
CHIR99021 Maintenance of the Cell Stemness by Regulating Cellular Iron Metabolism



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Preparation of MEF Feeder Cells and Culture of mESCs
2.3. Drug Treatment
2.4. Western Blot Analysis
2.5. Total RNA Extraction and qRT-PCR
2.6. RNA-Sequencing (RNA-Seq) and Data Analysis
2.7. Inductively Coupled Plasma Mass Spectrometry (ICP-MS)
2.8. Alkaline Phosphatase Staining
2.9. Electric Cell-Substrate Impedance Sensing (ECIS) Detection of Cell Proliferation
2.10. Profiles of Mitochondrial Respiration and Glycolysis
2.11. Statistical Analysis
3. Results
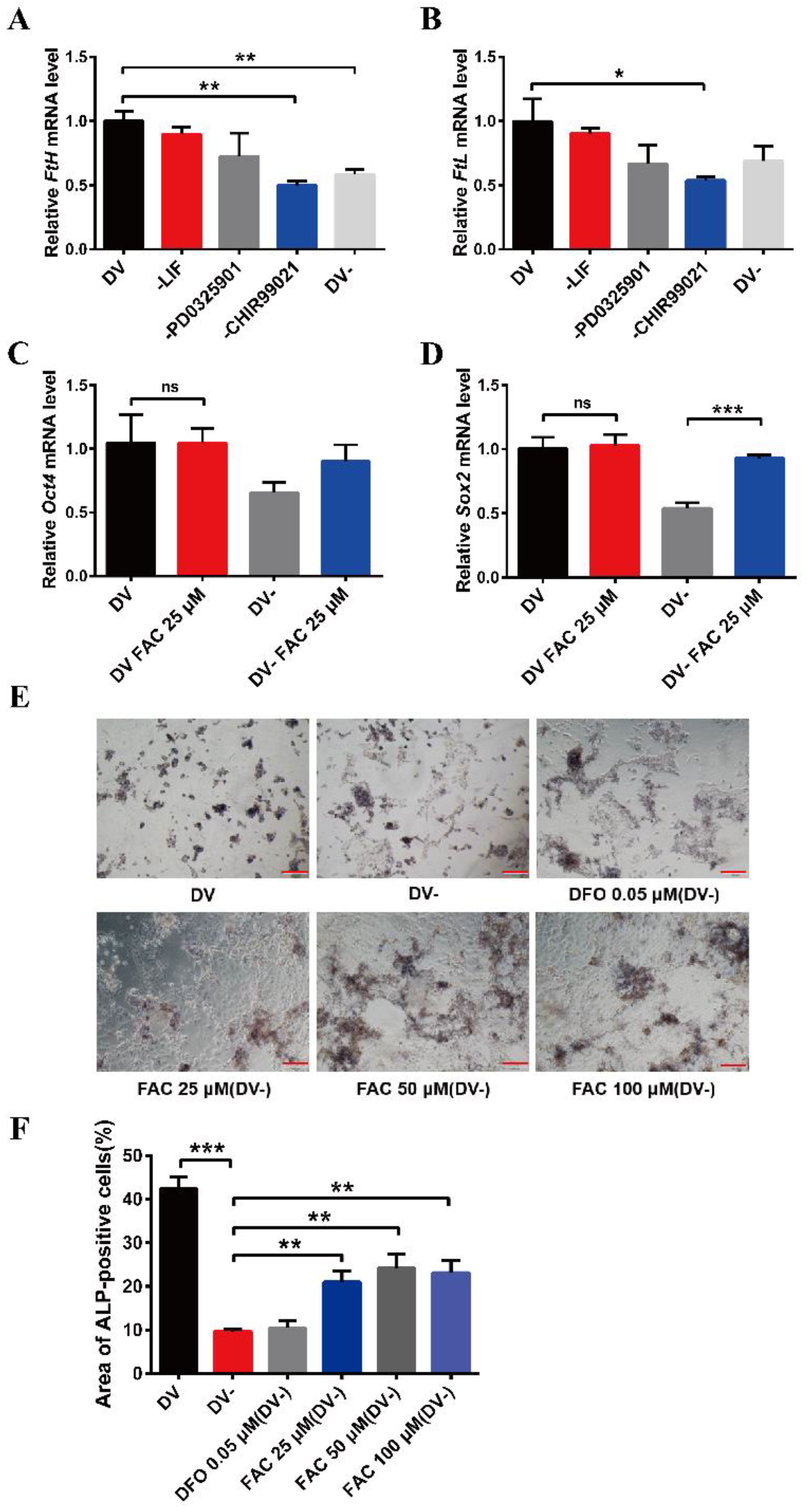
3.1. The Effects of CHIR99021 and Iron on Cell Stemness in mESCs
