



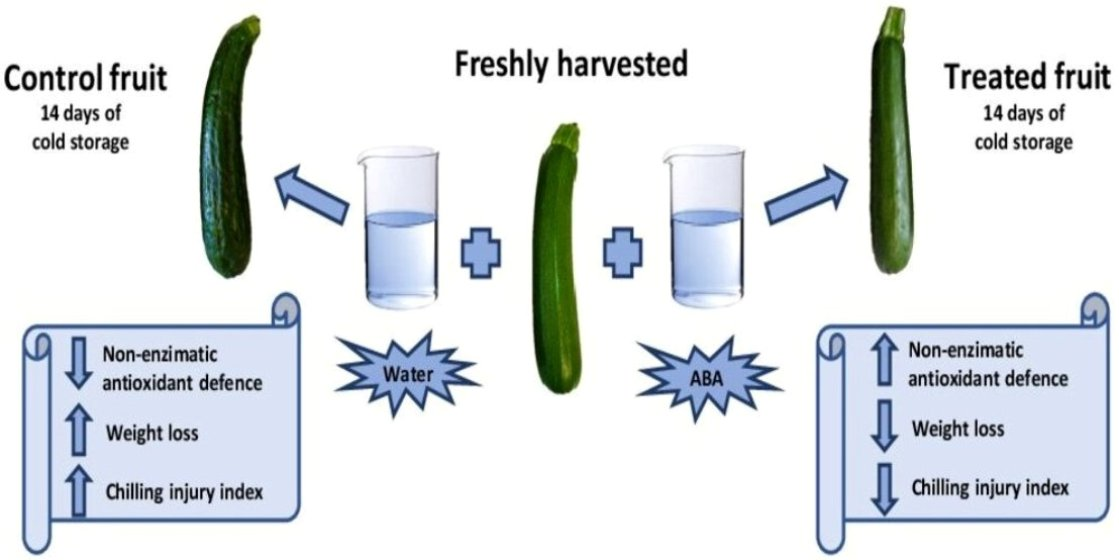
Postharvest Treatment with Abscisic Acid Alleviates Chilling Injury in Zucchini Fruit by Regulating Phenolic Metabolism and Non-Enzymatic Antioxidant System
 ,
, 
 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sample Material, Treatments and Cold Storage Conditions
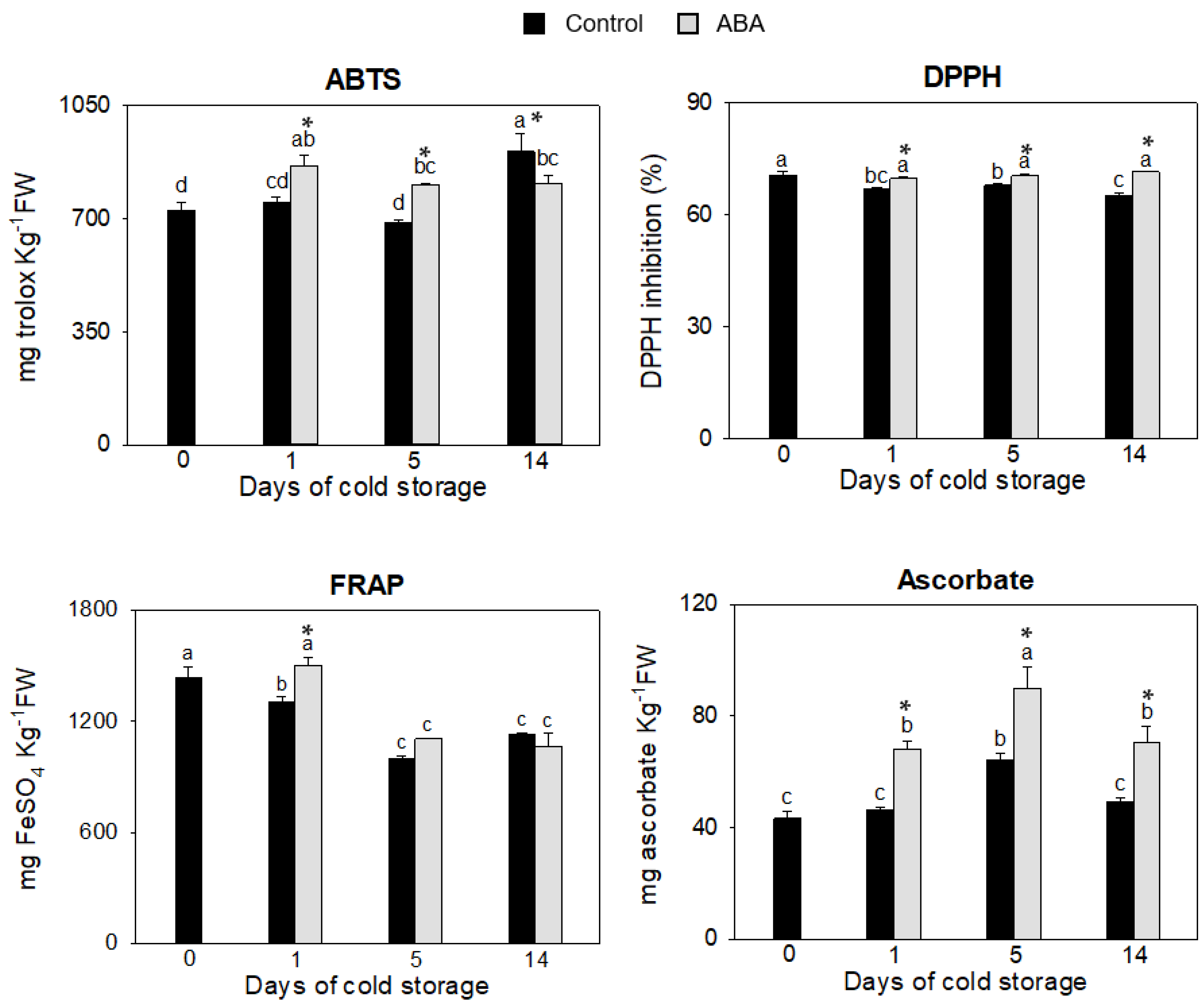
2.2. TEAC, DPPH and FRAP Assays
2.3. Ascorbate Content
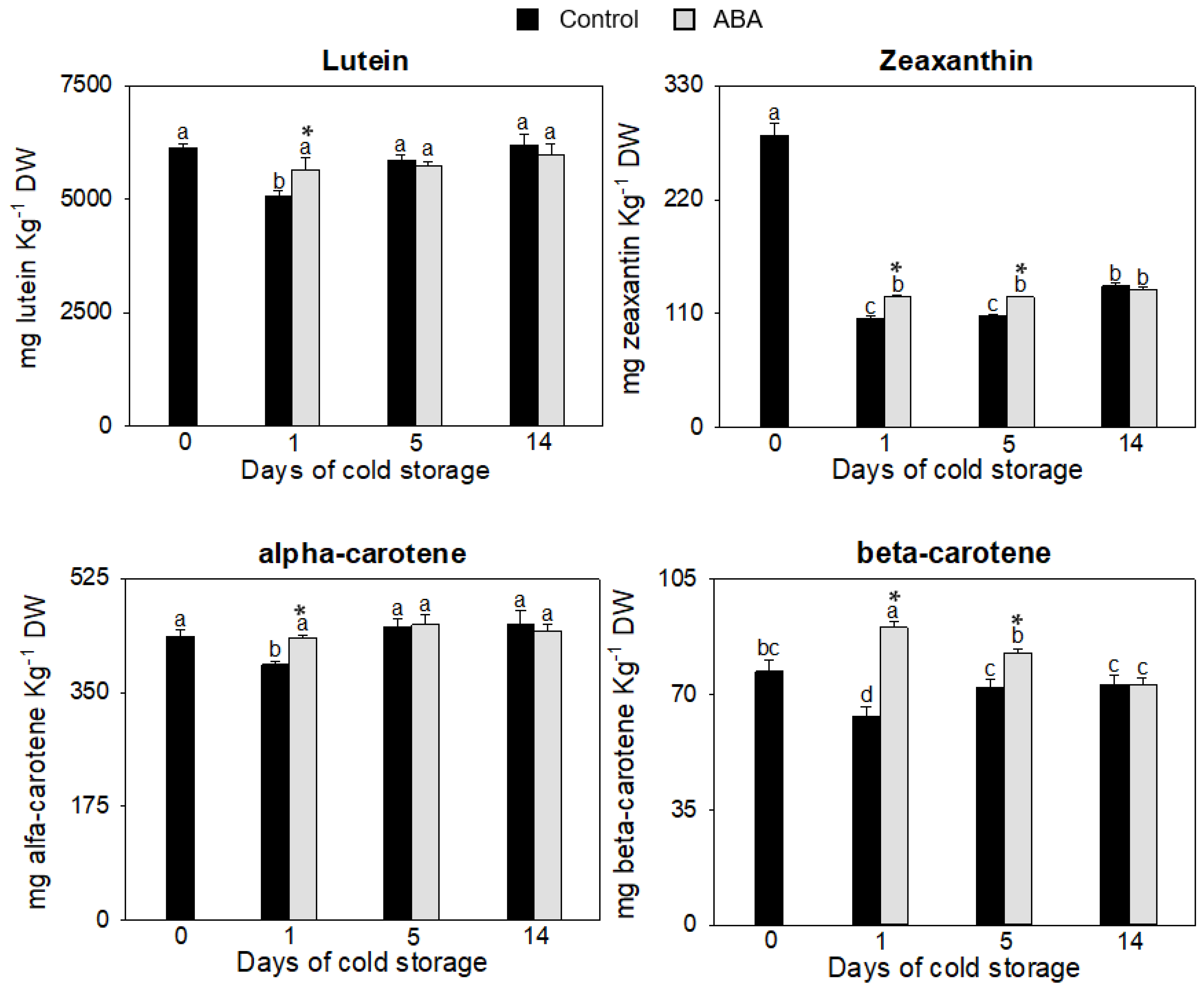
2.4. Carotenoids Quantification
2.5. Polyphenolic Metabolism
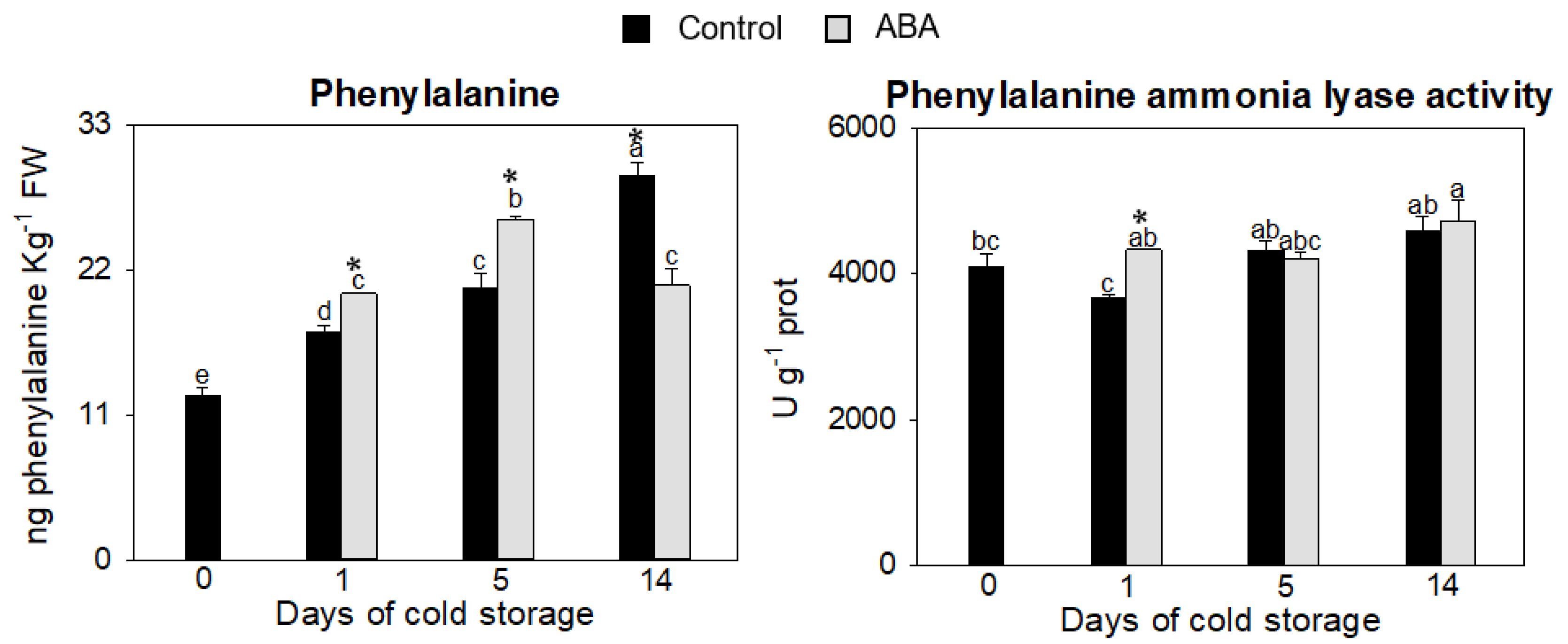
2.5.1. Phenylalanine Content
2.5.2. Soluble Phenolic Compounds Content
2.5.3. Flavonoid Content
2.5.4. Phenylalanine Ammonia Lyase Activity
2.5.5. Polyphenol Oxidase and Peroxidase Activities
2.5.6. Protein Determination
2.6. Quantification of Soluble Phenolic Compounds
2.7. Statistics
3. Results and Discussion
3.1. Evaluation of ABA Effect on Non-Enzymatic Antioxidants
3.1.1. Effect of ABA in the Phenylpropanoid Pathway
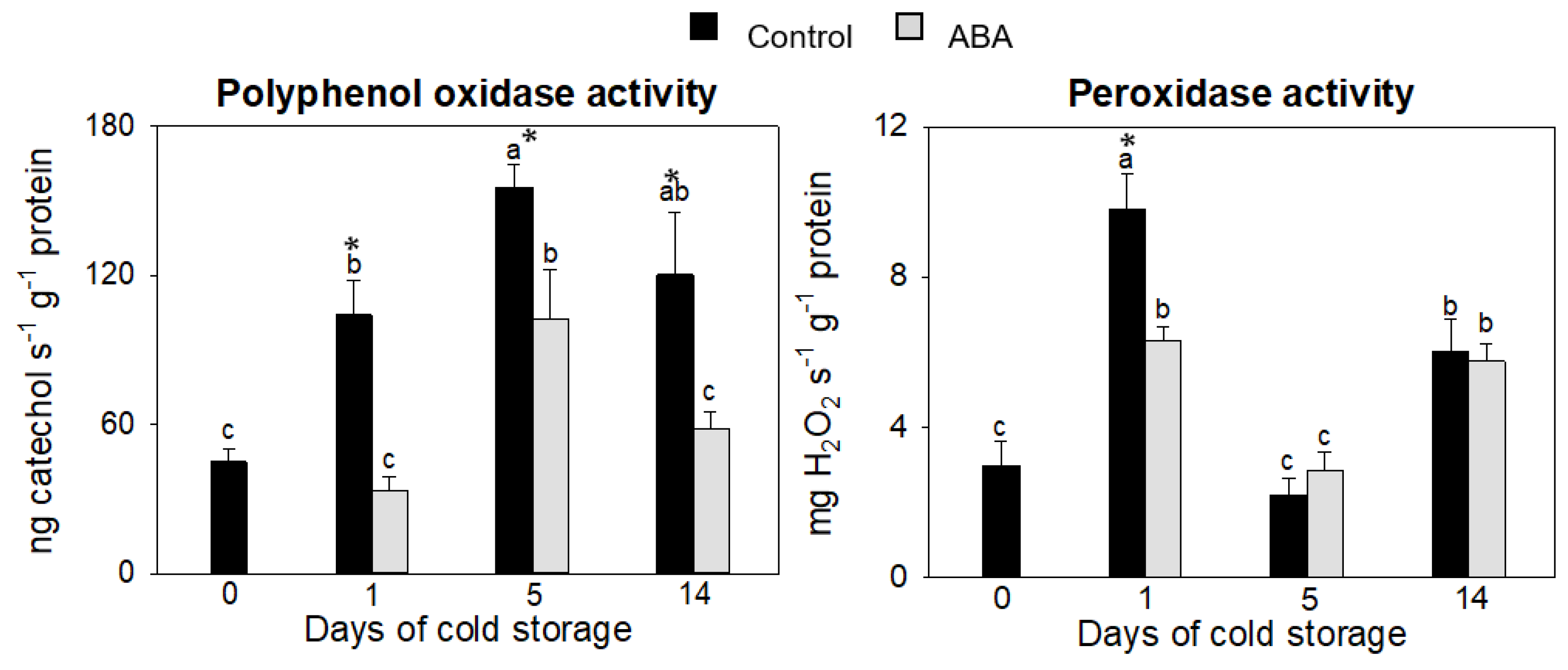
3.1.2. Effect of ABA on Phenol-Oxidizing Enzymes
3.2. Evaluation of ABA Effect on Individual Phenolic Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guajardo, E.; Correa, J.A.; Contreras-Porcia, L. Role of Abscisic Acid (ABA) in Activating Antioxidant Tolerance Responses to Desiccation Stress in Intertidal Seaweed Species. Planta 2016, 243, 767–781. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Q.; Asim, M.; Zhang, R.; Khan, R.; Farooq, S.; Wu, J. Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress. Biomolecules 2021, 11, 1159. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Tang, J.; Brummell, D.A.; Song, C.; Qi, S.; Lin, Q.; Bi, J.; Duan, Y. Abscisic Acid Alleviates Chilling Injury in Cold-Stored Peach Fruit by Regulating the Metabolism of Sucrose. Sci. Hortic. 2022, 298, 111000. [Google Scholar] [CrossRef]
- Carvajal, F.; Castro-Cegrí, A.; Jiménez-Muñoz, R.; Jamilena, M.; Garrido, D.; Palma, F. Changes in Morphology, Metabolism and Composition of Cuticular Wax in Zucchini Fruit During Postharvest Cold Storage. Front. Plant Sci. 2021, 12, 778745. [Google Scholar] [CrossRef]
- Carvajal, F.; Martinez, C.; Jamilena, M.; Garrido, D. Differential Response of Zucchini Varieties to Low Storage Temperature. Sci. Hortic. 2011, 130, 90–96. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Lluch, C.; Jamilena, M.; Garrido, D. Changes in Carbohydrate Content in Zucchini Fruit (Cucurbita Pepo L.) under Low Temperature Stress. Plant Sci. 2014, 217–218, 78–86. [Google Scholar] [CrossRef]
- Carvajal, F.; Palma, F.; Jamilena, M.; Garrido, D. Preconditioning Treatment Induces Chilling Tolerance in Zucchini Fruit Improving Different Physiological Mechanisms against Cold Injury. Ann. Appl. Biol. 2015, 166, 340–354. [Google Scholar] [CrossRef]
- Jiménez-Muñoz, R.; Palma, F.; Carvajal, F.; Castro-Cegrí, A.; Pulido, A.; Jamilena, M.; Romero-Puertas, M.C.; Garrido, D. Pre-Storage Nitric Oxide Treatment Enhances Chilling Tolerance of Zucchini Fruit (Cucurbita Pepo L.) by S-Nitrosylation of Proteins and Modulation of the Antioxidant Response. Postharvest Biol. Technol. 2021, 171, 111345. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Jamilena, M.; Garrido, D. Contribution of Polyamines and Other Related Metabolites to the Maintenance of Zucchini Fruit Quality during Cold Storage. Plant Physiol. Biochem. 2014, 82, 161–171. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Jiménez-Muñoz, R.; Pulido, A.; Jamilena, M.; Garrido, D. Exogenous γ-Aminobutyric Acid Treatment Improves the Cold Tolerance of Zucchini Fruit during Postharvest Storage. Plant Physiol. Biochem. 2019, 136, 188–195. [Google Scholar] [CrossRef]
