
Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota



Abstract
:1. Introduction
2. Healthy Effects of Polyphenols and Antioxidants
3. Polyphenols and Gut Microbiota
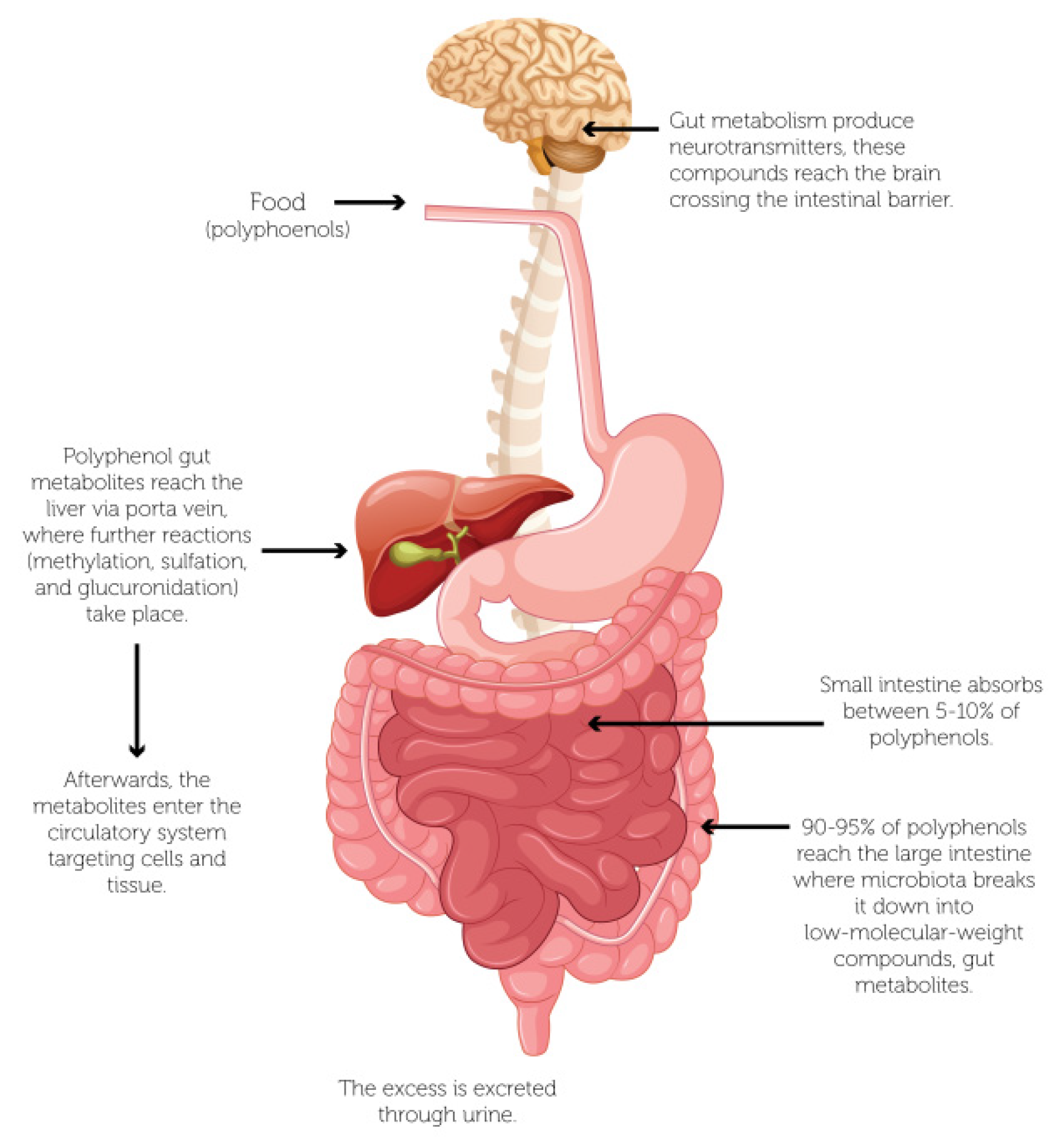
3.1. Biotransformation of Polyphenols by Gut Microbiota
3.2. Metabolism of Dietary Polyphenols: Role of the Microbiota
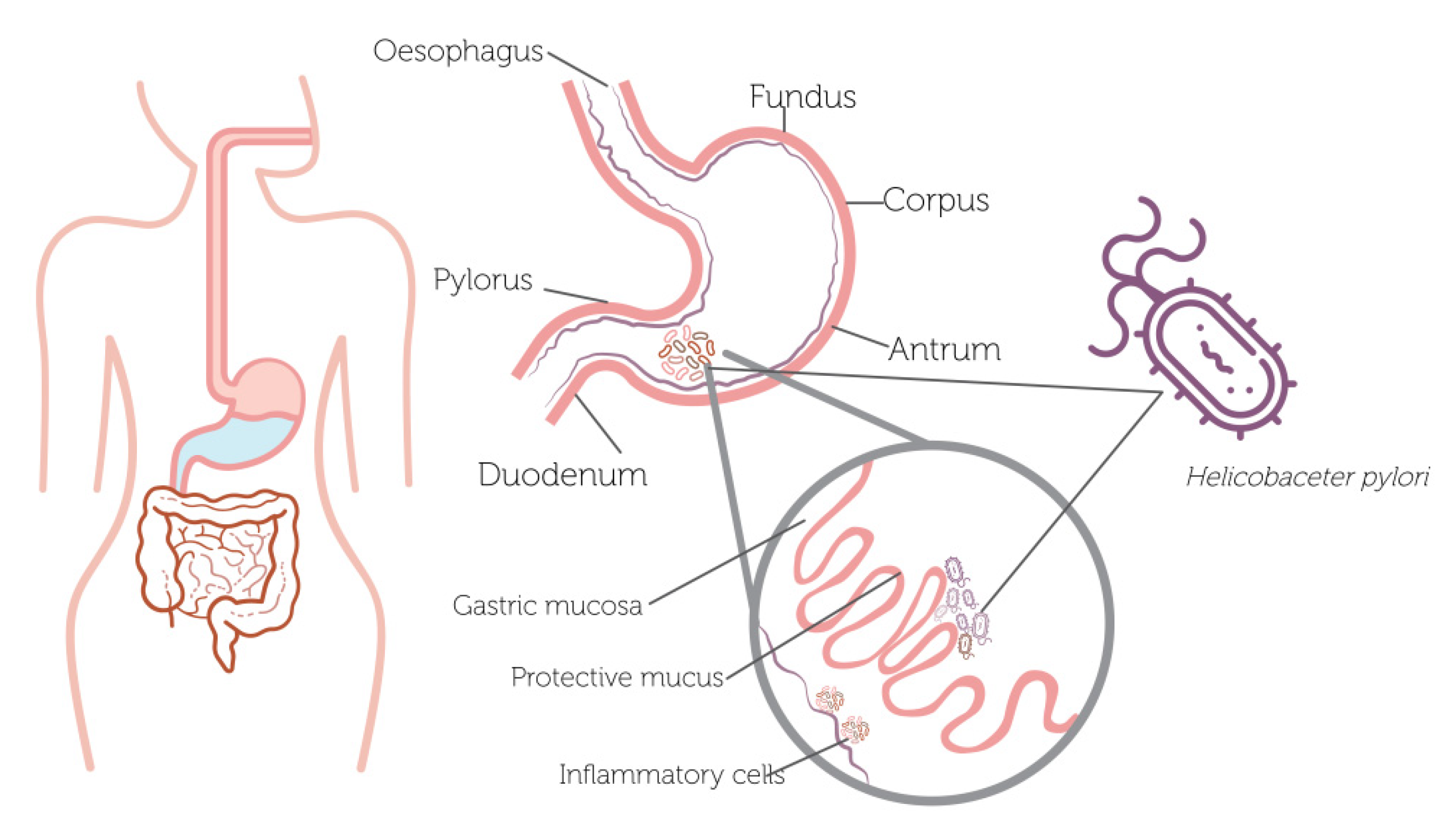
4. Anti-Helicobacter pylori Effects of Plant-Based Polyphenols
5. Impact of Polyphenols on the Composition of Gut Microbiota
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghobadi, E.; Ghanbarimasir, Z.; Emami, S. A review on the structures and biological activities of anti-Helicobacter pylori agents. Eur. J. Med. Chem. 2021, 223, 113669. [Google Scholar] [CrossRef]
- Baltas, N.; Karaoglu, S.A.; Tarakci, C.; Kolayli, S. Effect of propolis in gastric disorders: Inhibition studies on the growth of Helicobacter pylori and production of its urease. J. Enzyme Inhib. Med. Chem. 2016, 31, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Baranauskienė, L.; Matulis, D. Overview of human carbonic anhydrases. In Carbonic Anhydrase as Drug Target, 1st ed.; Matulis, D., Ed.; Springer: Cham, Switzerland, 2019; pp. 3–14. [Google Scholar]
- Howitt, M.R.; Lee, J.Y.; Lertsethtakarn, P.; Vogelmann, R.; Joubert, L.M.; Ottemann, K.M.; Amieva, M.R. ChePep controls Helicobacter pylori infection of the gastric glands and chemotaxis in the Epsilonproteobacteria. MBio 2011, 2, e00098-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolig, A.S.; Shanks, J.; Carter, J.E.; Ottemann, K.M. Helicobacter pylori requires TlpD-driven chemotaxis to proliferate in the antrum. Infect. Immun. 2012, 80, 3713–3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amieva, M.; Peek, R.M. Pathobiology of Helicobacter pylori–induced gastric cancer. Gastroenterology 2016, 150, 64–78. [Google Scholar] [CrossRef] [Green Version]
- Johnson, K.S.; Ottemann, K.M. Colonization, localization, and inflammation: The roles of H. pylori chemotaxis in vivo. Curr. Opin. Microbiol. 2018, 41, 51–57. [Google Scholar] [CrossRef]
- Keilberg, D.; Ottemann, K.M. How Helicobacter pylori senses, targets and interacts with the gastric epithelium. Environ. Microbiol. 2016, 18, 791–806. [Google Scholar] [CrossRef]
- McCracken, K.W.; Aihara, E.; Martin, B.; Crawford, C.M.; Broda, T.; Treguier, J.; Wells, J.M. Wnt/β-catenin promotes gastric fundus specification in mice and humans. Nature 2017, 541, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garza-González, E.; Perez-Perez, G.I.; Maldonado-Garza, H.J.; Bosques-Padilla, F.J. A review of Helicobacter pylori diagnosis, treatment, and methods to detect eradication. World J. Gastroenterol. 2014, 20, 1438–1449. [Google Scholar] [CrossRef]
- Suzuki, H.; Matsuzaki, J. Gastric cancer: Evidence boosts Helicobacter pylori eradication. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 458–460. [Google Scholar] [CrossRef]
- Sjomina, O.; Pavlova, J.; Niv, Y.; Leja, M. Epidemiology of Helicobacter pylori infection. Helicobacter 2018, 23, e12514. [Google Scholar] [CrossRef] [Green Version]
- Ghasemian, A.; Fattahi, A.; Shokouhi Mostafavi, S.K.; Almarzoqi, A.H.; Memariani, M.; Ben Braiek, O.; Mirforughi, S.A. Herbal medicine as an auspicious therapeutic approach for the eradication of Helicobacter pylori infection: A concise review. J. Cell. Physiol. 2019, 234, 16847–16860. [Google Scholar] [CrossRef]
- Parreira, P.; Soares, B.I.; Freire, C.S.; Silvestre, A.J.; Reis, C.A.; Martins, M.C.L.; Duarte, M.F. Eucalyptus spp. outer bark extracts inhibit Helicobacter pylori growth: In vitro studies. Ind. Crops Prod. 2017, 105, 207–214. [Google Scholar] [CrossRef]
