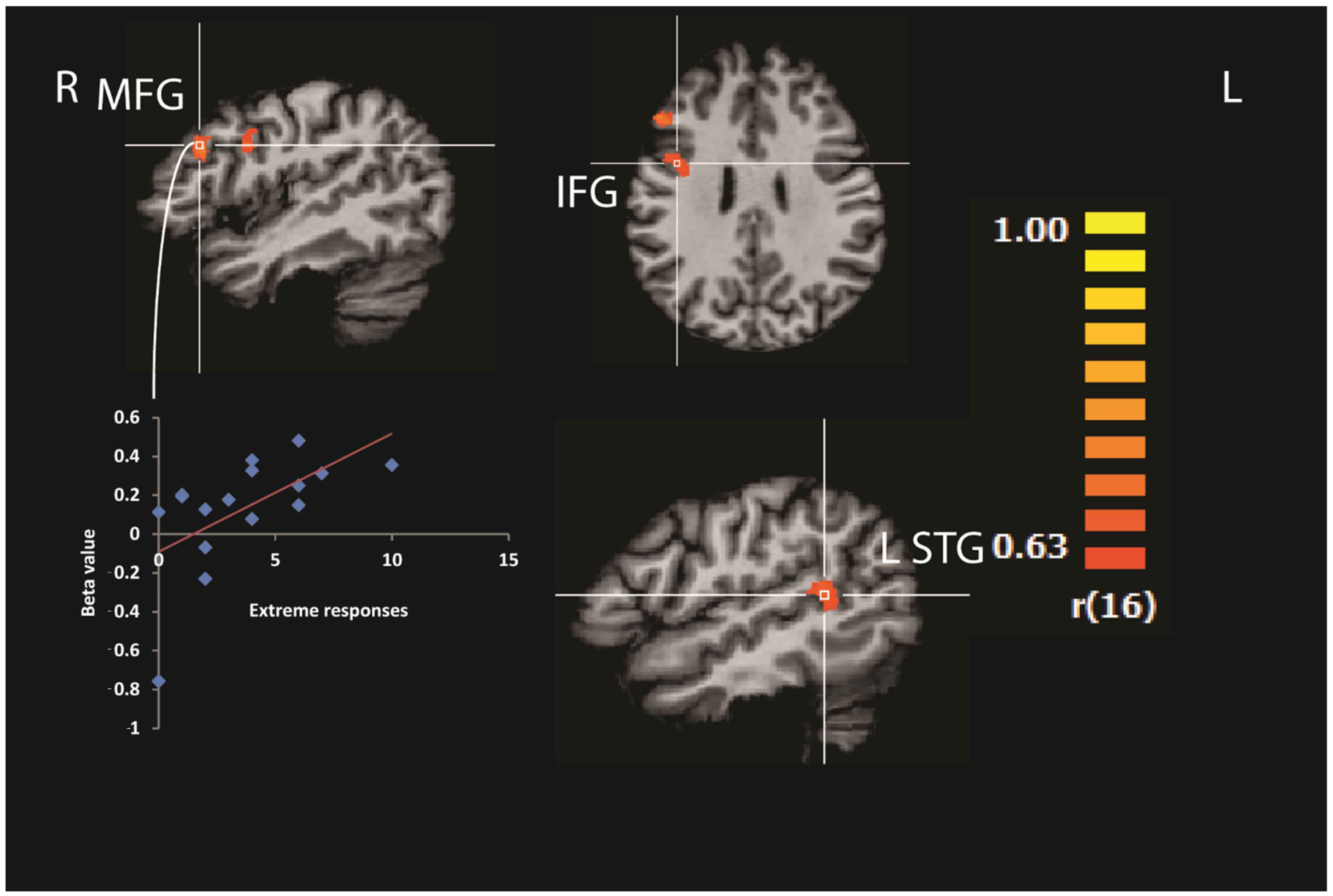{kind=link}
| Brain Region | Brodmann Area | Coordinates | t | Cluster Size (Voxels) | ||
|---|---|---|---|---|---|---|
| x | y | z | ||||
| CET > Exact Knowledge | ||||||
| R supramarginal gyrus | 40 | 59.02 | −47.32 | 22.22 | −4.2 | 1173 |
| R caudate extending to anterior cingulate and precentral gyrus | 6 | 9.33 | 11.54 | 27.47 | 7.8 | 39,576 |
| R precentral gyrus | 4 | 31.24 | −20.82 | 53.45 | 5.7 | 6717 |
| R lingual gyrus | 18 | 0.84 | −85.15 | −0.86 | 5.4 | 5930 |
| L R posterior cingulate | 30 | 14.01 | −52.1 | 6.64 | 5.2 | 1610 |
| L cerebellum | −26.22 | −53.53 | −20.98 | 5 | 12,457 | |
| R cerebellum | 32.32 | −56 | −28.23 | 5.7 | 6035 | |
| L lingual gyrus | 18 | −9.84 | −52.7 | 5.47 | 4.9 | 2210 |
| L insula extending to inferior frontal gyrus | 13 | −34.62 | 17.75 | 13.15 | 4.6 | 27,724 |
| L angular gyrus | 39 | −54 | −60.02 | 32.48 | 6.4 | 1065 |
| Numerical estimation compared to weight | ||||||
| No significant results | ||||||
| Numerical estimation compared to time | ||||||
| R lingual gyrus | 17 | 13.7 | −81.54 | −1.58 | 5.8 | 2273 |
| L lingual gyrus | 18 | −6.25 | −79.58 | −5.84 | 4.3 | 703 |
| L inferior frontal gyrus | 45 | −43.14 | −60.44 | 19.01 | −4.6 | 877 |
| Brain Region | Brodmann Area | Coordinates | t | Cluster Size (Voxels) | ||
|---|---|---|---|---|---|---|
| x | y | z | ||||
| Numerical estimation compared to weight | ||||||
| R postcentral gyrus | 43 | 57.44 | −12.12 | 14.76 | 81 | 423 |
| R cerebellum | 34.33 | −58.24 | −18.95 | 94 | 1353 | |
| R cerebellum | 35.39 | 16.59 | −5.45 | 81 | 554 | |
| R medial frontal gyrus | 9 | 1.6 | 37.27 | 26.41 | 94 | 1453 |
| L putamen | −23.73 | 2.89 | 18.6 | 100 | 805 | |
| L caudate | −18.85 | 2.7 | −0.98 | 88 | 684 | |
| Numerical estimation compared to time | ||||||
| R inferior frontal gyrus | 9 | 58.04 | 8.04 | 25.31 | 88 | 502 |
| R middle frontal gyrus | 8 | 45.6 | 26.61 | 41.04 | 88 | 403 |
