
Structural Optimization of Cannabidiol as Multifunctional Cosmetic Raw Materials

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
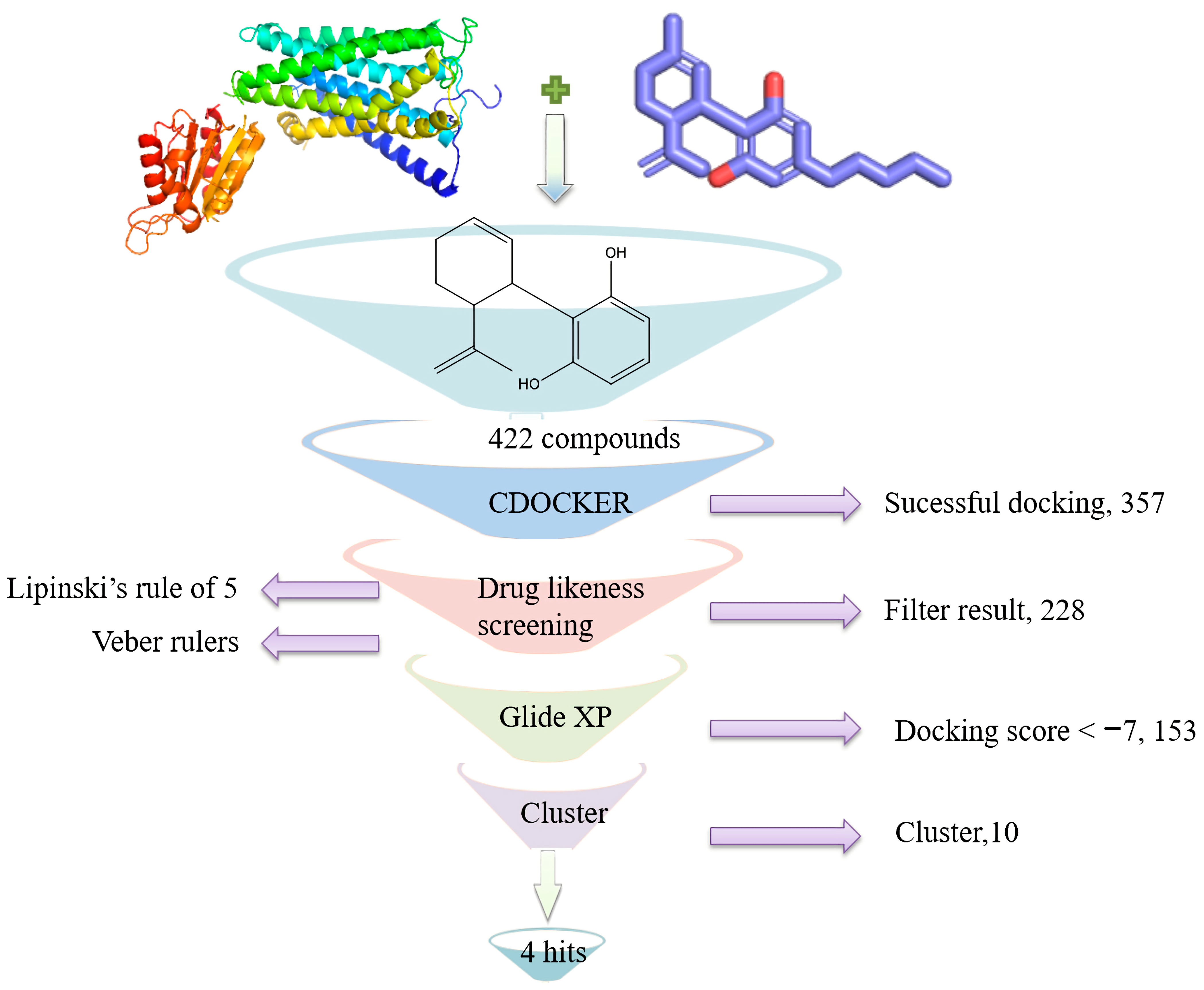
2.2. Computer-Aided Virtual Screening
2.2.1. CBD Substitutes Obtained and Primary Docking Screening
2.2.2. Drug Likeness Evaluation
2.2.3. High Precision Screening
2.2.4. Similarity Searching
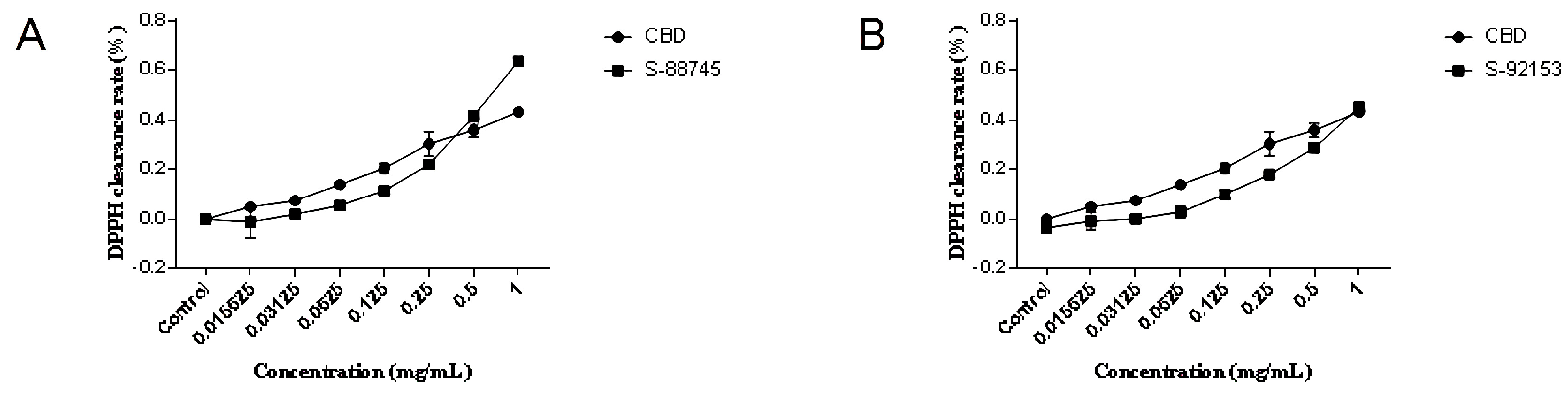
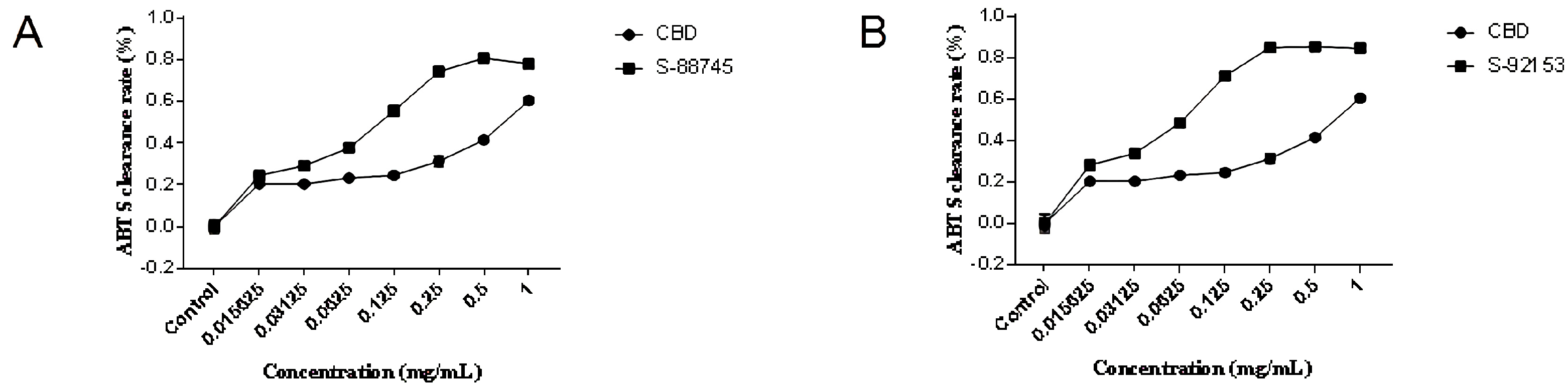
2.3. Antioxidant Capacity Evaluation
2.3.1. DPPH Radical Scavenging Activity
2.3.2. ABTS Radical Scavenging Activity
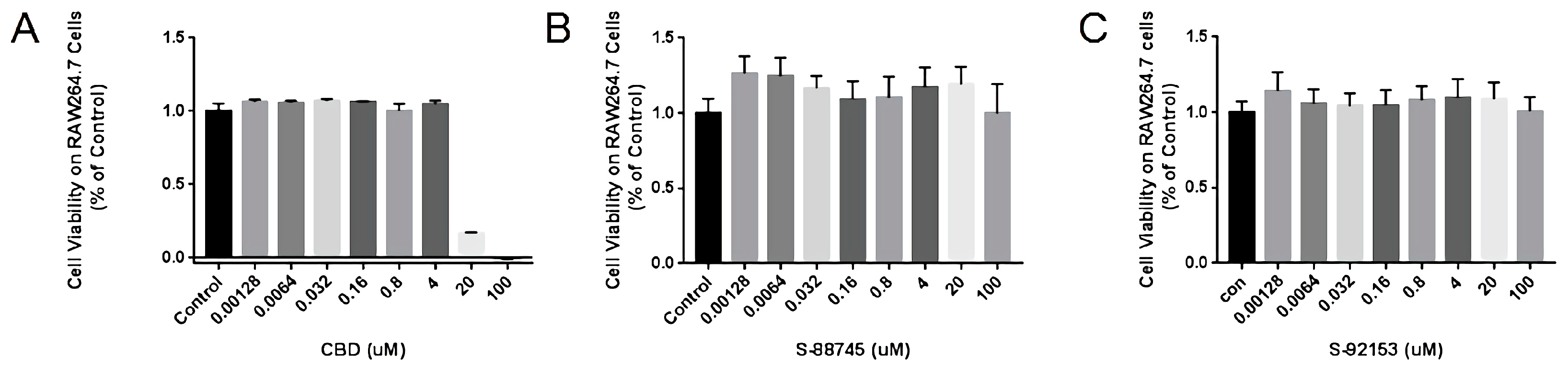
2.4. Cytotoxicity Measurement of CBD and Its Derivatives
2.5. Anti-Inflammatory Activity Test
2.5.1. Measurement of NO and IL-6 Levels
2.5.2. Western Blot Analysis
2.6. Antiwrinkle Activity Test
2.6.1. Repair Ultraviolet Radiation Damage Experiment
2.6.2. Detection of Collagen, Elastin and Fibronectin Levels
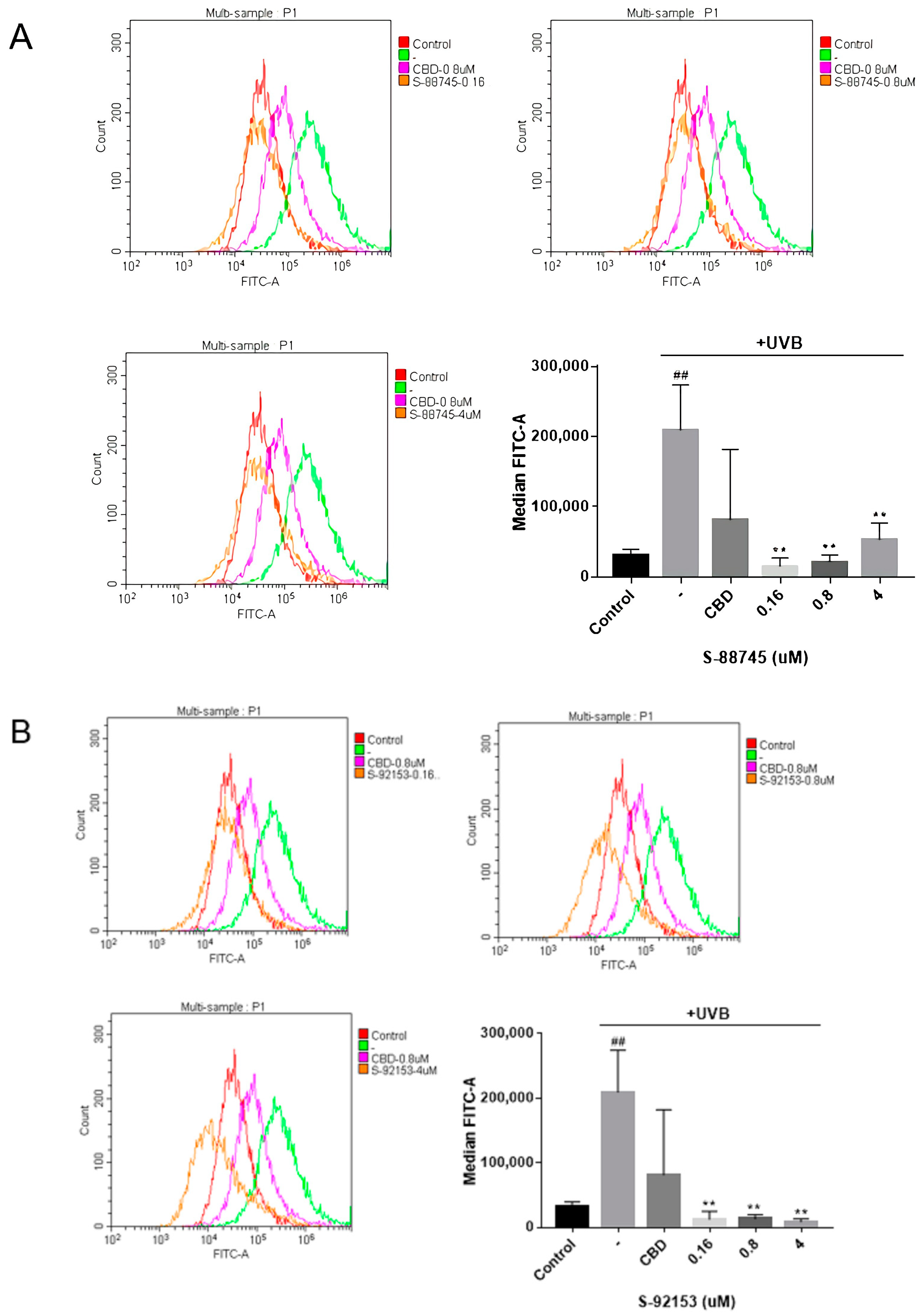
2.6.3. Measurement of the ROS Content
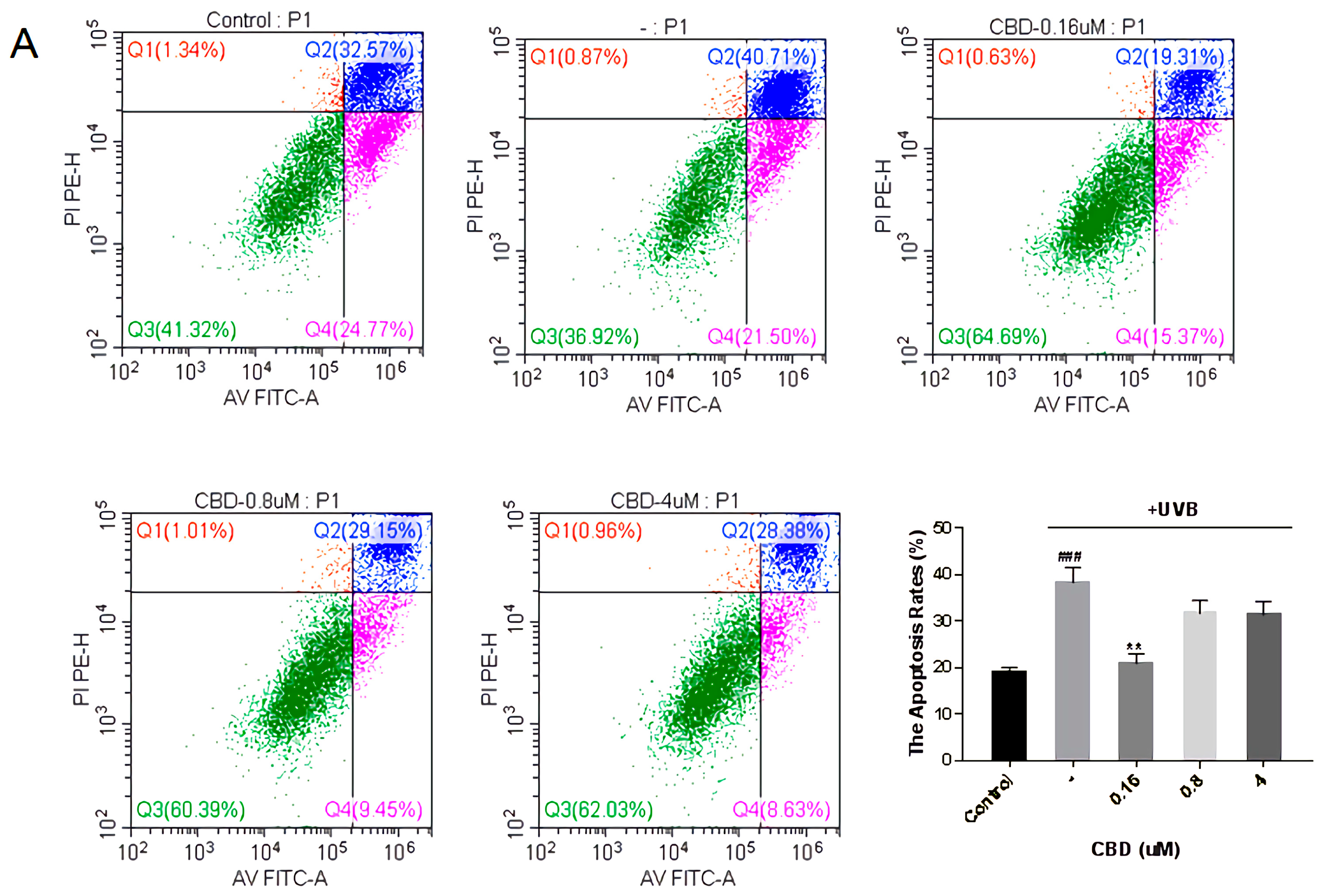
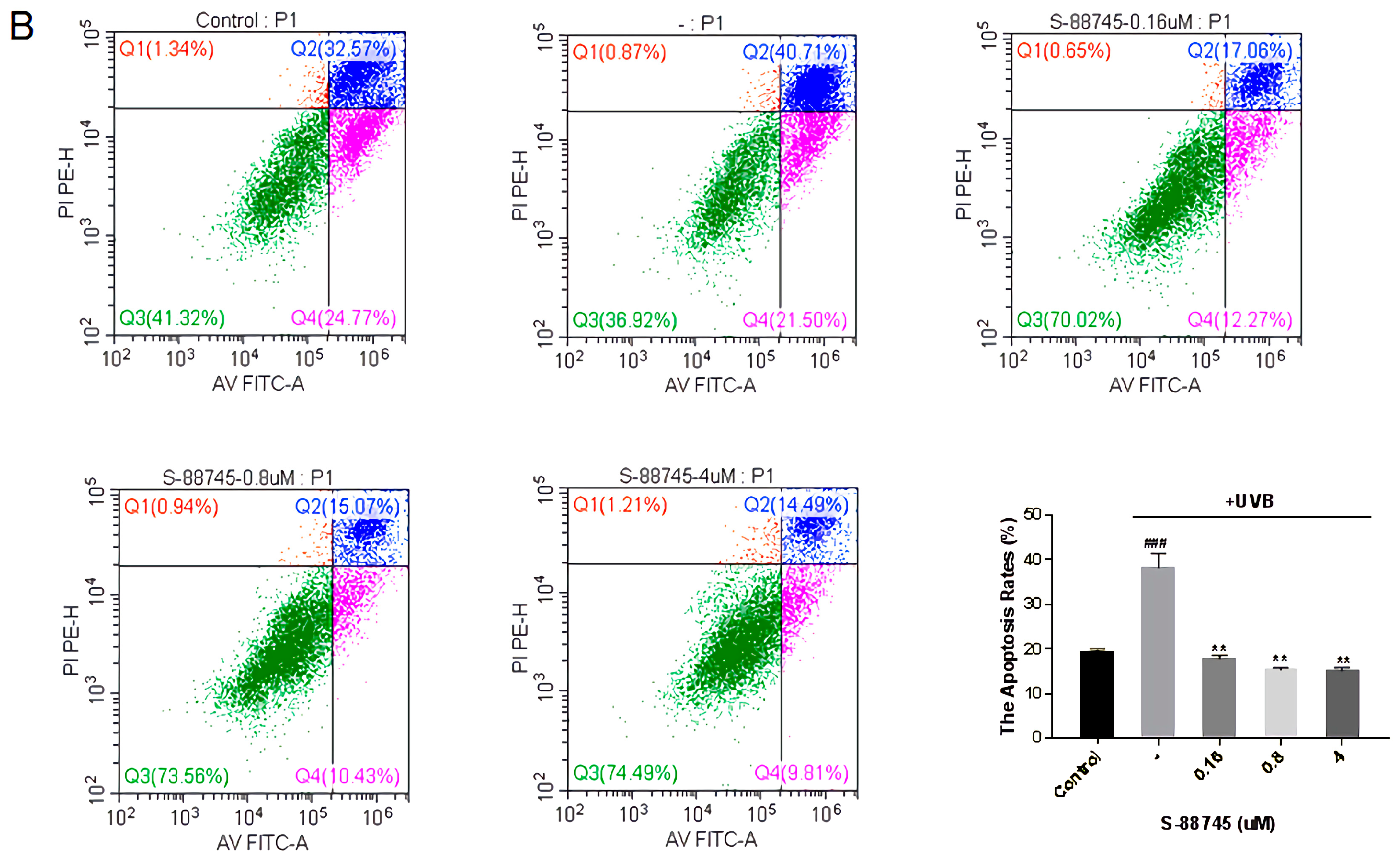
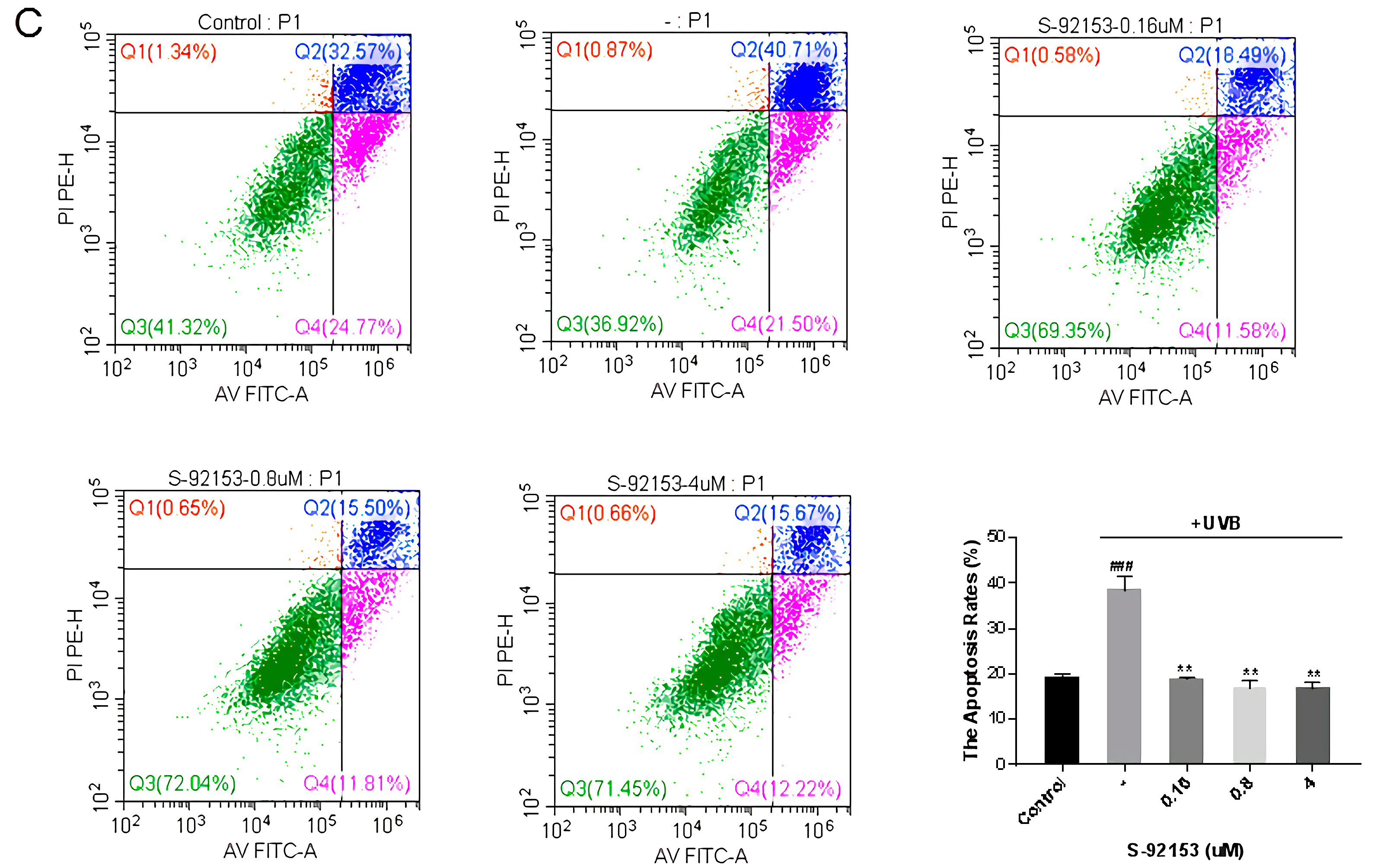
2.6.4. Detection the Effects on the Apoptosis of HFF-1 Cells
2.7. Determination of Melanin Content and Tyrosinase Activity
2.8. Statistical Analysis
3. Results
3.1. Virtual Screening
3.2. Extracellular Antioxidant Test
3.3. Anti-Inflammatory Activity Test
3.3.1. Cytotoxicities Measurement
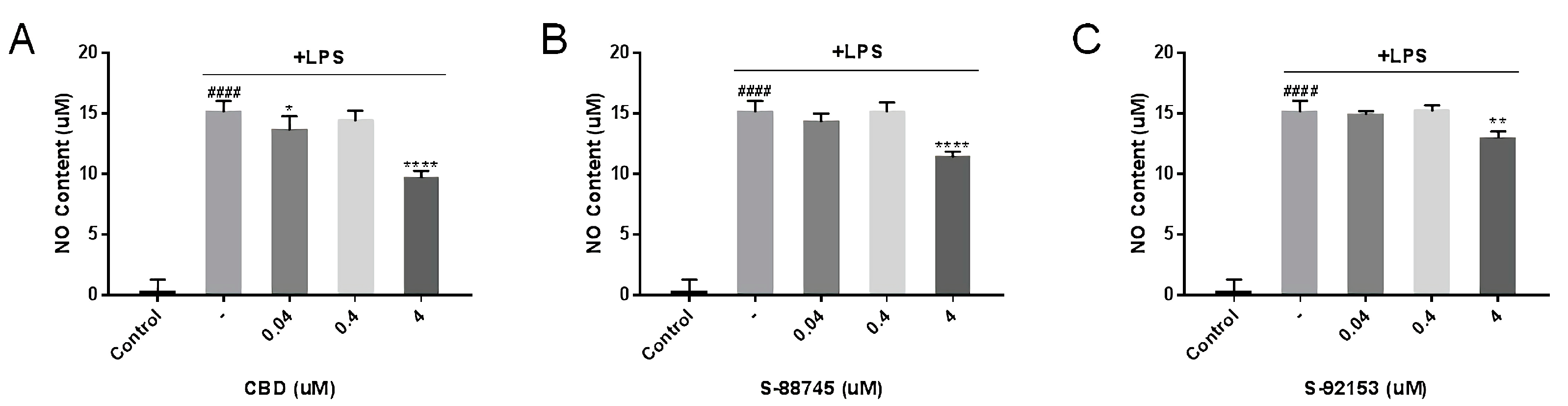
3.3.2. Suppression of NO
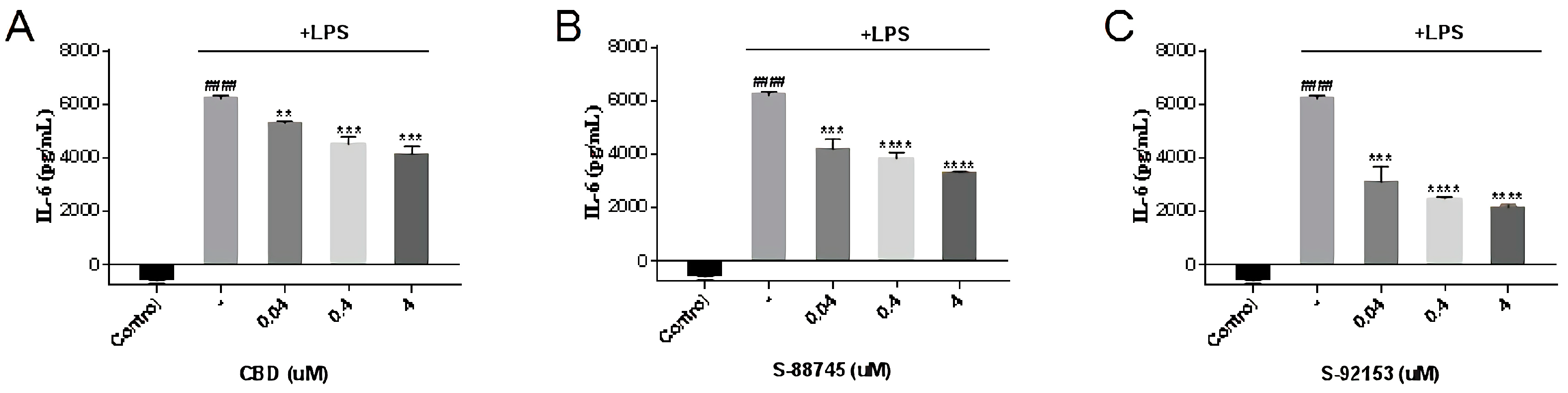
3.3.3. Inhibitory Effect on IL-6 Production
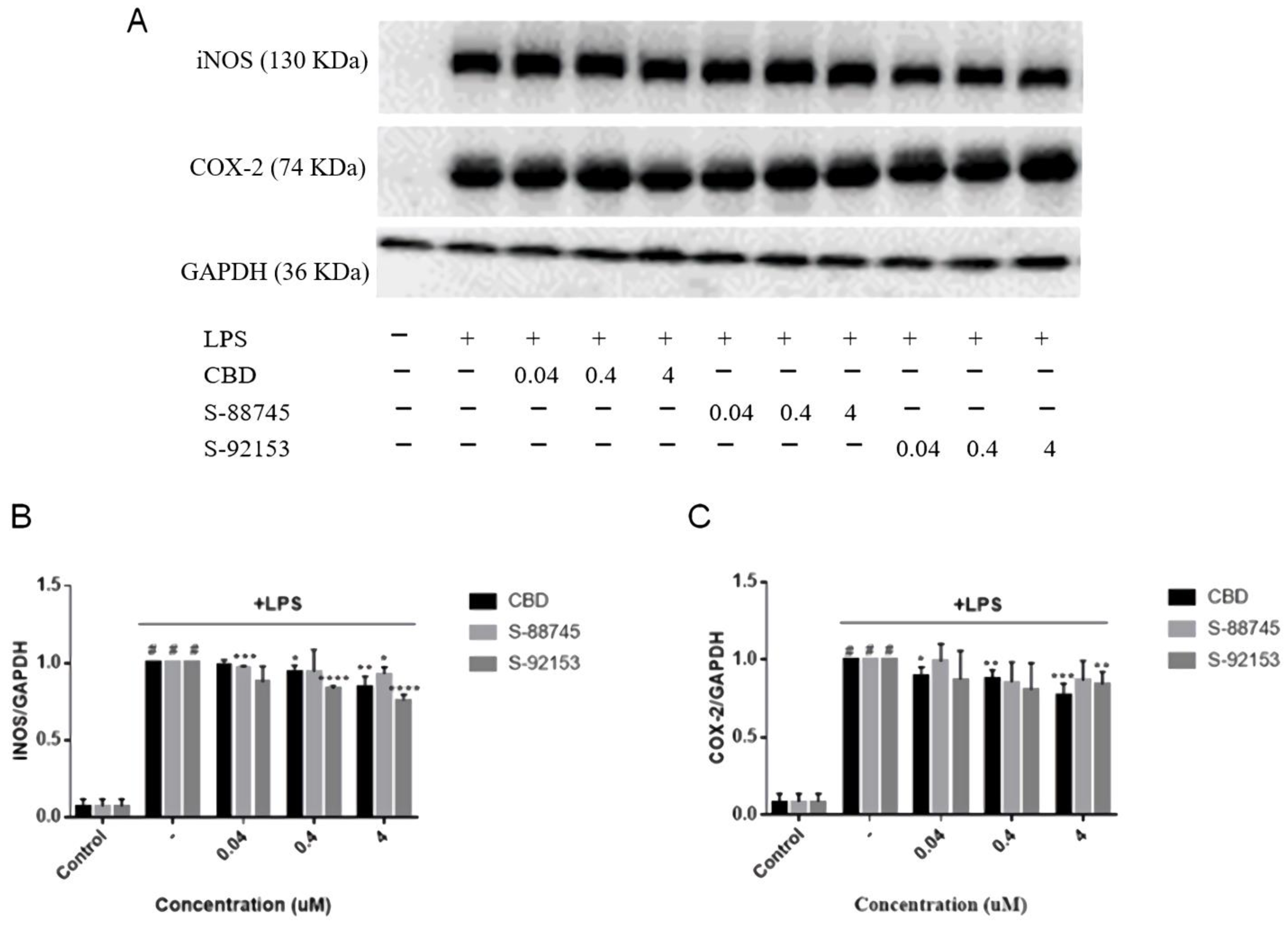
3.3.4. Effects on Protein Expression of iNOS and COX-2
3.4. Anti-Wrinkle Activity Test
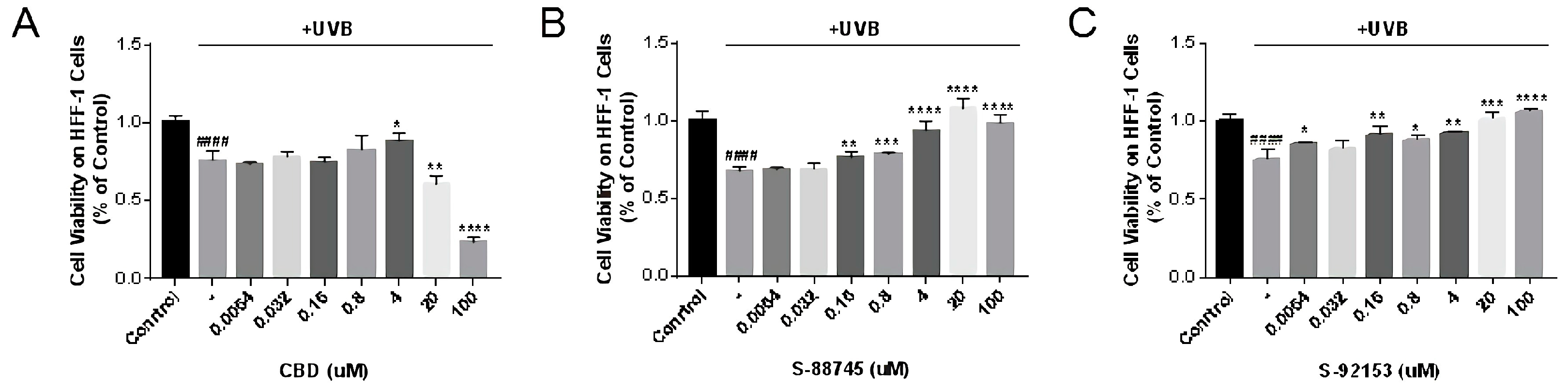
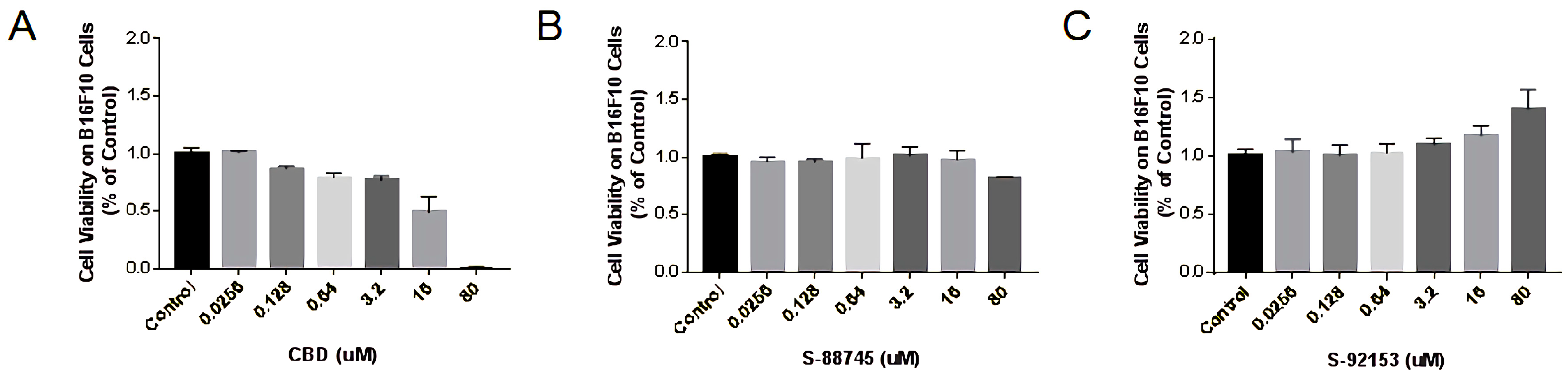
3.4.1. Cytotoxicity Measurement on HFF-1 Cell Line
3.4.2. Fibroblast Damage Repairment Evaluation
3.4.3. Intracellular Antioxidant Activity
3.4.4. Fibroblast Apoptosis Experiment
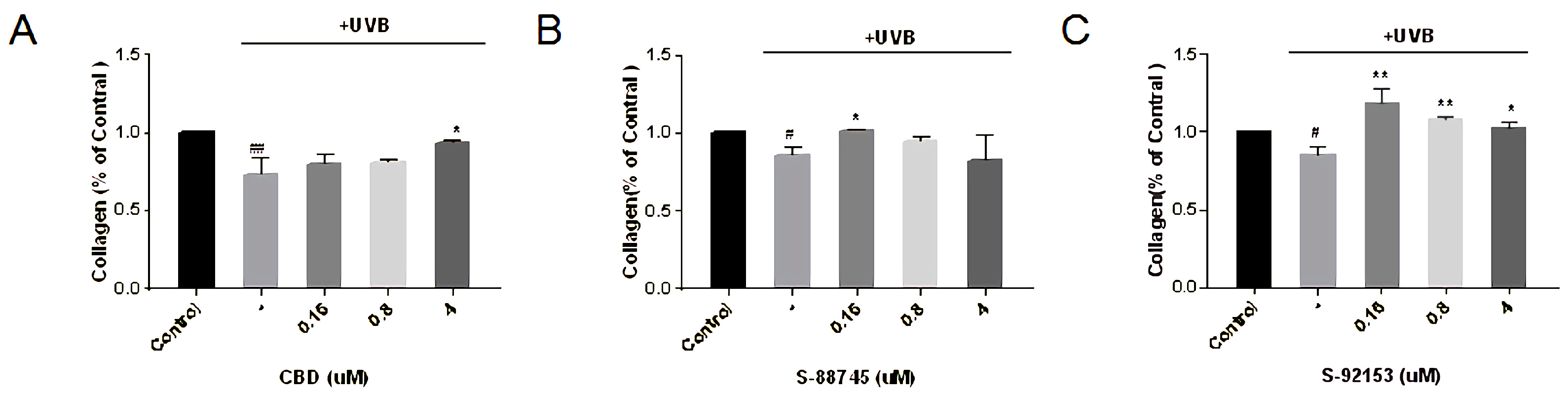
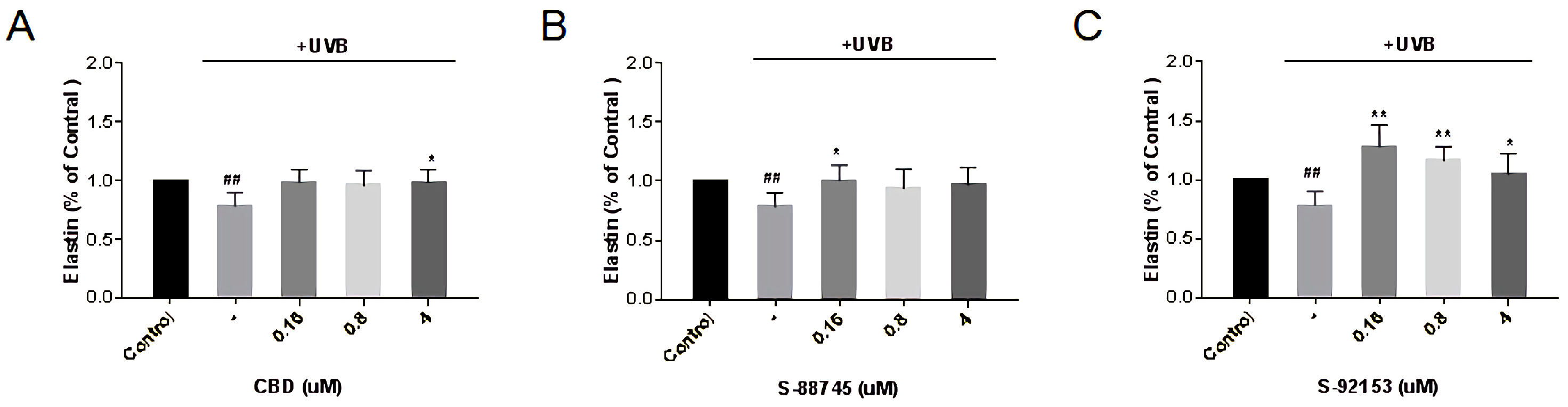