- Carvajal, F.; Palma, F.; Jiménez-Muñoz, R.; Jamilena, M.; Pulido, A.; Garrido, D. Unravelling the Role of Abscisic Acid in Chilling Tolerance of Zucchini during Postharvest Cold Storage. Postharvest Biol. Technol. 2017, 133, 26–35. [Google Scholar] [CrossRef]
- Benítez, Á.; Iglesias-Moya, J.; Segura, M.; Carvajal, F.; Palma, F.; Garrido, D.; Martínez, C.; Jamilena, M. RNA-Seq Based Analysis of Transcriptomic Changes Associated with ABA-Induced Postharvest Cold Tolerance in Zucchini Fruit. Postharvest Biol. Technol. 2022, 192, 112023. [Google Scholar] [CrossRef]
- Parvin, K.; Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Mohsin, S.M.; Fujita, M. Comparative Physiological and Biochemical Changes in Tomato (Solanum Lycopersicum L.) under Salt Stress and Recovery: Role of Antioxidant Defense and Glyoxalase Systems. Antioxidants 2019, 8, 350. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, J.L.; Manzano, S.; Palma, F.; Carvajal, F.; Garrido, D.; Jamilena, M. Oxidative Stress Associated with Chilling Injury in Immature Fruit: Postharvest Technological and Biotechnological Solutions. Int. J. Mol. Sci. 2017, 18, 1467. [Google Scholar] [CrossRef] [PubMed]
- Avashthi, H.; Pathak, R.K.; Pandey, N.; Arora, S.; Mishra, A.K.; Gupta, V.K.; Ramteke, P.W.; Kumar, A. Transcriptome-Wide Identification of Genes Involved in Ascorbate–Glutathione Cycle (Halliwell–Asada Pathway) and Related Pathway for Elucidating Its Role in Antioxidative Potential in Finger Millet (Eleusine Coracana (L.)). 3 Biotech 2018, 8, 499. [Google Scholar] [CrossRef]
- Li, Z.; Li, B.; Li, M.; Fu, X.; Zhao, X.; Min, D.; Li, F.; Li, X.; Zhang, X. Hot Air Pretreatment Alleviates Browning of Fresh-Cut Pitaya Fruit by Regulating Phenylpropanoid Pathway and Ascorbate-Glutathione Cycle. Postharvest Biol. Technol. 2022, 190, 111954. [Google Scholar] [CrossRef]
- Yao, M.; Ge, W.; Zhou, Q.; Zhou, X.; Luo, M.; Zhao, Y.; Wei, B.; Ji, S. Exogenous Glutathione Alleviates Chilling Injury in Postharvest Bell Pepper by Modulating the Ascorbate-Glutathione (AsA-GSH) Cycle. Food Chem. 2021, 352, 129458. [Google Scholar] [CrossRef]
- Díaz-Mula, H.M.; Zapata, P.J.; Guillén, F.; Martínez-Romero, D.; Castillo, S.; Serrano, M.; Valero, D. Changes in Hydrophilic and Lipophilic Antioxidant Activity and Related Bioactive Compounds during Postharvest Storage of Yellow and Purple Plum Cultivars. Postharvest Biol. Technol. 2009, 51, 354–363. [Google Scholar] [CrossRef]
- Buran, T.J.; Sandhu, A.K.; Azeredo, A.M.; Bent, A.H.; Williamson, J.G.; Gu, L. Effects of Exogenous Abscisic Acid on Fruit Quality, Antioxidant Capacities, and Phytochemical Contents of Southern High Bush Blueberries. Food Chem. 2012, 132, 1375–1381. [Google Scholar] [CrossRef]
- Diretto, G.; Frusciante, S.; Fabbri, C.; Schauer, N.; Busta, L.; Wang, Z.; Matas, A.J.; Fiore, A.; Rose, J.K.; Fernie, A.R.; et al. Manipulation of β-Carotene Levels in Tomato Fruits Results in Increased ABA Content and Extended Shelf Life. Plant Biotechnol. J. 2020, 18, 1185–1199. [Google Scholar] [CrossRef]