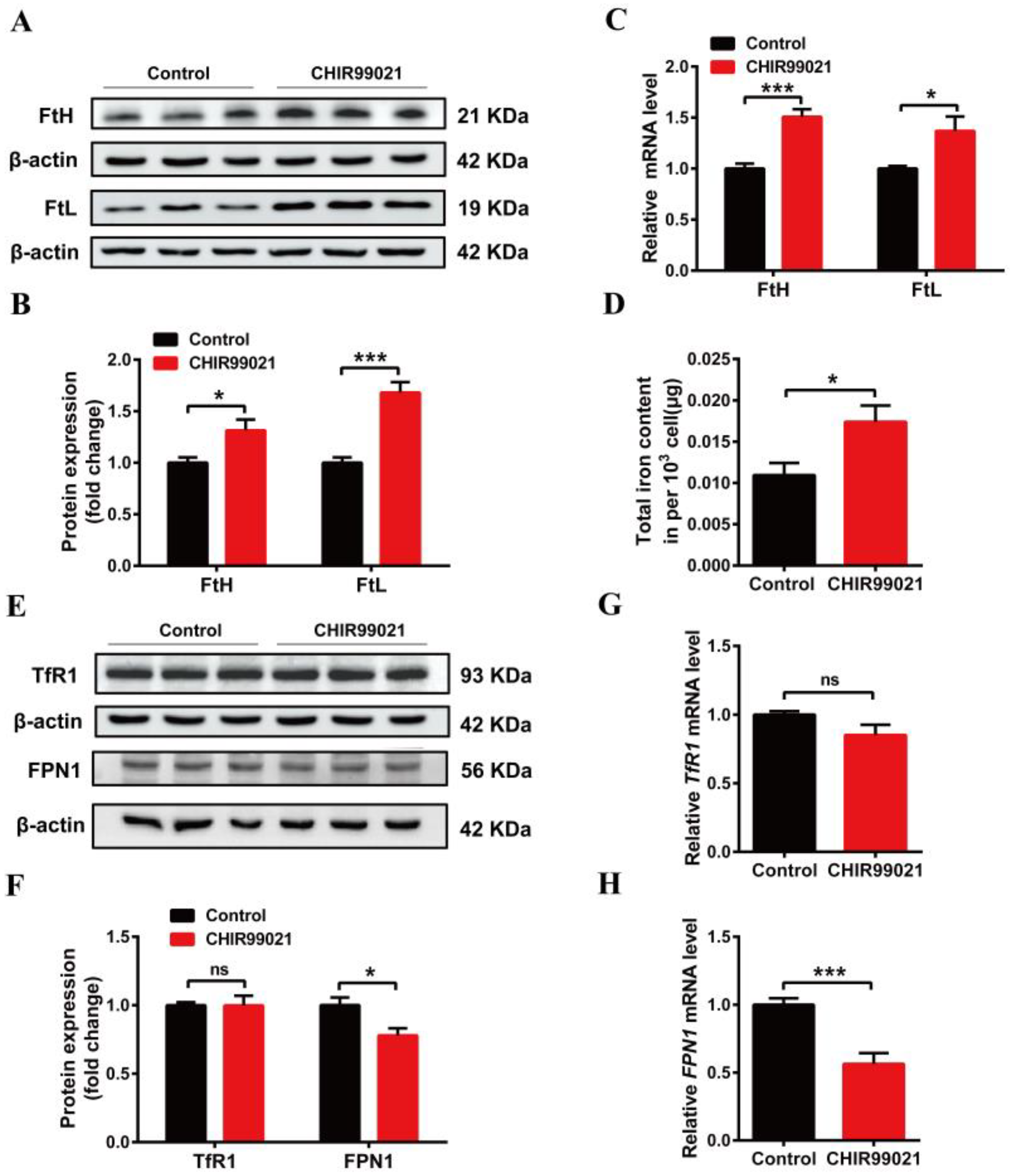
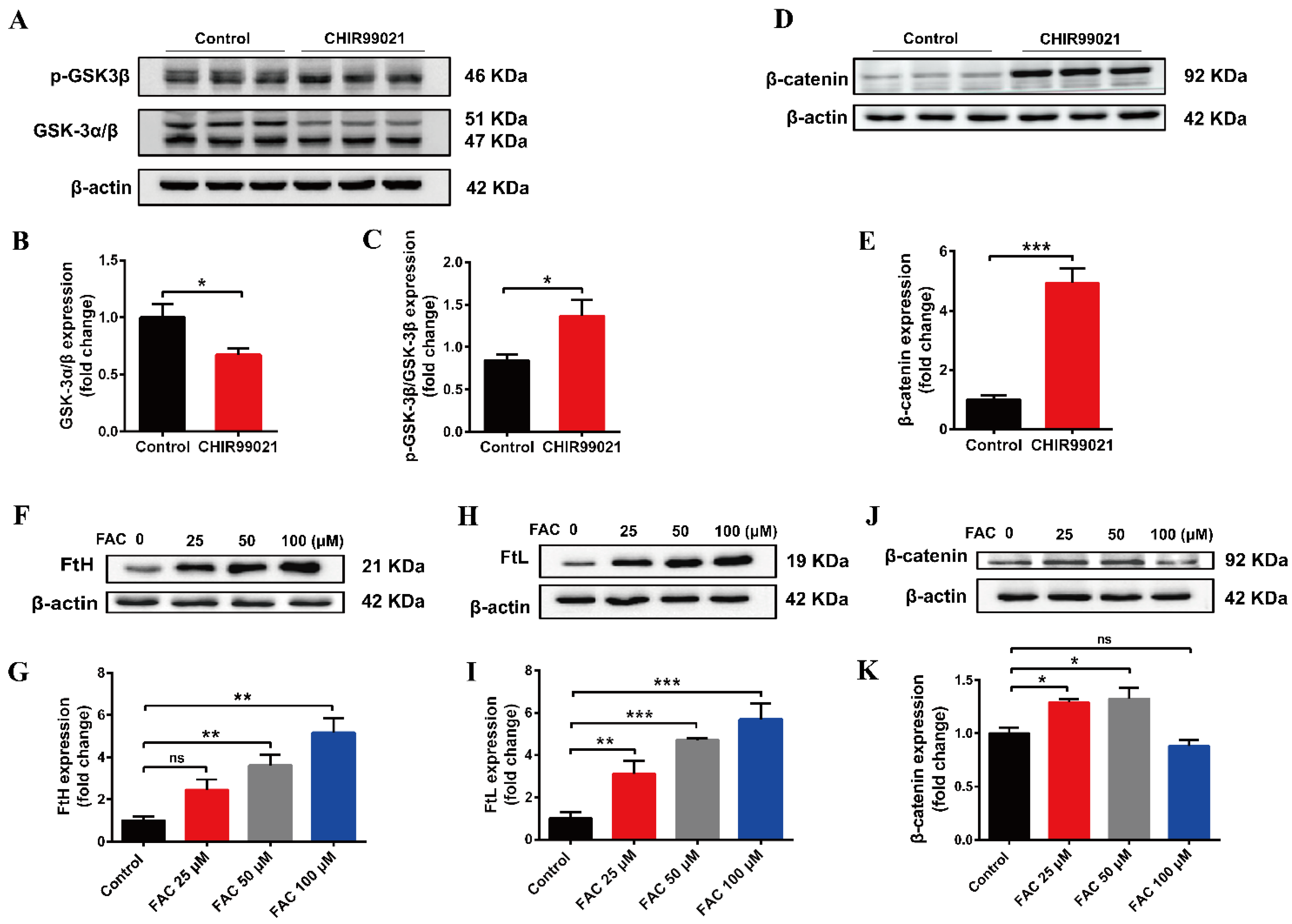
3.2. CHIR99021 Increasing the Expression of Ferritins, Cellular Iron Levels and Inhibiting the Expression of FPN1 in Neuro-2a Cells
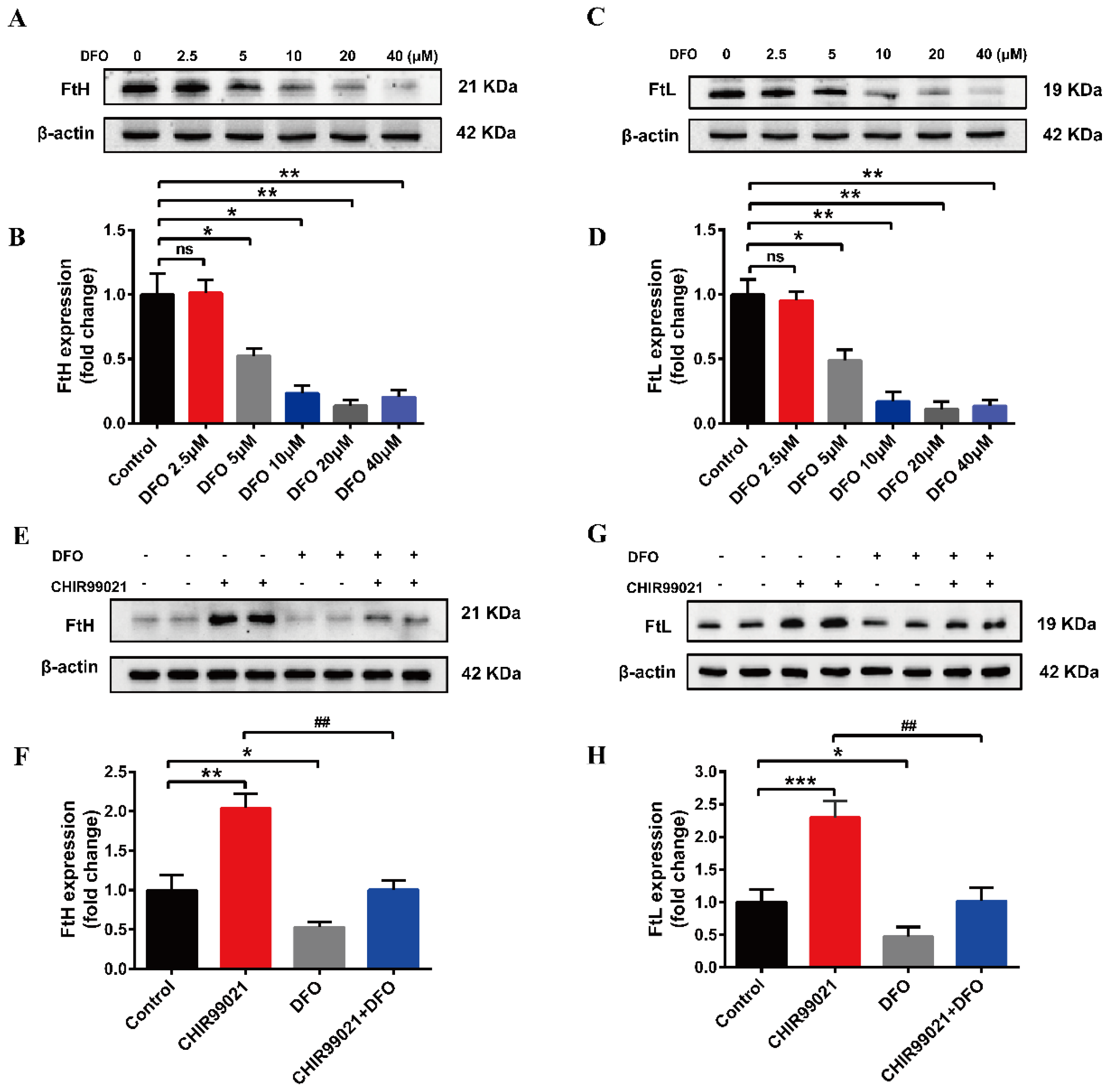
3.3. DFO Abrogates the Increase of Ferritin Induced by CHIR99021
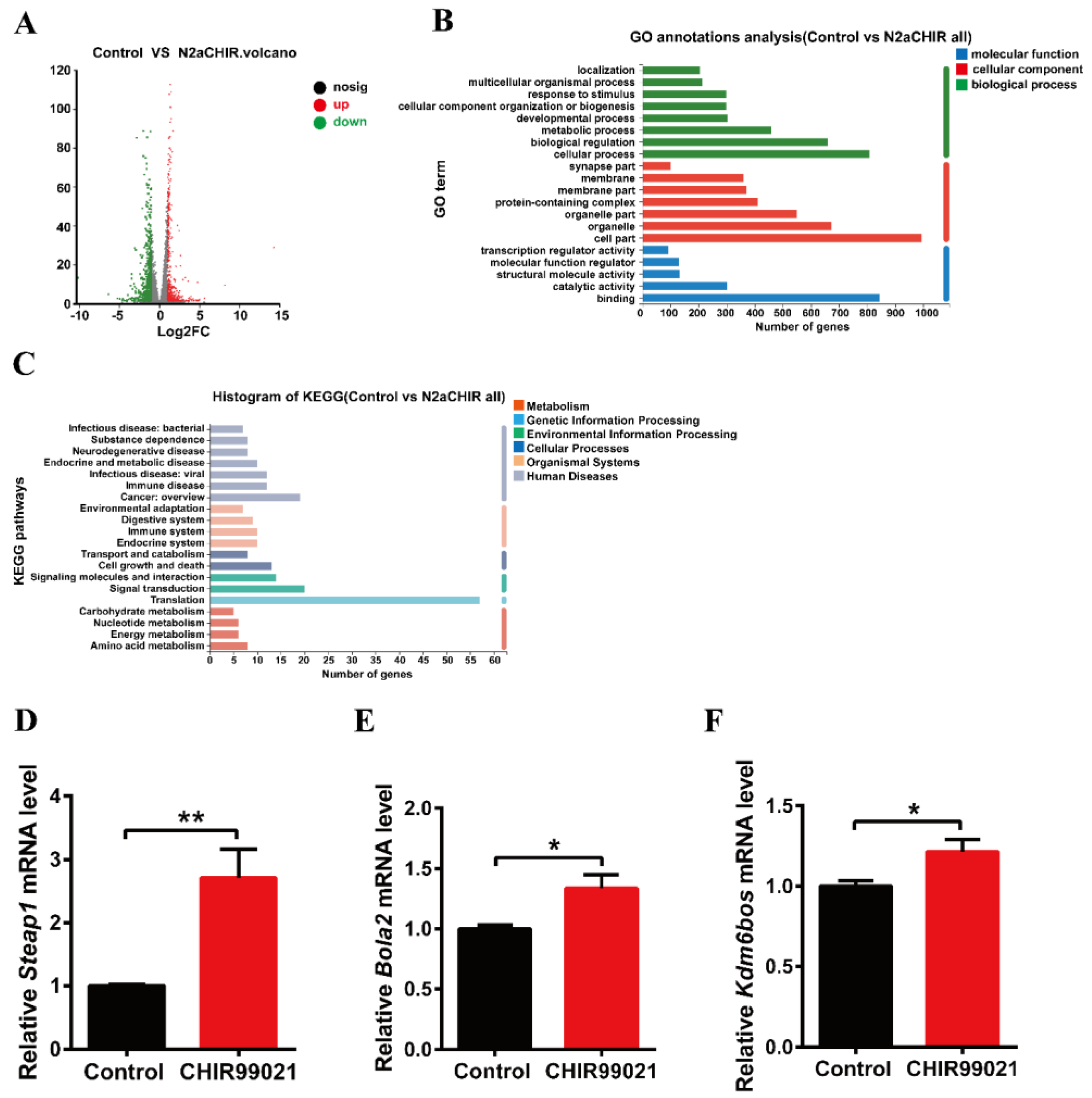
3.4. CHIR99021 Stimulates the Expression of Steap1, Bola2 and Kdm6bos Increasing in Neuro-2a Cells
3.5. Both CHIR99021 and Iron Promote the Expression of β-Catenin
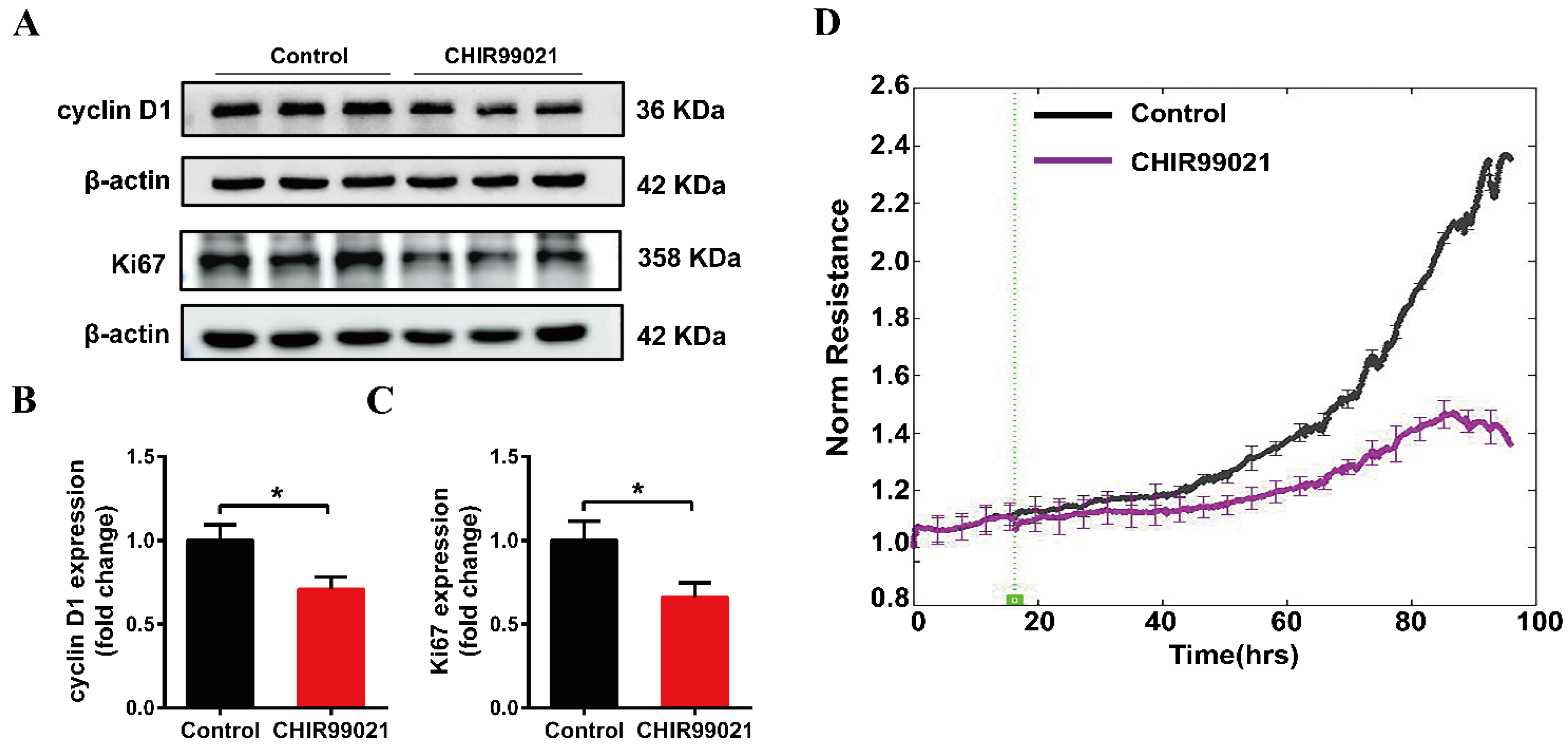
3.6. CHIR99021 Affects the Growth of Neuro-2a Cells
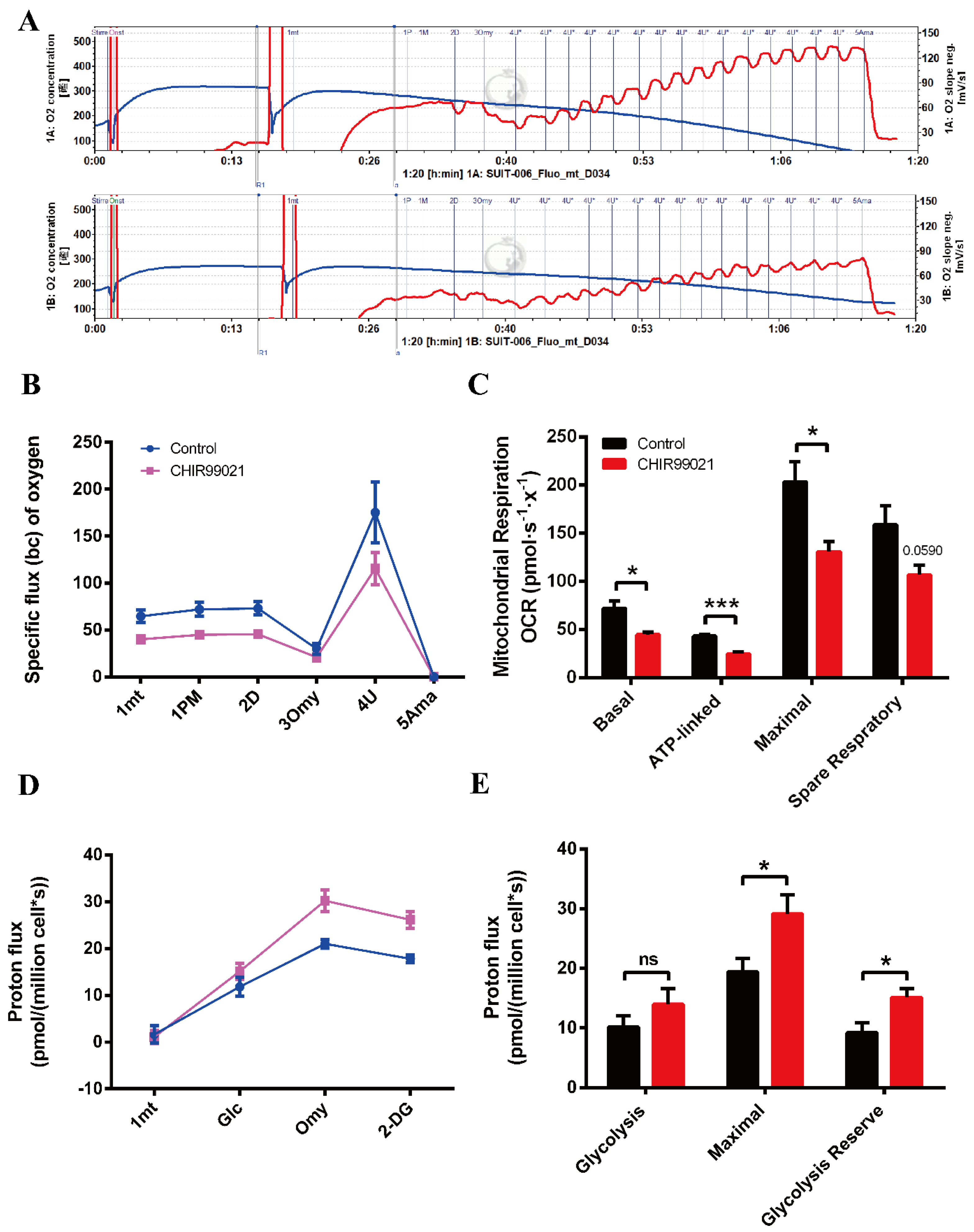