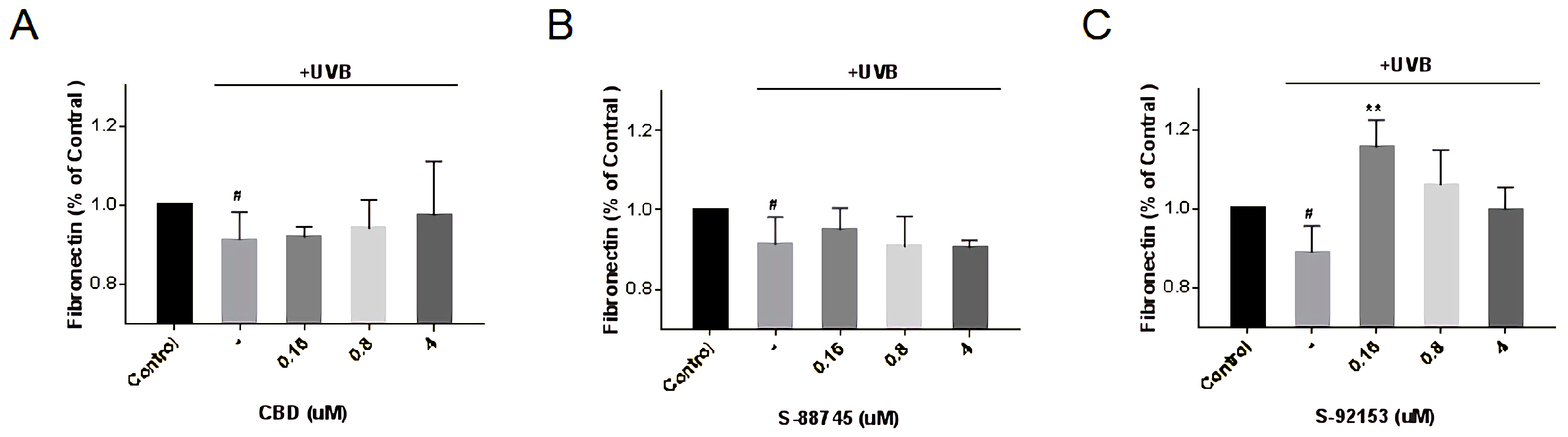
3.4.5. Effects Evaluation on Collagen, Elastin and Fibronectin Production
3.5. Whitening Activity Test
3.5.1. Cytotoxicity Measurements on B16F10
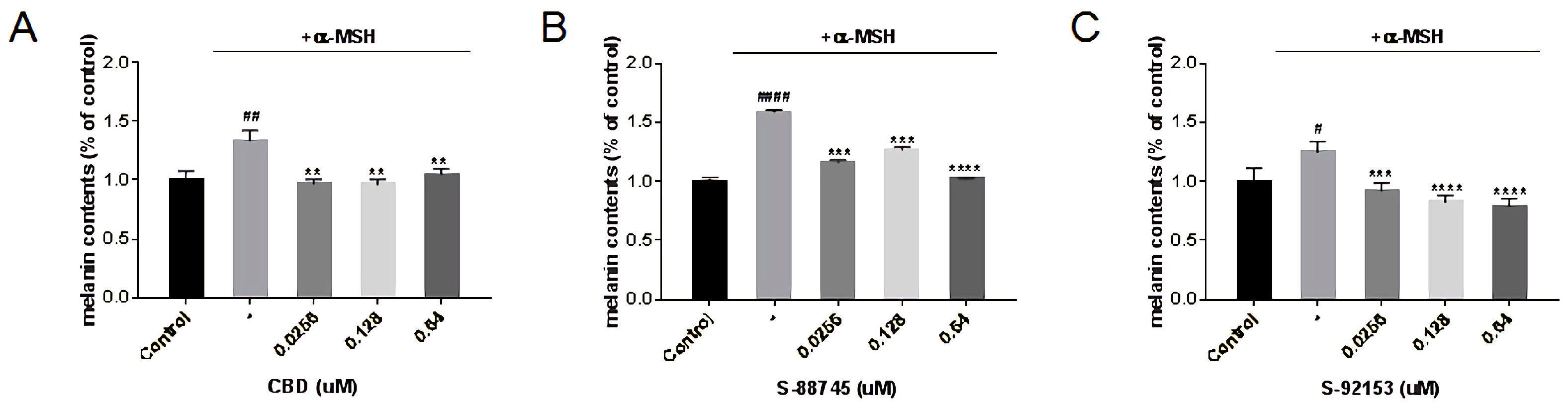
3.5.2. Whitening Activity Tests
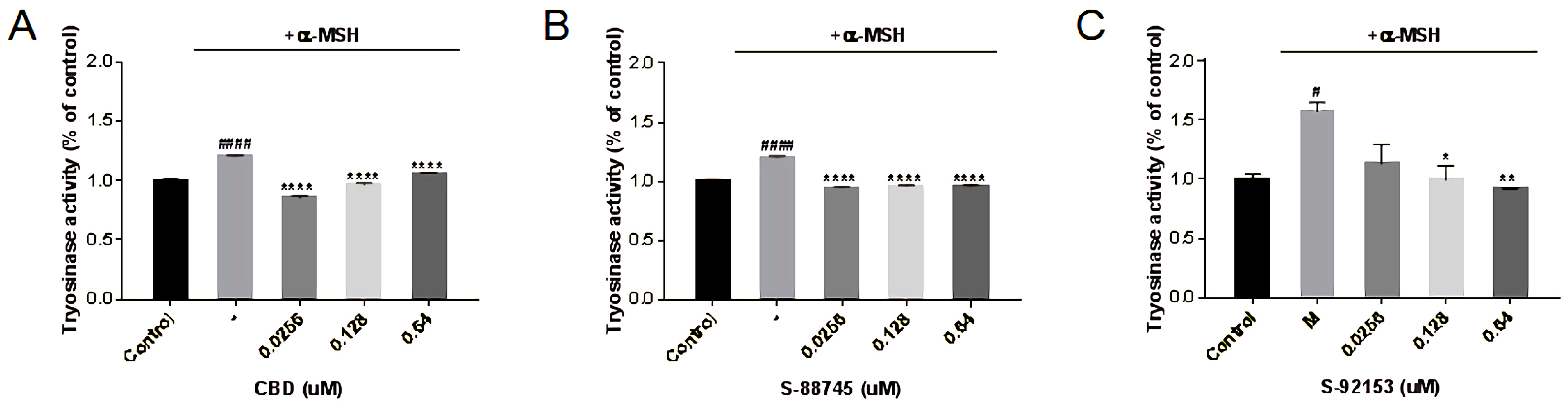
3.5.3. The Effects on Tyrosinase Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roh, E.; Kim, J.E.; Kwon, J.Y.; Park, J.S.; Bode, A.M.; Dong, Z.; Lee, K.W. Molecular mechanisms of green tea polyphenols with protective effects against skin photoaging. Crit. Rev. Food Sci. Nutr. 2017, 57, 1631–1637. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk-Grochala, J.; Wnuk, M.; Duda, M.; Zuczek, J.; Lewinska, A. Treatment with Modified Extracts of the Microalga Attenuates the Development of Stress-Induced Senescence in Human Skin Cells. Nutrients 2020, 12, 1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Huang, F.; McClements, D.J.; Xie, B.; Sun, Z.; Deng, Q. Oligomeric Procyanidin Nanoliposomes Prevent Melanogenesis and UV Radiation-Induced Skin Epithelial Cell (HFF-1) Damage. Molecules 2020, 25, 1458. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Lee, W.; Jayawardena, T.U.; Cha, S.-H.; Jeon, Y.-J. Dieckol, an algae-derived phenolic compound, suppresses airborne particulate matter-induced skin aging by inhibiting the expressions of pro-inflammatory cytokines and matrix metalloproteinases through regulating NF-κB, AP-1, and MAPKs signaling pathways. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2020, 146, 111823. [Google Scholar] [CrossRef]
- Lee, E.S.; Lee, E.Y.; Yoon, J.; Hong, A.; Nam, S.J.; Ko, J. Sarmentosamide, an Anti-Aging Compound from a Marine-Derived Streptomyces sp. APmarine042. Mar. Drugs 2020, 18, 463. [Google Scholar] [CrossRef] [PubMed]