- Singh, R.; Sripada, L.; Singh, R. Side effects of antibiotics during bacterial infection: Mitochondria, the main target in host cell. Mitochondrion 2014, 16, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.M.; Qian, W.; Qin, Y.Y.; He, J.; Zhou, Y.H. Probiotics in Helicobacter pylori eradication therapy: A systematic review and meta-analysis. World J. Gastroenterol. 2015, 21, 4345–4357. [Google Scholar] [CrossRef]
- Mandalari, G.; Bisignano, C.; Cirmi, S.; Navarra, M. Effectiveness of citrus fruits on Helicobacter pylori. Evid. Based Complement. Altern. Med. 2017, 2017, 379262. [Google Scholar] [CrossRef] [Green Version]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef]
- Liu, J.; He, Z.; Ma, N.; Chen, Z.Y. Beneficial effects of dietary polyphenols on high-fat diet-induced obesity linking with modulation of gut microbiota. J. Agric. Food Chem. 2019, 68, 33–47. [Google Scholar] [CrossRef]
- Tsimogiannis, D.; Oreopoulou, V. Classification of phenolic compounds in plants. In Polyphenols in Plants: Isolation, Purification and Extract Preparation, 2nd ed.; Watson, R.R., Ed.; Academic Press: New York, NY, USA, 2019; pp. 263–281. [Google Scholar]
- Aravind, S.M.; Wichienchot, S.; Tsao, R.; Ramakrishnan, S.; Chakkaravarthi, S. Role of dietary polyphenols on gut microbiota, their metabolites and health benefits. Food Res. Int. 2021, 142, 110189. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.; Rocha, S.; de Freitas, V. Going “Green” in the prevention and management of atherothrombotic diseases: The role of dietary polyphenols. J. Clin. Med. 2021, 10, 1490. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, G.; Sureda, A.; Orhan, I.E.; Battino, M.; Arnone, A.; Jiménez-García, M.; Capó, X.; Cabot, J.; Sanadgol, N.; Giampieri, F.; et al. The neuroprotective effects of polyphenols, their role in innate immunity and the interplay with the microbiota. Neurosci. Biobehav. Rev. 2021, 128, 437–453. [Google Scholar] [CrossRef]
- Prabhu, S.; Molath, A.; Choksi, H.; Kumar, S.; Mehra, R. Classifications of polyphenols and their potential application in human health and diseases. Int. J. Physiol. Nutr. Phys. Educ. 2021, 6, 293–301. [Google Scholar] [CrossRef]
- Zhang, L.; Han, Z.; Granato, D. Polyphenols in foods: Classification, methods of identification, and nutritional aspects in human health. In Advances in Food and Nutrition Research Series 98: Application of Polyphenols in Foods and Food Models, 1st ed.; Granato, D., Ed.; Academic Press: New York, NY, USA, 2021; pp. 1–33. [Google Scholar]
- Kaur, K.; Asthir, B.; Verma, D.K. Biosynthesis, bioavailability, and metabolism of plant polyphenols: Biological activities and their potential benefits in human health. In Phytochemicals in Food and Health: Perspectives for Research and Technological Development, 1st ed.; Verma, D.K., Thakur, M., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 233–258. [Google Scholar]
- Huang, R.; Xu, C. An overview of the perception and mitigation of astringency associated with phenolic compounds. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1036–1074. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Yang, H.; Capanoglu, E.; Cao, H.; Xiao, J. Technological aspects and stability of polyphenols. In Polyphenols: Properties, Recovery, and Applications, 1st ed.; Galanakis, C.M., Ed.; Woodhead Publishing: Sawston, UK, 2018; pp. 295–323. [Google Scholar]
- Guerra-Valle, M.; Lillo-Pérez, S.; Petzold, G.; Orellana-Palma, P. Effect of freeze crystallization on quality properties of two endemic Patagonian berries juices: Murta (Ugni molinae) and arrayan (Luma apiculata). Foods 2021, 10, 466. [Google Scholar] [CrossRef] [PubMed]