- Palma, F.; Carvajal, F.; Jamilena, M.; Garrido, D. Putrescine Treatment Increases the Antioxidant Response and Carbohydrate Content in Zucchini Fruit Stored at Low Temperature. Postharvest Biol. Technol. 2016, 118, 68–70. [Google Scholar] [CrossRef]
- Iqbal, N.; Sehar, Z.; Fatma, M.; Umar, S.; Sofo, A.; Khan, N.A. Nitric Oxide and Abscisic Acid Mediate Heat Stress Tolerance through Regulation of Osmolytes and Antioxidants to Protect Photosynthesis and Growth in Wheat Plants. Antioxidants 2022, 11, 372. [Google Scholar] [CrossRef]
- Parveen, A.; Ahmar, S.; Kamran, M.; Malik, Z.; Ali, A.; Riaz, M.; Abbasi, G.H.; Khan, M.; Sohail, A.B.; Rizwan, M.; et al. Abscisic Acid Signaling Reduced Transpiration Flow, Regulated Na+ Ion Homeostasis and Antioxidant Enzyme Activities to Induce Salinity Tolerance in Wheat (Triticum Aestivum L.) Seedlings. Environ. Technol. Innov. 2021, 24, 101808. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, B.; Zhang, C.; Yang, L.; Wang, Y.; Zhao, H. Effects of Exogenous Abscisic Acid (ABA) on Carotenoids and Petal Color in Osmanthus Fragrans ‘Yanhonggui’. Plants 2020, 9, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Luo, Z.; Mou, W.; Wang, Y.; Ying, T.; Mao, L. ABA and UV-C Effects on Quality, Antioxidant Capacity and Anthocyanin Contents of Strawberry Fruit (Fragaria Ananassa Duch.). Postharvest Biol. Technol. 2014, 90, 56–62. [Google Scholar] [CrossRef]
- Mattus-Araya, E.; Guajardo, J.; Herrera, R.; Moya-León, M.A. ABA Speeds Up the Progress of Color in Developing F. Chiloensis Fruit through the Activation of PAL, CHS and ANS, Key Genes of the Phenylpropanoid/Flavonoid and Anthocyanin Pathways. Int. J. Mol. Sci. 2022, 23, 3854. [Google Scholar] [CrossRef]
- Tao, X.; Wu, Q.; Aalim, H.; Li, L.; Mao, L.; Luo, Z.; Ying, T. Effects of Exogenous Abscisic Acid on Bioactive Components and Antioxidant Capacity of Postharvest Tomato during Ripening. Molecules 2020, 25, 1346. [Google Scholar] [CrossRef] [Green Version]
- Sala, J.M.; Sanchez-Ballesta, M.T.; Alférez, F.; Mulas, M.; Zacarias, L.; Lafuente, M.T. A Comparative Study of the Postharvest Performance of an ABA-Deficient Mutant of Oranges: II. Antioxidant Enzymatic System and Phenylalanine Ammonia-Lyase in Non-Chilling and Chilling Peel Disorders of Citrus Fruit. Postharvest Biol. Technol. 2005, 37, 232–240. [Google Scholar] [CrossRef]
- Miret, J.A.; Munné-Bosch, S. Abscisic Acid and Pyrabactin Improve Vitamin C Contents in Raspberries. Food Chem. 2016, 203, 216–223. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, F.; Carvajal, F.; Ramos, J.M.; Jamilena, M.; Garrido, D. Effect of Putrescine Application on Maintenance of Zucchini Fruit Quality during Cold Storage: Contribution of GABA Shunt and Other Related Nitrogen Metabolites. Postharvest Biol. Technol. 2015, 99, 131–140. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Oxidants and Antioxidants Part A; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar] [CrossRef]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of Total Flavonoid Content in Propolis by Two Complementary Colometric Methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar] [CrossRef]