- Lorz, L.R.; Yoo, B.C.; Kim, M.Y.; Cho, J.Y. Anti-Wrinkling and Anti-Melanogenic Effect of Pradosia mutisii Methanol Extract. Int. J. Mol. Sci. 2019, 20, 1043. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.Y.; Jeong, D.; Park, S.H.; Shin, K.K.; Hong, Y.H.; Kim, E.; Yu, Y.G.; Kim, T.R.; Kim, H.; Lee, J.; et al. Antiwrinkle and Antimelanogenesis Effects of Tyndallized Lactobacillus acidophilus KCCM12625P. Int. J. Mol. Sci. 2020, 21, 1620. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.C.; Lien, H.M.; Ke, H.J.; Chang, L.L.; Chen, C.C.; Chang, T.M. Antioxidative characteristics of Anisomeles indica extract and inhibitory effect of ovatodiolide on melanogenesis. Int. J. Mol. Sci. 2012, 13, 6220–6235. [Google Scholar] [CrossRef] [Green Version]
- Bourhim, T.; Villareal, M.O.; Gadhi, C.; Isoda, H. Elucidation of Melanogenesis-Associated Signaling Pathways Regulated by Argan Press Cake in B16 Melanoma Cells. Nutrients 2021, 13, 2697. [Google Scholar] [CrossRef]
- Yamakoshi, J.; Otsuka, F.; Sano, A.; Tokutake, S.; Saito, M.; Kikuchi, M.; Kubota, Y. Lightening effect on ultraviolet-induced pigmentation of guinea pig skin by oral administration of a proanthocyanidin-rich extract from grape seeds. Pigment Cell Res. 2003, 16, 629–638. [Google Scholar] [CrossRef]
- Pucci, M.; Pasquariello, N.; Battista, N.; Di Tommaso, M.; Rapino, C.; Fezza, F.; Zuccolo, M.; Jourdain, R.; Finazzi Agro, A.; Breton, L.; et al. Endocannabinoids stimulate human melanogenesis via type-1 cannabinoid receptor. J. Biol. Chem. 2012, 287, 15466–15478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.-H.; Min, S.-Y.; Yu, H.-W.; Kim, K.; Kim, S.; Lee, H.-J.; Kim, J.-H.; Park, Y.-J. Effects of Apigenin on RBL-2H3, RAW264.7, and HaCaT Cells: Anti-Allergic, Anti-Inflammatory, and Skin-Protective Activities. Int. J. Mol. Sci. 2020, 21, 4620. [Google Scholar] [CrossRef]
- Luczaj, W.; Domingues, M.D.R.; Domingues, P.; Skrzydlewska, E. Changes in Lipid Profile of Keratinocytes from Rat Skin Exposed to Chronic UVA or UVB Radiation and Topical Application of Cannabidiol. Antioxidants 2020, 9, 1178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, Y.; Lai, R.; Deng, H.; Zhou, F.; Wang, P.; Pang, X.; Huang, G.; Chen, X.; Lin, H.; et al. Protective Effect of the Pearl Extract from Pinctada fucata martensii Dunker on UV-Induced Photoaging in Mice. Chem. Biodivers. 