- Orellana-Palma, P.; Tobar-Bolaños, G.; Casas-Forero, N.; Zúñiga, R.N.; Petzold, G. Quality attributes of cryoconcentrated calafate (Berberis microphylla) juice during refrigerated storage. Foods 2020, 9, 1314. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenol-mediated gut microbiota modulation: Toward prebiotics and further. Front. Nutr. 2021, 8, 689456. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, M.; Sundralingam, U.; Palanisamy, U.D. Polyphenols as prebiotics in the management of high-fat diet-induced obesity: A systematic review of animal studies. Foods 2021, 10, 299. [Google Scholar] [CrossRef]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the future of probiotics and prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef]
- Joseph, S.V.; Edirisinghe, I.; Burton-Freeman, B.M. Fruit polyphenols: A review of anti-inflammatory effects in humans. Crit. Rev. Food Sci. Nutr. 2016, 56, 419–444. [Google Scholar] [CrossRef]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef]
- Shaw, P.; Chattopadhyay, A. Nrf2–ARE signaling in cellular protection: Mechanism of action and the regulatory mechanisms. J. Cell. Physiol. 2020, 235, 3119–3130. [Google Scholar] [CrossRef]
- Skrovankova, S.; Sumczynski, D.; Mlcek, J.; Jurikova, T.; Sochor, J. Bioactive compounds and antioxidant activity in different types of berries. Int. J. Mol. Sci. 2015, 16, 24673–24706. [Google Scholar] [CrossRef] [Green Version]
- Trujillo-Mayol, I.; Guerra-Valle, M.; Casas-Forero, N.; Sobral, M.M.C.; Viegas, O.; Alarcón-Enos, J.; Pinho, O. Western dietary pattern antioxidant intakes and oxidative stress: Importance during the SARS-CoV-2/COVID-19 Pandemic. Adv. Nutr. 2021, 12, 670–681. [Google Scholar] [CrossRef]
- Liu, Q.; Meng, X.; Li, Y.; Zhao, C.N.; Tang, G.Y.; Li, S.; Li, H.B. Natural products for the prevention and management of Helicobacter pylori infection. Compr. Rev. Food Sci. Food Saf. 2018, 17, 937–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.E.; Khoi, P.N.; Xia, Y.; Park, J.S.; Joo, Y.E.; Kim, K.K.; Choi, S.Y.; Jung, Y.D. Helicobacter pylori and interleukin-8 in gastric cancer. World J. Gastroenterol. 2013, 45, 8192–8202. [Google Scholar] [CrossRef] [PubMed]
- Keates, S.; Hitti, Y.S.; Upton, M.; Kelly, C.P. Helicobacter pylori infection activates NF-kappa B in gastric epithelial cells. Gastroenterology 1997, 113, 1099–1109. [Google Scholar] [CrossRef]
- Seo, J.H.; Lim, J.W.; Kim, H.; Kim, K.H. Helicobacter pylori in a Korean isolate activates mitogen-activated protein kinases, AP-1, and NF-kB and induces chemokine expression in gastric epithelial AGS cells. Lab. Investig. 2003, 84, 49–62. [Google Scholar] [CrossRef]
- Song, M.Y.; Lee, D.Y.; Kim, E.H. Anti-inflammatory and anti-oxidative effect of Korean propolis on Helicobacter pylori-induced gastric damage in vitro. J. Microbiol. 2020, 58, 878–885. [Google Scholar] [CrossRef]
- Tominaga, K.; Kawahara, T.; Sano, T.; Toida, K.; Kuwano, Y.; Sasaki, H.; Kawai, T.; Teshima-Kondo, S.; Rokutan, K. Evidence for cancer-associated expression of NADPH oxidase 1 (Nox1)-based oxidase system in the human stomach. Free Radic. Biol. Med. 2007, 43, 1627–1638. [Google Scholar] [CrossRef]
- Khan, H.; Sureda, A.; Belwal, T.; Çetinkaya, S.; Süntar, İ.; Tejada, S.; Ullah, H.; Devkota, H.P.; Aschner, M.; Aschner, M. Polyphenols in the treatment of autoimmune diseases. Autoimmun. Rev. 2019, 18, 647–657. [Google Scholar] [CrossRef]
- Khan, H.; Ullah, H.; Castilho, P.C.M.F.; Gomila, A.S.; D’Onofrio, G.; Filosa, R.; Wang, F.; Nabavi, S.M.; Daglia, M.; Sanches-Silva, A.; et al. Targeting NF-κB signaling pathway in cancer by dietary polyphenols. Crit. Rev. Food Sci. Nutr. 2020, 60, 2790–2800. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Rolt, A.; Cox, L.S. Structural basis of the anti-ageing effects of polyphenolics: Mitigation of oxidative stress. BMC Chem. 2020, 14, 1–13. [Google Scholar] [CrossRef]