- Civello, P.M.; Martínez, G.A.; Chaves, A.R.; Añón, M.C. Heat Treatments Delay Ripening and Postharvest Decay of Strawberry Fruit. J. Agric. Food Chem. 1997, 45, 4589–4594. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Mellidou, I.; Koukounaras, A.; Kostas, S.; Patelou, E.; Kanellis, A.K. Regulation of Vitamin C Accumulation for Improved Tomato Fruit Quality and Alleviation of Abiotic Stress. Genes 2021, 12, 694. [Google Scholar] [CrossRef]
- Abdelgawad, K.F.; El-Mogy, M.M.; Mohamed, M.I.A.; Garchery, C.; Stevens, R.G. Increasing Ascorbic Acid Content and Salinity Tolerance of Cherry Tomato Plants by Suppressed Expression of the Ascorbate Oxidase Gene. Agronomy 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Ke, W.; Shao, X.; Gong, Y.; Xu, F.; Wang, H. Effects of Postharvest Hot Air Treatment on Gene Expression Associated with Ascorbic Acid Metabolism in Peach Fruit. Plant Mol. Biol. Rep. 2014, 32, 881–887. [Google Scholar] [CrossRef]
- Panjai, L.; Röhlen-Schmittgen, S.; Ellenberger, J.; Noga, G.; Hunsche, M.; Fiebig, A. Effect of Postharvest Irradiation with Red Light on Epidermal Color and Carotenoid Concentration in Different Parts of Tomatoes. J. Food Meas. Charact. 2021, 15, 1737–1746. [Google Scholar] [CrossRef]
- Tavarini, S.; Degl’Innocenti, E.; Remorini, D.; Massai, R.; Guidi, L. Antioxidant Capacity, Ascorbic Acid, Total Phenols and Carotenoids Changes during Harvest and after Storage of Hayward Kiwifruit. Food Chem. 2008, 107, 282–288. [Google Scholar] [CrossRef]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Abscisic Acid Increases Carotenoid and Chlorophyll Concentrations in Leaves and Fruit of Two Tomato Genotypes. J. Am. Soc. Hortic. Sci. 2014, 139, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Rey, F.; Zacarías, L.; Rodrigo, M.J. Carotenoids, Vitamin C, and Antioxidant Capacity in the Peel of Mandarin Fruit in Relation to the Susceptibility to Chilling Injury during Postharvest Cold Storage. Antioxidants 2020, 9, 1296. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Ji, C.Y.; Kim, S.H.; Ke, Q.; Park, S.-C.; Kim, H.S.; Lee, H.-U.; Lee, J.S.; Park, W.S.; Ahn, M.-J.; et al. Suppression of the β-Carotene Hydroxylase Gene Increases β-Carotene Content and Tolerance to Abiotic Stress in Transgenic Sweetpotato Plants. Plant Physiol. Biochem. 2017, 117, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Chapter 9—Role and Regulation of Plants Phenolics in Abiotic Stress Tolerance: An Overview. In Plant Signaling Molecules; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 157–168. [Google Scholar] [CrossRef]
- Wang, L.; Shan, T.; Xie, B.; Ling, C.; Shao, S.; Jin, P.; Zheng, Y. Glycine Betaine Reduces Chilling Injury in Peach Fruit by Enhancing Phenolic and Sugar Metabolisms. Food Chem. 2019, 272, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, J.; Niu, X.; Deng, C.; Song, X.; Li, W.; Cheng, Z.; Xu, Q.; Zhang, B.; Guo, W. GhANN1 Modulates the Salinity Tolerance by Regulating ABA Biosynthesis, Ion Homeostasis and Phenylpropanoid Pathway in Cotton. Environ. Exp. Bot. 2021, 185, 104427. [Google Scholar] [CrossRef]
- Barros, J.; Serrani-Yarce, J.C.; Chen, F.; Baxter, D.; Venables, B.J.; Dixon, R.A. Role of Bifunctional Ammonia-Lyase in Grass Cell Wall Biosynthesis. Nat. Plants 2016, 2, 16050. [Google Scholar] [CrossRef]
- Vighi, I.L.; Crizel, R.L.; Perin, E.C.; Rombaldi, C.V.; Galli, V. Crosstalk During Fruit Ripening and Stress Response Among Abscisic Acid, Calcium-Dependent Protein Kinase and Phenylpropanoid. Crit. Rev. Plant Sci. 2019, 38, 99–116. [Google Scholar] [CrossRef]