3.7. CHIR99021 Treatment Decreases Mitochondrial Oxidative Phosphorylation and Enhances Glycolytic Capacity in Neuro-2a Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, M.J.; Kaufman, M.H. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA 1981, 78, 7634–7638. [Google Scholar] [CrossRef] [PubMed]
- Chambers, I.; Smith, A. Self-renewal of teratocarcinoma and embryonic stem cells. Oncogene 2004, 23, 7150–7160. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Yan, R.; Zhang, X.; Wang, L.; Ke, S.; Qu, Y. Activating Wnt/beta-catenin signaling pathway for disease therapy: Challenges and opportunities. Pharmacol. Ther. 2019, 196, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Corda, G.; Sala, A. Non-canonical WNT/PCP signalling in cancer: Fzd6 takes centre stage. Oncogenesis 2017, 6, e364. [Google Scholar] [CrossRef]
- Lang CM, R.; Chan, C.K.; Veltri, A.; Lien, W.H. Wnt Signaling Pathways in Keratinocyte Carcinomas. Cancers 2019, 11, 1216. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Wu, Y.; Awadasseid, A.; Tanaka, Y.; Zhang, W. New Advances in Canonical Wnt/beta-Catenin Signaling in Cancer. Cancer Manag. Res. 2020, 12, 6987–6998. [Google Scholar] [CrossRef]
- Wang, F.; Liu, L.; Gao, W.Q. Activation of Wnt/ß-catenin signaling enhanced stemness of prostate cancer cells. Tumor 2019, 39, 525–533+567. [Google Scholar]
- Tang, Q.; Chen, J.; Di, Z.; Yuan, W.; Zhou, Z.; Liu, Z.; Han, S.; Liu, Y.; Ying, G.; Shu, X.; et al. TM4SF1 promotes EMT and cancer stemness via the Wnt/beta-catenin/SOX2 