- Vezza, T.; Rodríguez-Nogales, A.; Algieri, F.; Utrilla, M.P.; Rodriguez-Cabezas, M.E.; Galvez, J. Flavonoids in inflammatory bowel disease: A review. Nutrients 2016, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efenberger-Szmechtyk, M.; Nowak, A.; Czyzowska, A. Plant extracts rich in polyphenols: Antibacterial agents and natural preservatives for meat and meat products. Crit. Rev. Food Sci. Nutr. 2021, 61, 149–178. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Chen, Y. Polyphenol supplementation benefits human health via gut microbiota: A systematic review via meta-analysis. J. Funct. Foods 2020, 66, 103829. [Google Scholar] [CrossRef]
- Burkholder-Cooley, N.; Rajaram, S.; Haddad, E.; Fraser, G.E.; Jaceldo-Siegl, K. Comparison of polyphenol intakes according to distinct dietary patterns and food sources in the Adventist Health Study-2 cohort. Br. J. Nutr. 2016, 115, 2162–2169. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Goñi, I. Antioxidant capacity of the Spanish Mediterranean diet. Food Chem. 2006, 94, 442–447. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Knaze, V.; Rothwell, J.A.; Hémon, B.; Moskal, A.; Overvad, K.; Touillaud, M. Dietary polyphenol intake in Europe: The European Prospective Investigation into Cancer and Nutrition (EPIC) study. Eur. J. Nutr. 2016, 55, 1359–1375. [Google Scholar] [CrossRef] [PubMed]
- Huang, D. Dietary antioxidants and health promotion. Antioxidants 2018, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Tsugawa, H.; Suzuki, H. Oxidative stress in stomach cancer. In Cancer: Oxidative Stress and Dietary Antioxidants, 2nd ed.; Preedy, V.R., Patel, V.B., Eds.; Academic Press: New York, NY, USA, 2021; pp. 49–54. [Google Scholar]
- Capparelli, R.; Iannelli, D. Genetics of host protection against Helicobacter pylori infections. Int. J. Mol. Sci. 2021, 22, 3192. [Google Scholar] [CrossRef]
- Jain, U.; Saxena, K.; Chauhan, N. Helicobacter pylori induced reactive oxygen species: A new and developing platform for detection. Helicobacter 2021, 26, e12796. [Google Scholar] [CrossRef] [PubMed]
- Öztekin, M.; Yilmaz, B.; Ağagündüz, D.; Capasso, R. Overview of Helicobacter pylori Infection: Clinical Features, treatment, and nutritional aspects. Diseases 2021, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Judaki, A.; Rahmani, A.; Feizi, J.; Asadollahi, K.; Hafezi Ahmadi, M.R. Curcumin in combination with triple therapy regimes ameliorates oxidative stress and histopathologic changes in chronic gastritis-associated Helicobacter pylori infection. Arq. Gastroenterol. 2017, 54, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, N.L.; Shabib, S.; Sherman, P.M. Capsaicin as an inhibitor of the growth of the gastric pathogen Helicobacter pylori. FEMS Microbiol. Lett. 1997, 146, 223–227. [Google Scholar] [CrossRef]
- Lee, I.O.; Lee, K.H.; Pyo, J.H.; Kim, J.H.; Choi, Y.J.; Lee, Y.C. Anti-inflammatory effect of capsaicin in Helicobacter pylori-infected gastric epithelial cells. Helicobacter 2007, 12, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, Y.; Yamamoto, Y.; Hara, Y.; Shimamura, T. A combination effect of epigallocatechin gallate, a major compound of green tea catechins, with antibiotics on Helicobacter pylori growth in vitro. Curr. Microbiol. 2003, 47, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.F.; Pogačnik, L. Polyphenols from food and natural products: Neuroprotection and safety. Antioxidants 2020, 9, 61. [Google Scholar] [CrossRef] [Green Version]
- Reis, A.; Perez-Gregorio, R.; Mateus, N.; de Freitas, V. Interactions of dietary polyphenols with epithelial lipids: Advances from membrane and cell models in the study of polyphenol absorption, transport and delivery to the epithelium. Crit. Rev. Food Sci. Nutr. 2021, 61, 3007–3030. [Google Scholar] [CrossRef]