- Villalobos-González, L.; Peña-Neira, A.; Ibáñez, F.; Pastenes, C. Long-Term Effects of Abscisic Acid (ABA) on the Grape Berry Phenylpropanoid Pathway: Gene Expression and Metabolite Content. Plant Physiol. Biochem. 2016, 105, 213–223. [Google Scholar] [CrossRef]
- Kumar Patel, M.; Fanyuk, M.; Feyngenberg, O.; Maurer, D.; Sela, N.; Ovadia, R.; Oren-Shamir, M.; Alkan, N. Phenylalanine Induces Mango Fruit Resistance against Chilling Injuries during Storage at Suboptimal Temperature. Food Chem. 2023, 405, 134909. [Google Scholar] [CrossRef]
- Markhart, A.H. Chilling Injury: A Review of Possible Causes. HortScience 1986, 21, 1329–1333. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Moradi, M.; Razavi, F.; Rabiei, V. Exogenous Phenylalanine Application Promotes Chilling Tolerance in Tomato Fruits during Cold Storage by Ensuring Supply of NADPH for Activation of ROS Scavenging Systems. Sci. Hortic. 2019, 246, 818–825. [Google Scholar] [CrossRef]
- Cao, S.F.; Hu, Z.C.; Wang, H.O. Effect of Salicylic Acid on the Activities of Anti-Oxidant Enzymes and Phenylalanine Ammonia-Lyase in Cucumber Fruit in Relation to Chilling Injury. J. Hortic. Sci. Biotechnol. 2009, 84, 125–130. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, W.; Liu, J.; Liu, H.; Lv, Z.; Zhang, C.; Chen, D.; Jiao, Z. Postharvest UV-C Irradiation Increased the Flavonoids and Anthocyanins Accumulation, Phenylpropanoid Pathway Gene Expression, and Antioxidant Activity in Sweet Cherries (Prunus Avium L.). Postharvest Biol. Technol. 2021, 175, 111490. [Google Scholar] [CrossRef]
- Fan, S.; Li, Q.; Feng, S.; Lei, Q.; Abbas, F.; Yao, Y.; Chen, W.; Li, X.; Zhu, X. Melatonin Maintains Fruit Quality and Reduces Anthracnose in Postharvest Papaya via Enhancement of Antioxidants and Inhibition of Pathogen Development. Antioxidants 2022, 11, 804. [Google Scholar] [CrossRef] [PubMed]
- Carmona, L.; Sulli, M.; Diretto, G.; Alquezar, B.; Alves, M.; Pena, L. Improvement of Antioxidant Properties in Fruit from Two Blood and Blond Orange Cultivars by Postharvest Storage at Low Temperature. Antioxidants 2022, 11, 547. [Google Scholar] [CrossRef]
- Yoruk, R.; Marshall, M.R. Physicochemical Properties and Function of Plant Polyphenol Oxidase: A Review1. J. Food Biochem. 2003, 27, 361–422. [Google Scholar] [CrossRef]
- Cenobio-Galindo, A.D.J.; Ocampo-López, J.; Reyes-Munguía, A.; Carrillo-Inungaray, M.L.; Cawood, M.; Medina-Pérez, G.; Fernández-Luqueño, F.; Campos-Montiel, R.G. Influence of Bioactive Compounds Incorporated in a Nanoemulsion as Coating on Avocado Fruits (Persea Americana) during Postharvest Storage: Antioxidant Activity, Physicochemical Changes and Structural Evaluation. Antioxidants 2019, 8, 500. [Google Scholar] [CrossRef]
- Pasquariello, M.S.; Di Patre, D.; Mastrobuoni, F.; Zampella, L.; Scortichini, M.; Petriccione, M. Influence of Postharvest Chitosan Treatment on Enzymatic Browning and Antioxidant Enzyme Activity in Sweet Cherry Fruit. Postharvest Biol. Technol. 2015, 109, 45–56. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, W.; Liu, S.; Liu, C.; Zheng, L. Effects of Melatonin Treatment on the Enzymatic Browning and Nutritional Quality of Fresh-Cut Pear Fruit. Food Chem. 2019, 299, 125116. [Google Scholar] [CrossRef]