2022, 19, e202100876. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Yoon, S.; Kim, S.; Jung, J.; Kor, M.; Shin, K.; Lim, C.; Han, H.S.; Lee, H.; Park, K.Y.; et al. Anti-Wrinkle Benefits of Peptides Complex Stimulating Skin Basement Membrane Proteins Expression. Int. J. Mol. Sci. 2019, 21, 73. [Google Scholar] [CrossRef] [Green Version]
- Naylor, E.C.; Watson, R.E.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef]
- Biernacki, M.; Brzóska, M.M.; Markowska, A.; Gałażyn-Sidorczuk, M.; Cylwik, B.; Gęgotek, A.; Skrzydlewska, E. Oxidative Stress and Its Consequences in the Blood of Rats Irradiated with UV: Protective Effect of Cannabidiol. Antioxidants 2021, 10, 821. [Google Scholar] [CrossRef]
- Jankovic, A.; Saso, L.; Korac, A.; Korac, B. Relation of Redox and Structural Alterations of Rat Skin in the Function of Chronological Aging. Oxidative Med. Cell. Longev. 2019, 2019, 2471312. [Google Scholar] [CrossRef] [Green Version]
- Mussard, E.; Jousselin, S.; Cesaro, A.; Legrain, B.; Lespessailles, E.; Esteve, E.; Berteina-Raboin, S.; Toumi, H. Andrographis Paniculata and Its Bioactive Diterpenoids Protect Dermal Fibroblasts Against Inflammation and Oxidative Stress. Antioxidants 2020, 9, 432. [Google Scholar] [CrossRef]
- Li, F.M.; Pirzgalska, R.M.; Marques-da-Silva, D.; Ledent, C.; Kofalvi, A.; Carvalho, E. Diabetes and Cannabinoid CB1 receptor deficiency promote similar early onset aging-like changes in the skin. Exp. Gerontol. 2021, 154, 111528. [Google Scholar] [CrossRef]
- Hsu, J.Y.; Lin, H.H.; Li, T.S.; Tseng, C.Y.; Wong, Y.; Chen, J.H. Anti-Melanogenesis Effects of Lotus Seedpod In Vitro and In Vivo. Nutrients 2020, 12, 3535. [Google Scholar] [CrossRef] [PubMed]
- Oh, T.I.; Jung, H.J.; Lee, Y.M.; Lee, S.; Kim, G.H.; Kan, S.Y.; Kang, H.; Oh, T.; Ko, H.M.; Kwak, K.C.; et al. Zerumbone, a Tropical Ginger Sesquiterpene of Zingiber officinale Roscoe, Attenuates alpha-MSH-Induced Melanogenesis in B16F10 Cells. Int. J. Mol. Sci. 2018, 19, 3149. [Google Scholar] [CrossRef] [Green Version]