| R middle frontal gyrus | 6 | 30.44 | 9.64 | 43.44 | 81 | 418 |
| Thalamus | 16.74 | −26.53 | −0.64 | 100 | 2179 | |
| R superior frontal gyrus | 6 | 12.95 | 11.84 | 64.61 | 100 | 1095 |
| R anterior cingulate | 33 | 3.72 | 18.84 | 20.89 | 100 | 3282 |
| R subcallosal gyrus | 25 | 7.79 | −12 | −12.42 | 88 | 564 |
| L superior frontal gyrus | 8 | −8.67 | 46.15 | 35.18 | 88 | 4011 |
| L lingual gyrus | 18 | −15.9 | −82.65 | 2.45 | 82 | 522 |
| L angular gyrus | 39 | −42.09 | −73.39 | 27.02 | 88 | 705 |
| L anterior cingulate | −35.73 | −2.04 | 28.19 | 94 | 494 | |
| L middle frontal gyrus | 8 | −48.4 | 13.72 | 41.36 | 100 | 1299 |
| L precentral gyrus | 4 | −54.19 | −15.9 | 37.31 | 88 | 644 |
| L middle frontal gyrus | 9 | −57.56 | 20.24 | 26.58 | 88 | 439 |
| Brain Region | Brodmann Area | Coordinates | r | Cluster Size (Voxels) | ||
|---|---|---|---|---|---|---|
| x | y | z | ||||
| R middle frontal gyrus | 46 | 55 | 27 | 29 | 0.78 | 546 |
| R inferior frontal gyrus | 9 | 46 | 6 | 27 | 0.74 | 757 |
| L cingulate gyrus | 32 | −26 | −12 | 35 | 0.84 | 1383 |
| L superior temporal gyrus | 22 | −56 | −48 | 11 | 0.81 | 1049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashkenazi, S.; Gliksman, Y.; Henik, A. Understanding Estimations of Magnitudes: An fMRI Investigation. Brain Sci. 2022, 12, 104. https://doi.org/10.3390/brainsci12010104
Ashkenazi S, Gliksman Y, Henik A. Understanding Estimations of Magnitudes: An fMRI Investigation. Brain Sciences. 2022; 12(1):104. https://doi.org/10.3390/brainsci12010104
Chicago/Turabian StyleAshkenazi, Sarit, Yarden Gliksman, and Avishai Henik. 2022. "Understanding Estimations of Magnitudes: An fMRI Investigation" Brain Sciences 12, no. 1: 104. https://doi.org/10.3390/brainsci12010104
APA StyleAshkenazi, S., Gliksman, Y., & Henik, A. (2022). Understanding Estimations of Magnitudes: An fMRI Investigation. Brain Sciences, 12(1), 104. https://doi.org/10.3390/brainsci12010104