- Nazoori, F.; Mollai, S.; Sobhani, F.; Mirdehghan, S.H.; Sahhafi, S.R. Carboxymethyl Cellulose and Carnauba Wax Treatments Kept the Pomegranate Fruit (Punica Granatum L.) Quality during Cold Storage via Improving Enzymatic Defense System and Bioactive Compounds. Sci. Hortic. 2023, 309, 111645. [Google Scholar] [CrossRef]
- Sandhu, A.K.; Gray, D.J.; Lu, J.; Gu, L. Effects of Exogenous Abscisic Acid on Antioxidant Capacities, Anthocyanins, and Flavonol Contents of Muscadine Grape (Vitis Rotundifolia) Skins. Food Chem. 2011, 126, 982–988. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, S.Y. Antioxidant Activity and Phenolic Compounds in Selected Herbs. J. Agric. Food Chem. 2001, 49, 5165–5170. [Google Scholar] [CrossRef] [PubMed]
- Mason, T.J.; Bettenhausen, H.M.; Chaparro, J.M.; Uchanski, M.E.; Prenni, J.E. Evaluation of Ambient Mass Spectrometry Tools for Assessing Inherent Postharvest Pepper Quality. Hortic. Res. 2021, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Flores, G.; Blanch, G.P.; del Castillo, M.L.R. Effect of Postharvest Methyl Jasmonate Treatment on Fatty Acid Composition and Phenolic Acid Content in Olive Fruits during Storage. J. Sci. Food Agric. 2017, 97, 2767–2772. [Google Scholar] [CrossRef]
- Vithana, M.D.; Singh, Z.; Johnson, S.K. Dynamics in the Concentrations of Health-Promoting Compounds: Lupeol, Mangiferin and Different Phenolic Acids during Postharvest Ripening of Mango Fruit. J. Sci. Food Agric. 2018, 98, 1460–1468. [Google Scholar] [CrossRef]
- Osondu, H.A.A.; Akinola, S.A.; Shoko, T.; Pillai, S.K.; Sivakumar, D. Coating Properties, Resistance Response, Molecular Mechanisms and Anthracnose Decay Reduction in Green Skin Avocado Fruit (‘Fuerte’) Coated with Chitosan Hydrochloride Loaded with Functional Compounds. Postharvest Biol. Technol. 2022, 186, 111812. [Google Scholar] [CrossRef]
- Mattila, P.; Hellström, J.; Törrönen, R. Phenolic Acids in Berries, Fruits, and Beverages. J. Agric. Food Chem. 2006, 54, 7193–7199. [Google Scholar] [CrossRef]
- Rasheed, H.; Ahmad, D.; Bao, J. Genetic Diversity and Health Properties of Polyphenols in Potato. Antioxidants 2022, 11, 603. [Google Scholar] [CrossRef]
- Zhao, Z.; Moghadasian, M.H. Chemistry, Natural Sources, Dietary Intake and Pharmacokinetic Properties of Ferulic Acid: A Review. Food Chem. 2008, 109, 691–702. [Google Scholar] [CrossRef]
- Baraiya, N.S.; Rao, T.V.R.; Thakkar, V.R. Composite Coating as a Carrier of Antioxidants Improves the Postharvest Shelf Life and Quality of Table Grapes (Vitis Vinifera L. Var. Thompson Seedless). J. Agric. Sci. Technol. 2016, 18, 93–107. [Google Scholar]
- Liu, H.; Huang, Z.; Lyu, L.; Fan, S.; Wu, W.; Li, W. Effect of Ferulic Acid and Natamycin Treatments on Quality and Reactive Oxygen Species Metabolism of Postharvest Blackberry. J. Food Nutr. Res. 2021, 60, 236–243. [Google Scholar]
- Liu, X.; Ji, D.; Cui, X.; Zhang, Z.; Li, B.; Xu, Y.; Chen, T.; Tian, S. p-Coumaric Acid Induces Antioxidant Capacity and Defense Responses of Sweet Cherry Fruit to Fungal Pathogens. Postharvest Biol. Technol. 2020, 169, 111297. [Google Scholar] [CrossRef]