- Jeong, D.; Lee, J.; Park, S.H.; Kim, Y.A.; Park, B.J.; Oh, J.; Sung, G.-H.; Aravinthan, A.; Kim, J.-H.; Kang, H.; et al. Antiphotoaging and Antimelanogenic Effects of Pursh Ethanol Extract due to Antioxidant- and Autophagy-Inducing Properties. Oxidative Med. Cell. Longev. 2019, 2019, 9679731. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.-S.; Oh, Y.N.; Hyun, S.K.; Kwon, H.J.; Kim, B.W. Betulinic acid isolated from Vitis amurensis root inhibits 3-isobutyl-1-methylxanthine induced melanogenesis via the regulation of MEK/ERK and PI3K/Akt pathways in B16F10 cells. Food Chem. Toxicol. 2014, 68, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhao, B.; Liu, Y.; Wang, R.; Yang, Y.; Yang, L.; Dong, C. MITF-M regulates melanogenesis in mouse melanocytes. J. Dermatol. Sci. 2018, 90, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Li, P.H.; Chiu, Y.P.; Shih, C.C.; Wen, Z.H.; Ibeto, L.K.; Huang, S.H.; Chiu, C.C.; Ma, D.L.; Leung, C.H.; Chang, Y.N.; et al. Biofunctional Activities of Equisetum ramosissimum Extract: Protective Effects against Oxidation, Melanoma, and Melanogenesis. Oxidative Med. Cell. Longev. 2016, 2016, 2853543. [Google Scholar] [CrossRef] [Green Version]
- Pisanti, S.; Malfitano, A.M.; Ciaglia, E.; Lamberti, A.; Ranieri, R.; Cuomo, G.; Abate, M.; Faggiana, G.; Proto, M.C.; Fiore, D.; et al. Cannabidiol: State of the art and new challenges for therapeutic applications. Pharmacol. Ther. 2017, 175, 133–150. [Google Scholar] [CrossRef]
- Sunda, F.; Arowolo, A. A molecular basis for the anti-inflammatory and anti-fibrosis properties of cannabidiol. FASEB J. 2020, 34, 14083–14092. [Google Scholar] [CrossRef]
- Jarocka-Karpowicz, I.; Biernacki, M.; Wronski, A.; Gegotek, A.; Skrzydlewska, E. Cannabidiol Effects on Phospholipid Metabolism in Keratinocytes from Patients with Psoriasis Vulgaris. Biomolecules 2020, 10, 367. [Google Scholar] [CrossRef] [Green Version]