- Makarewicz, M.; Drożdż, I.; Tarko, T.; Duda-Chodak, A. The Interactions between polyphenols and microorganisms, especially gut microbiota. Antioxidants 2021, 10, 188. [Google Scholar] [CrossRef]
- Martínez, I.; Wallace, G.; Zhang, C.; Legge, R.; Benson, A.K.; Carr, T.P.; Walter, J. Diet-induced metabolic improvements in a hamster model of hypercholesterolemia are strongly linked to alterations of the gut microbiota. Appl. Environ. Microbiol. 2009, 75, 4175–4184. [Google Scholar] [CrossRef] [Green Version]
- Hervert-Hernández, D.; Goñi, I. Dietary polyphenols and human gut microbiota: A review. Food Rev. Int. 2011, 27, 154–169. [Google Scholar] [CrossRef]
- Filosa, S.; di Meo, F.; Crispi, S. Polyphenols-gut microbiota interplay and brain neuromodulation. Neural Regen. Res. 2018, 13, 154–169. [Google Scholar] [CrossRef]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowd, V.; Karim, N.; Shishir, M.R.I.; Xie, L.; Chen, W. Dietary polyphenols to combat the metabolic diseases via altering gut microbiota. Trends Food Sci. Technol. 2019, 93, 81–93. [Google Scholar] [CrossRef]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Bartolomé, B. A survey of modulation of gut microbiota by dietary polyphenols. BioMed Res. Int. 2015, 2015, 850902. [Google Scholar] [CrossRef]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef]
- Kumar-Singh, A.; Cabral, C.; Kumar, R.; Ganguly, R.; Kumar-Rana, H.; Gupta, A.; Pandey, A.K. Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef] [Green Version]
- Williamson, G.; Clifford, M.N. Role of the small intestine, colon and microbiota in determining the metabolic fate of polyphenols. Biochem. Pharmacol. 2017, 139, 24–39. [Google Scholar] [CrossRef] [Green Version]
- Dey, P. Gut microbiota in phytopharmacology: A comprehensive overview of concepts, reciprocal interactions, biotransformations and mode of actions. Pharmacol. Res. 2019, 147, 104367. [Google Scholar] [CrossRef] [PubMed]
- Danneskiold-Samsøe, N.B.; Barros, H.D.D.F.Q.; Santos, R.; Bicas, J.L.; Cazarin, C.B.B.; Madsen, L.; Junior, M.R.M. Interplay between food and gut microbiota in health and disease. Food Res. Int. 2019, 115, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Chen, Y.; Cheng, M.; Zhang, X.; Zheng, X.; Zhang, Z. The modulatory effect of polyphenols from green tea, oolong tea and black tea on human intestinal microbiota in vitro. J. Food Sci. Technol. 2018, 55, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Alves-Santos, A.M.; Sugizaki, C.S.A.; Lima, G.C.; Naves, M.M.V. Prebiotic effect of dietary polyphenols: A systematic review. J. Funct. Foods 2020, 74, 104169. [Google Scholar] [CrossRef]
- Milutinović, M.; Dimitrijević-Branković, S.; Rajilić-Stojanović, M. Plant extracts rich in polyphenols as potent modulators in the growth of probiotic and pathogenic intestinal microorganisms. Front. Nutr. 2021, 8, 688843. [Google Scholar] [CrossRef]
- Chiu, H.F.; Venkatakrishnan, K.; Golovinskaia, O.; Wang, C.K. Gastroprotective Effects of polyphenols against various gastro-intestinal disorders: A mini-review with special focus on clinical evidence. Molecules 2021, 26, 2090. [Google Scholar] [CrossRef]
- Mandici, A.; Covrig, A.A.; Macovei, I. Food phytochemicals against Helicobacter pylori. Rom. J. Pharm. Pract. 2021, 14, 11–17. [Google Scholar] [CrossRef]
- Chung, J.G. Inhibitory actions of ellagic acid on growth and arylamine N-acetyltransferase activity in strains of Helicobacter pylori from peptic ulcer patients. Microbios 1998, 93, 115–127. [Google Scholar]