pathway in colorectal cancer. J. Exp. Clin. Cancer Res. 2020, 39, 232. [Google Scholar] [CrossRef]
- Niu, Y.M.; Yang, T.L. A number of important biochemical function of trace metal elements and its relationship to human health. Stud. Trace Elem. Health 2014, 31, 78–80. [Google Scholar]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschella, C. Two to tango: Regulation of Mammalian iron metabolism. Cell 2010, 142, 24–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantopoulos, K.; Porwal, S.K.; Tartakoff, A.; Devireddy, L. Mechanisms of mammalian iron homeostasis. Biochemistry 2012, 51, 5705–5724. [Google Scholar] [CrossRef] [PubMed]
- Philpott, C.C. Coming into view: Eukaryotic iron chaperones and intracellular iron delivery. J. Biol. Chem. 2012, 287, 13518–13523. [Google Scholar] [CrossRef]
- Zhang, D.L.; Ghosh, M.C.; Rouault, T.A. The physiological functions of iron regulatory proteins in iron homeostasis—An update. Front. Pharmacol. 2014, 5, 124. [Google Scholar] [CrossRef]
- Zhu, J.; Wu, X.Y. Modulating factors in the differentiation of stem cells: Present research and progress. J. Clin. Rehabil. Tissue Eng. Res. 2009, 13, 4. [Google Scholar]
- Lein, P.; Gallagher, P.J.; Amodeo, J.; Howie, H.; Roth, J.A. Manganese induces neurite outgrowth in PC12 cells via upregulation of alpha(v) integrins. Brain Res. 2000, 885, 220–230. [Google Scholar] [CrossRef]