- Gegotek, A.; Atalay, S.; Domingues, P.; Skrzydlewska, E. The Differences in the Proteome Profile of Cannabidiol-Treated Skin Fibroblasts following UVA or UVB Irradiation in 2D and 3D Cell Cultures. Cells 2019, 8, 995. [Google Scholar] [CrossRef]
- Kennedy, P.J. Anti-Aging and Skin Tone Lightening Compositions and Methods For Same. U.S. Patent No 11,337,905, 2019. [Google Scholar]
- Grimaldi, C.; Capasso, A. The endocannabinoid system in the cancer therapy: An overview. Curr. Med. Chem. 2011, 18, 1575–1583. [Google Scholar] [CrossRef]
- Baswan, S.M.; Klosner, A.E.; Glynn, K.; Rajgopal, A.; Malik, K.; Yim, S.; Stern, N. Therapeutic Potential of Cannabidiol (CBD) for Skin Health and Disorders. Clin. Cosmet. Investig. Dermatol. 2020, 13, 927–942. [Google Scholar] [CrossRef]
- Pertwee, R.G. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: Delta9-tetrahydrocannabinol, cannabidiol and delta9-tetrahydrocannabivarin. Br. J. Pharm. 2008, 153, 199–215. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Rossi, E.; Saubamea, B.; Chasseigneaux, S.; Cochois, V.; Choublier, N.; Smirnova, M.; Glacial, F.; Perrière, N.; Bourdoulous, S.; et al. Cannabidiol Increases Proliferation, Migration, Tubulogenesis, and Integrity of Human Brain Endothelial Cells through TRPV2 Activation. Mol. Pharm. 2019, 16, 1312–1326. [Google Scholar] [CrossRef]
- Hua, T.; Vemuri, K.; Pu, M.; Qu, L.; Han, G.W.; Wu, Y.; Zhao, S.; Shui, W.; Li, S.; Korde, A.; et al. Crystal Structure of the Human Cannabinoid Receptor CB. Cell 2016, 167, 750–762.e14. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Chen, X.; Qiang, S.; Su, J.; Li, J. Anti-Oxidation and Anti-Inflammatory Potency Evaluation of Ferulic Acid Derivatives Obtained through Virtual Screening. Int. J. Mol. Sci. 2021, 22, 11305. [Google Scholar] [CrossRef]
- Han, W.; Shi, Y.; Su, J.; Zhao, Z.; Wang, X.; Li, J.; Liu, H. Virtual Screening and Bioactivity Evaluation of Novel Androgen Receptor Antagonists From Anti-PCa Traditional Chinese Medicine Prescriptions. Front. Chem. 2020, 8, 582861. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Choi, Y.-J.; Rohmah, Z.; Jeong, S.-B.; Hwang, D.-J.; Choi, B.-D. Antioxidant Activities in Processed Cockle(Tegillarca granosa) from the Yeosu. J. Agric. Life Sci. 2017, 51, 131–138. [Google Scholar] [CrossRef]