- Martini, S.; d’Addario, C.; Colacevich, A.; Focardi, S.; Borghini, F.; Santucci, A.; Figura, N.; Rossi, C. Antimicrobial activity against Helicobacter pylori strains and antioxidant properties of blackberry leaves (Rubus ulmifolius) and isolated compounds. Int. J. Antimicrob. Agents 2009, 34, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Jiang, A.; Qi, B.; Ma, Z.; Xiong, Y.; Dou, J.; Wang, J. Resveratrol protects against Helicobacter pylori-associated gastritis by combating oxidative stress. Int. J. Mol. Sci. 2015, 16, 27757–27769. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; Yeo, M.; Choue, J.S.; Jin, J.H.; Park, S.J.; Cheong, J.Y.; Lee, K.J.; Kim, J.H.; Hahm, K.B. Protective mechanism of epigallocatechin-3-gallate against Helicobacter pylori-induced gastric epithelial cytotoxicity via the blockage of TLR-4 signaling. Helicobacter 2004, 9, 632–642. [Google Scholar] [CrossRef]
- Takeuchi, H.; Trang, V.T.; Morimoto, N.; Nishida, Y.; Matsumura, Y.; Sugiura, T. Natural products and food components with anti-Helicobacter pylori activities. World J. Gastroenterol. 2014, 20, 8971–8978. [Google Scholar] [CrossRef] [PubMed]
- Escandón, R.A.; del Campo, M.; López-Solis, R.; Obreque-Slier, E.; Toledo, H. Antibacterial effect of kaempferol and (-)-epicatechin on Helicobacter pylori. Eur. Food Res. Technol. 2016, 242, 1495–1502. [Google Scholar] [CrossRef]
- Chua, C.S.; Yang, K.C.; Chen, J.H.; Liu, Y.H.; Hsu, Y.H.; Lee, H.C.; Huang, S.Y. The efficacy of blueberry and grape seed extract combination on triple therapy for Helicobacter pylori eradication: A randomized controlled trial. Int. J. Food Sci. Nutr. 2016, 67, 177–183. [Google Scholar] [CrossRef]
- Cardoso, O.; Donato, M.M.; Luxo, C.; Almeida, N.; Liberal, J.; Figueirinha, A.; Batista, M.T. Anti-Helicobacter pylori potential of Agrimonia eupatoria L. and Fragaria vesca. J. Funct. Foods 2018, 44, 299–303. [Google Scholar] [CrossRef]
- Park, J.U.; Cho, J.S.; Kim, J.S.; Kim, H.K.; Jo, Y.H.; Rahman, M.A.A.; Lee, Y.I. Synergistic effect of Rubus crataegifolius and Ulmus macrocarpa against Helicobacter pylori clinical isolates and gastritis. Front. Pharmacol. 2020, 11, 4. [Google Scholar] [CrossRef]
- Torres, E.; García, A.; Aranda, M.; Saéz, V.; Zúñiga, F.; Alarcón, J.; Pastene, E. One-step purification of two semi-synthetic epicatechin adducts prepared from avocado peels procyanidins by centrifugal partition chromatography and evaluation of their anti-inflammatory effects on adenocarcinoma gastric cells infected with Helicobacter pylori. J. Chil. Chem. Soc. 2018, 63, 4222–4228. [Google Scholar] [CrossRef] [Green Version]
- Spósito, L.; Oda, F.B.; Vieira, J.H.; Carvalho, F.A.; dos Santos Ramos, M.A.; de Castro, R.C.; Chorilli, M. In vitro and in vivo anti-Helicobacter pylori activity of Casearia sylvestris leaf derivatives. J. Ethnopharmacol. 2019, 233, 1–12. [Google Scholar] [CrossRef]
- Betoret, E.; Betoret, N.; Arilla, A.; Bennár, M.; Barrera, C.; Codoñer, P.; Fito, P. No invasive methodology to produce a probiotic low humid apple snack with potential effect against Helicobacter pylori. J. Food Eng. 2012, 110, 289–293. [Google Scholar] [CrossRef]
- Barrera, C.; Burca, C.; Betoret, E.; García-Hernández, J.; Hernández, M.; Betoret, N. Improving antioxidant properties and probiotic effect of clementine juice inoculated with Lactobacillus salivarius spp. salivarius (CECT 4063) by trehalose addition and/or sublethal homogenisation. Int. J. Food Sci. Technol. 2019, 54, 2109–2122. [Google Scholar] [CrossRef]
- Gao, T.; Hou, M.; Zhang, B.; Pan, X.; Liu, C.; Sun, C.; Jia, M.; Lin, S.; Xiong, K.; Ma, A. Effects of cranberry beverage on oxidative stress and gut microbiota in subjects with Helicobacter pylori infection: A randomized, double-blind, placebo-controlled trial. Food Funct. 2021, 12, 6878–6888. [Google Scholar] [CrossRef]