- Yuan, S.; Li, W.; Li, Q.; Wang, L.; Cao, J.; Jiang, W. Defense Responses, Induced by p-Coumaric Acid and Methyl p-Coumarate, of Jujube (Ziziphus Jujuba Mill.) Fruit against Black Spot Rot Caused by Alternaria alternata. J. Agric. Food Chem. 2019, 67, 2801–2810. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.I.; Ferreres, F.; Tomás-Barberán, F.A. Effect of Postharvest Storage and Processing on the Antioxidant Constituents (Flavonoids and Vitamin C) of Fresh-Cut Spinach. J. Agric. Food Chem. 1999, 47, 2213–2217. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Muñoz, A.; Sánchez-Hernández, S.; Samaniego-Sánchez, C.; Giménez-Martínez, R.; Olalla-Herrera, M. Differences in the Phenolic Profile by UPLC Coupled to High Resolution Mass Spectrometry and Antioxidant Capacity of Two Diospyros Kaki Varieties. Antioxidants 2021, 10, 31. [Google Scholar] [CrossRef]
- Santin, M.; Lucini, L.; Castagna, A.; Rocchetti, G.; Hauser, M.-T.; Ranieri, A. Comparative “Phenol-Omics” and Gene Expression Analyses in Peach (Prunus Persica) Skin in Response to Different Postharvest UV-B Treatments. Plant Physiol. Biochem. 2019, 135, 511–519. [Google Scholar] [CrossRef]
- Santin, M.; Ranieri, A.; Hauser, M.-T.; Miras-Moreno, B.; Rocchetti, G.; Lucini, L.; Strid, Å.; Castagna, A. The Outer Influences the Inner: Postharvest UV-B Irradiation Modulates Peach Flesh Metabolome Although Shielded by the Skin. Food Chem. 2021, 338, 127782. [Google Scholar] [CrossRef]
- Mendoza, L.; Yañez, K.; Vivanco, M.; Melo, R.; Cotoras, M. Characterization of Extracts from Winery By-Products with Antifungal Activity against Botrytis Cinerea. Ind. Crop. Prod. 2013, 43, 360–364. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Schena, L.; De Girolamo, A.; Ippolito, A.; González-Candelas, L. Characterization of Genes Associated with Induced Resistance against Penicillium Expansum in Apple Fruit Treated with Quercetin. Postharvest Biol. Technol. 2010, 56, 1–11. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, L.; Zhang, L.; Guo, Y.; Qi, X.; He, L. Effects of Quercetin on Postharvest Blue Mold Control in Kiwifruit. Sci. Hortic. 2018, 228, 18–25. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Hou, Y.; Zhu, X.; Zheng, Y.; Jin, P. Physiological and Metabolomic Analyses of Hot Water Treatment on Amino Acids and Phenolic Metabolisms in Peach Cold Tolerance. Postharvest Biol. Technol. 2021, 179, 111593. [Google Scholar] [CrossRef]
- Liu, X.; Cui, X.; Ji, D.; Zhang, Z.; Li, B.; Xu, Y.; Chen, T.; Tian, S. Luteolin-Induced Activation of the Phenylpropanoid Metabolic Pathway Contributes to Quality Maintenance and Disease Resistance of Sweet Cherry. Food Chem. 2021, 342, 128309. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.-M. Luteolin, a Flavonoid with Potential for Cancer Prevention and Therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Flores, G.; del Castillo, M.L.R. Influence of Preharvest and Postharvest Methyl Jasmonate Treatments on Flavonoid Content and Metabolomic Enzymes in Red Raspberry. Postharvest Biol. Technol. 2014, 97, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.S.; Pérez-Gregorio, M.R.; García-Falcón, M.S.; Simal-Gándara, J.; Almeida, D.P.F. Effect of Post-Harvest Practices on Flavonoid Content of Red and White Onion Cultivars. Food Control 2010, 21, 878–884. [Google Scholar] [CrossRef]
- Zhao, J.; Ao, M.; He, X.; Li, W.; Deng, L.; Zeng, K.; Ming, J. Changes in Phenolic Content, Composition, and Antioxidant Activity of Blood Oranges during Cold and on-Tree Storage. J. Integr. Agric. 2022, 21, 3669–3683. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days of Storing at 4 °C | 1 | 5 | 14 | |||
|---|---|---|---|---|---|---|