- Garcin, G.; Le Gallic, L.; Stoebner, P.-E.; Guezennec, A.; Guesnet, J.; Lavabre-Bertrand, T.; Martinez, J.; Meunier, L. Constitutive expression of MC1R in HaCaT keratinocytes inhibits basal and UVB-induced TNF-alpha production. Photochem. Photobiol. 2009, 85, 1440–1450. [Google Scholar] [CrossRef] [PubMed]
- Voravuthikunchai, S.P.; Kanchanapoom, T.; Sawangjaroen, N.; Hutadilok-Towatana, N. Antioxidant, antibacterial and antigiardial activities of Walsura robusta Roxb. Nat. Prod. Res. 2010, 24, 813–824. [Google Scholar] [CrossRef]
- Syed Haneef, S.A.; Ranganathan, S. Structural bioinformatics analysis of variants on GPCR function. Curr. Opin. Struct. Biol. 2019, 55, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.L.; Devi, L.A. Diversity of molecular targets and signaling pathways for CBD. Pharmacol. Res. Perspect. 2020, 8, e00682. [Google Scholar] [CrossRef] [PubMed]
- Burstein, S. Cannabidiol (CBD) and its analogs: A review of their effects on inflammation. Bioorganic. Med. Chem. 2015, 23, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Dos-Santos-Pereira, M.; Guimaraes, F.S.; Del-Bel, E.; Raisman-Vozari, R.; Michel, P.P. Cannabidiol prevents LPS-induced microglial inflammation by inhibiting ROS/NF-kappaB-dependent signaling and glucose consumption. Glia 2020, 68, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Ko, J.H.; Kang, H.K.; Kim, S.; Kang, C.I.; Lee, J.N.; Park, S.M.; Hyun, C.G. Antimelanogenic Effects of Polygonum tinctorium Flower Extract from Traditional Jeju Fermentation via Upregulation of Extracellular Signal-Regulated Kinase and Protein Kinase B Activation. Int. J. Mol. Sci. 2018, 19, 2895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, X.; Yang, Y.; Quan, Q.; Huo, T.; Yang, S.; Ju, R.; An, Q. Comparison of the in vitro Anti-Inflammatory Effect of Cannabidiol to Dexamethasone. Clin. Cosmet. Investig. Dermatol. 