- Jahan, S.; Mahmud, M.H.; Khan, Z.; Alam, A.; Khalil, A.A.; Rauf, A.; Tareq, A.M.; Nainu, F.; Tareq, S.M.; Emran, B.T.; et al. Health promoting benefits of pongamol: An overview. Biomed. Pharmacother. 2021, 142, 112109. [Google Scholar] [CrossRef] [PubMed]
- Mishra, L.K.; Sarkar, D.; Mentreddy, R.; Shetty, K. Evaluation of phenolic bioactive-linked anti-hyperglycemic and Helicobacter pylori inhibitory activities of Asian Basil (Ocimum spp.) varieties. J. Herb. Med. 2020, 20, 100310. [Google Scholar] [CrossRef]
- Amaral, O.; Fernandes, I.; Veiga, N.; Pereira, C.; Chaves, C.; Nelas, P.; Silva, D. Living conditions and Helicobacter pylori in adults. Biomed Res. Int. 2017, 2017, 9082716. [Google Scholar] [CrossRef] [Green Version]
- Mard, S.A.; Haghighian, H.K.; Sebghatulahi, V.; Ahmadi, B. Dietary factors in relation to Helicobacter pylori infection. Gastroenterol. Res. Pract. 2014, 2014, 826910. [Google Scholar] [CrossRef] [Green Version]
- Ashaolu, T.J. Immune boosting functional foods and their mechanisms: A critical evaluation of probiotics and prebiotics. Biomed. Pharmacother. 2020, 130, 110625. [Google Scholar] [CrossRef]
- Fatima, N.; Akhtar, T.; Sheikh, N. Prebiotics: A novel approach to treat hepatocellular carcinoma. Can. J. Gastroenterol. Hepatol. 2017, 2017, 6238106. [Google Scholar] [CrossRef]
- Pathak, S.; Kesavan, P.; Banerjee, A.; Banerjee, A.; Celep, G.S.; Bissi, L.; Marotta, F. Metabolism of dietary polyphenols by human gut microbiota and their health benefits. In Polyphenols: Mechanisms of Action in Human Health and Disease, 2nd ed.; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: New York, NY, USA, 2018; pp. 347–359. [Google Scholar]
- Kemperman, R.A.; Bolca, S.; Roger, L.C.; Vaughan, E.E. Novel approaches for analysing gut microbes and dietary polyphenols: Challenges and opportunities. Microbiology 2010, 156, 3224–3231. [Google Scholar] [CrossRef] [Green Version]
- Morais, C.A.; de Rosso, V.V.; Estadella, D.; Pisani, L.P. Anthocyanins as inflammatory modulators and the role of the gut microbiota. J. Nutr. Biochem. 2016, 33, 1–7. [Google Scholar] [CrossRef]
- Vamanu, E.; Gatea, F. Correlations between microbiota bioactivity and bioavailability of functional compounds: A mini-review. Biomedicines 2020, 8, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The two way polyphenols-microbiota interactions and their effects on obesity and related metabolic diseases. Front. Nutr. 2019, 20, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashwin, K.; Pattanaik, A.K.; Howarth, G.S. Polyphenolic bioactives as an emerging group of nutraceuticals for promotion of gut health: A review. Food Biosci. 2021, 44, 101376. [Google Scholar] [CrossRef]
- Havlik, J.; Marinello, V.; Gardyne, A.; Hou, M.; Mullen, W.; Morrison, D.J.; Preston, T.; Combet, E.; Edwards, C.A. Dietary fibres differentially impact on the production of phenolic acids from rutin in an in vitro fermentation model of the human gut microbiota. Nutrients 2020, 12, 1577. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Polyphenol Group | Compound | Gut Microbiome | Metabolites |
|---|---|---|---|
| Anthocyanins | Cyanidin | Bifidobacterium lactis BB-12 | 3,4-Dihydroxybenzoic Acid |
| Malvidin | Lactobacillus acidophilus LA-5 | 3,4-Dimethocybenzoic Acid | |
| Ponidin Pelargonidin | Lactobacillus casei Lactobacillus plantarum | 3-Methoxy-4-Hydroxybenzoic Acid 4-Hydroxybenzoic Acid | |
| Flavan-3-ols | Catechin | Clostridium cocoides | 3-(3-Hydroxyphenyl)-Propionic Acid |
| Epicatechin Epigallocatechin | Bifidobacterium spp. | 5-(3′,4′-Dihydroxyphenil)-Γ-Valerolactone 5-(3,4-Dihydroxyphenyl)-Valeric Acid 3-(3,4-Dihydroxyphenyl)-Propionic Acid | |