| Control | ABA | Control | ABA | Control | ABA | |
| Weight loss (%) | 1.38 ± 0.1 e | 1.48 ± 0.06 e | 4.06 ± 0.1 c* | 3.37 ± 0.08 d | 11.99 ± 0.4 a* | 8.39 ± 0.7 b |
| Chilling Injury | 0 ± 0 c | 0 ± 0 c | 0.18 ± 0.06 c | 0.02 ± 0.02 c | 2.23 ± 0.2 a* | 0.69 ± 0.16 b |
| Days of Storing at 4 °C | 0 | 1 | 5 | 14 | |||
|---|---|---|---|---|---|---|---|
| At Harvest | Control | ABA | Control | ABA | Control | ABA | |
| Hydroxybenzoic acids | |||||||
| Vanillic acid (mg/kgDW) | 27.3 ± 2.4 Aab | 22.8 ± 1.5 AB | 22.5 ± 0.6 b | 20.4 ± 0.3 B | 27.9 ± 1.4 a* | 20.7 ± 0.2 B | 25.8 ± 1.4 ab* |
| Ellagic acid (µg/kgDW) | 5.7 ± 0.1 Bb | 19.8 ± 0.6 A | 19.8 ± 1 a | 18.6 ± 0.8 A | 18.1 ± 0.4 a | 18.7 ± 0.5 A | 18.3 ± 0.2 a |
| Hydroxycinnamic acids | |||||||
| Ferulic acid (mg/kgDW) | 8.9 ± 0.4 Bb | 3.6 ± 0.5 D | 3.3 ± 0.2 c | 5.4 ± 0.1 C | 8.7 ± 0.5 b* | 13.3 ± 0.5 A | 12.8 ± 0.7 a |
| Coumaric acid (µg/kgDW) | 21.3 ± 1.4 Aa | 25.2 ± 3.4 A | 25.8 ± 1 a | NDB | 13.5 ± 0.8 ab* | NDB | NDB |
| Flavonoids | |||||||
| Quercetin (mg/kgDW) | 187.0 ± 30.6 Aa | 147.1 ± 7.9 AB | 207.7 ± 13.9 a* | 181.5 ± 1.8 A | 196.8 ± 3.6 a* | 130.2 ± 9.3 B | 174.4 ± 18.5 a* |
| Isorhamnetin (µg/kgDW) | 30.5 ± 0.4 Bc | 31.9 ± 0.7 B | 35.6 ± 0.1 b* | 34.2 ± 0.1 B | 40.1 ± 3.7 ab* | 46.5 ± 4.9 A | 43.1 ± 1.3 a |
| Myricetin (mg/kgDW) | 2.8 ± 0.1 ABb | 2.7 ± 0.1 AB | 3.8 ± 0.1 a* | 3.0 ± 0.1 A | 2.9 ± 0.4 b | 2.3 ± 0.3 B | 2.6 ± 0.1 b |
| Naringenin (µg/kgDW) | 10.5 ± 1.4 Aa | 9.4 ± 0.8 A | 10.3 ± 0.8 a | 9.1 ± 0.7 A | 11.0 ± 0.3 a | 9.8 ± 0.8 A | 10.8 ± 0.6 a |
| Luteolin (µg/kgDW) | 16.5 ± 0.1 ABb | 20.8 ± 2.0 A | 43.6 ± 1.2 a* | 13.9 ± 1 B | 44.8 ± 6 a* | 12.6 ± 2.3 B | 39.8 ± 6.2 a* |
| Rhoifolin (µg/kgDW) | 16.9 ± 0.4 Aa | 8.8 ± 0.4 B | 11.5 ± 1.4 b | 7.8 ± 0.5 B | 11.0 ± 1.1* | 8.8 ± 3.2 B | 5.6 ± 0.5 b |
| Chrysoeriol (µg/kgDW) | 24.5 ± 0.3 Aa | 6.4 ± 0.5 B | 10.3 ± 0.2 b* | 4.5 ± 1.5 B | 5.9 ± 1.4 c | 0.3 ± 0.1 C | 4.8 ± 1.4 c* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro-Cegrí, A.; Sierra, S.; Hidalgo-Santiago, L.; Esteban-Muñoz, A.; Jamilena, M.; Garrido, D.; Palma, F. Postharvest Treatment with Abscisic Acid Alleviates Chilling Injury in Zucchini Fruit by Regulating Phenolic Metabolism and Non-Enzymatic Antioxidant System. Antioxidants 2023, 12, 211. https://doi.org/10.3390/antiox12010211
Castro-Cegrí A, Sierra S, Hidalgo-Santiago L, Esteban-Muñoz A, Jamilena M, Garrido D, Palma F. Postharvest Treatment with Abscisic Acid Alleviates Chilling Injury in Zucchini Fruit by Regulating Phenolic Metabolism and Non-Enzymatic Antioxidant System. Antioxidants. 2023; 12(1):211. https://doi.org/10.3390/antiox12010211
Chicago/Turabian StyleCastro-Cegrí, Alejandro, Sandra Sierra, Laura Hidalgo-Santiago, Adelaida Esteban-Muñoz, Manuel Jamilena, Dolores Garrido, and Francisco Palma. 2023. "Postharvest Treatment with Abscisic Acid Alleviates Chilling Injury in Zucchini Fruit by Regulating Phenolic Metabolism and Non-Enzymatic Antioxidant System" Antioxidants 12, no. 1: 211. https://doi.org/10.3390/antiox12010211
APA StyleCastro-Cegrí, A., Sierra, S., Hidalgo-Santiago, L., Esteban-Muñoz, A., Jamilena, M., Garrido, D., & Palma, F. (2023). Postharvest Treatment with Abscisic Acid Alleviates Chilling Injury in Zucchini Fruit by Regulating Phenolic Metabolism and Non-Enzymatic Antioxidant System. Antioxidants, 12(1), 211. https://doi.org/10.3390/antiox12010211