2022, 15, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Juknat, A.; Pietr, M.; Kozela, E.; Rimmerman, N.; Levy, R.; Gao, F.; Coppola, G.; Geschwind, D.; Vogel, Z. Microarray and pathway analysis reveal distinct mechanisms underlying cannabinoid-mediated modulation of LPS-induced activation of BV-2 microglial cells. PloS ONE 2013, 8, e61462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X. The Effects and Mechanism of Cannabidol on Photoaging Dermal Fibroblasts. Master’s Thesis, Kunming Medical University, Kunming, China, 2021. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Database | Structure | Docking Score |
|---|---|---|---|
| CBD | InterBioScreen |  | −10.274 |
| S-88614 | InterBioScreen |  | −8.494 |
| S-88745 | InterBioScreen |  | −9.267 |
| S-92151 | InterBioScreen |  | −7.285 |
| S-92153 | InterBioScreen |  | −8.219 |
| No. | Stereo View of the Docking Results | Floor Plan View of the Docking Results |
|---|---|---|
| CBD |  |  |
| S-88614 |  |  |
| S-88745 |  |  |
| S-92151 |  |  |
| S-92153 |  |  |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Su, J.; Wang, R.; Hao, R.; Fu, C.; Chen, J.; Li, J.; Wang, X. Structural Optimization of Cannabidiol as Multifunctional Cosmetic Raw Materials. Antioxidants 2023, 12, 314. https://doi.org/10.3390/antiox12020314
Chen X, Su J, Wang R, Hao R, Fu C, Chen J, Li J, Wang X. Structural Optimization of Cannabidiol as Multifunctional Cosmetic Raw Materials. Antioxidants. 2023; 12(2):314. https://doi.org/10.3390/antiox12020314
Chicago/Turabian StyleChen, Xuelian, Jie Su, Runan Wang, Rui Hao, Chenggong Fu, Jingjing Chen, Jiazhong Li, and Xin Wang. 2023. "Structural Optimization of Cannabidiol as Multifunctional Cosmetic Raw Materials" Antioxidants 12, no. 2: 314. https://doi.org/10.3390/antiox12020314
APA StyleChen, X., Su, J., Wang, R., Hao, R., Fu, C., Chen, J., Li, J., & Wang, X. (2023). Structural Optimization of Cannabidiol as Multifunctional Cosmetic Raw Materials. Antioxidants, 12(2), 314. https://doi.org/10.3390/antiox12020314