| 5-(3′,4′-Dihydroxyphenyl)-Γ-Valerolactone 5-(3′,5′-Dihydroxyphenyl)-Γ-Valerolactone | |||
| Flavanones | Naringenin | Clostridum Strains | 3-(4-hydroxyphenyl)-propionic acid |
| Eubacterium ramulus | |||
| Flavones | Luteolin | Clostridium orbiscindens | 3-(3,4-Dihydroxyphenyl)-Propionic Acid |
| Apigenin | Enterococcus avium | 3-(4-Dihydroxyphenyl)-Propionic Acid 3-(3-Dihydroxyphenyl)-Propionic Acid 4-Hydroxycinnamic Acid | |
| Flavonols | Kaempferol | Clostridium orbiscindens | 2-(4-Hydroxyphenyl)-Propionic Acid |
| Quercetin Myricetin | Clostridium orbiscindens Eubacterium oxidoreducens Eubacterium ramulus Enterococcus casseliflavus Clostridium orbiscindens Eubacterium oxidoreducens | 2-(3,4-Dihydroxyphenyl)-Acetic Acid 2-(3-Hydroxyphenyl)-Acetic Acid 3-(3,4-Dihydroxyphenyl)-Propionic Acid 3-(3-Hydroxyphenyl)-Acetic Acid 2-(3,5-Dihydrosyphenyl)-Acetic Acid 2-(3-Hydroxyphenyl)-Acetic Acid | |
| Isoflavones | Daidzein | Bacteroides ovatus | (S)-Equol |
| Streptococcus intermedius Rumnococcus products Eggerthella sp. Julong 732 Enterococcus faecium EPI1 Lactobacillus mucosae EPI2 Finegoldia magna EPI3 Clostridium sp. HGHA136 | O-Demethylangolesin |
| Sample | Test Type | Anti-Helicobacter pylori Effect | Reference |
|---|---|---|---|
| Bergamot Juice | In vitro | Inhibited the Growth and Reduced the Viability | [17] |
| Citrus Fruit Extract | In vitro | Antimicrobial Activity, Affecting the Urease Activity | [17] |
| 6-Shogaol, 6-Gingerol, 8-Gingerol 10-Gingerol Curcumin Propolis Muscadine Grapes | In vitro | Antibacterial Activity Inhibited the Growth and Damaged Its Cytoplasmic Membrane | [89] |
| Kaempferol And (-)-Epicatechin | Inhibition Halo Test | Suppressed the Growth, also proved the Inhibitory Effect depends on the Pathogen Exposure Time to the Polyphenol | [90] |
| Blueberry and Grape Seed Extract | Standard Triple Therapy Plus | There was no Significant Difference between this Therapy and one Standard Triple Therapy Plus Placebo | [91] |
| Aqueous Extract from Fragaria Vesca Leaves | Antibacterial Activity Evaluation | Revealed an Intense Activity Due to the Source of Ellagitannins | [92] |
| Red Raspberry Elm Tree | In vitro | Inhibited (Alone or in Combination) the Growth of H. Pylori | [93] |
| Avocado | Anti-Inflamatory Activity | Lowered the Early Inflammatory Signals | [94] |
| Caseria Sylvestris Leaves | In vitro and in vivo | Inhibited the Growth of H. Pylori | [95] |
| Low Humidity Apple Snack Impregnated with Mandarin and Pineapple/Grape Juice, and Inoculated with Lactobacillus Salivarius spp. Salivarius | In vivo | No Evidence Suggested Whether Bioactive Compounds Present in the Juices Affected the Eradication Rate | [96] |
| Clementine Juice Inoculated with Lactobacillus Salivarius spp. Salivarius | In vitro | Inhibited the Growth of H. Pylori | [97] |
| Cranberry Juice | Standard Triple Therapy Plus | Inhibited the Growth of H. Pylori | [98] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra-Valle, M.; Orellana-Palma, P.; Petzold, G. Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota. Antioxidants 2022, 11, 109. https://doi.org/10.3390/antiox11010109
Guerra-Valle M, Orellana-Palma P, Petzold G. Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota. Antioxidants. 2022; 11(1):109. https://doi.org/10.3390/antiox11010109
Chicago/Turabian StyleGuerra-Valle, María, Patricio Orellana-Palma, and Guillermo Petzold. 2022. "Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota" Antioxidants 11, no. 1: 109. https://doi.org/10.3390/antiox11010109
APA StyleGuerra-Valle, M., Orellana-Palma, P., & Petzold, G. (2022). Plant-Based Polyphenols: Anti-Helicobacter pylori Effect and Improvement of Gut Microbiota. Antioxidants, 11(1), 109. https://doi.org/10.3390/antiox11010109