- Hong, J.-H.; Noh, K.-M.; Yoo, Y.-E.; Choi, S.-Y.; Park, S.-Y.; Kim, Y.-H.; Chung, J.-M. Iron promotes the survival and neurite extension of serum-starved PC12 cells in the presence of NGF by enhancing cell attachment. Mol. Cells 2003, 15, 10–19. [Google Scholar] [PubMed]
- Rostami, A.A.; Kouchesfahani, H.M.; Kiani, S.; Fakheri, R. Iron Oxide Nanoparticles Reduced Retinoic Acid Induced-neuronal Differentiation of Mouse Embryonic Stem Cells By ROS Generation. Arch. Iran. Med. 2015, 18, 586–590. [Google Scholar]
- Zhou, Y.; Zhang, H.; Liao, S.; Hu, F.Q.; Yi, J.; Liu, Y.B.; Jin, J.D. Immunomodulatory effects of deferoxamine and interferon gamma on human dental pulp stem cells. Chin. J. Tissue Eng. Res. 2022, 26, 8. [Google Scholar]
- Hu, K.; Olsen, B.R. The roles of vascular endothelial growth factor in bone repair and regeneration. Bone 2016, 91, 30–38. [Google Scholar] [CrossRef]
- Pricola, K.L.; Kuhn, N.Z.; Haleem-Smith, H.; Song, Y.; Tuan, R.S. Interleukin-6 maintains bone marrow-derived mesenchymal stem cell stemness by an ERK1/2-dependent mechanism. J. Cell. Biochem. 2009, 108, 577–588. [Google Scholar] [CrossRef]
- Mehta, K.J. Role of iron and iron-related proteins in mesenchymal stem cells: Cellular and clinical aspects. J. Cell. Physiol. 2021, 236, 7266–7289. [Google Scholar] [CrossRef]
- Samara, A.; Shapira, S.; Lubin, I.; Shpilberg, O.; Avigad, S.; Granot, G.; Raanani, P. Deferasirox induces cyclin D1 degradation and apoptosis in mantle cell lymphoma in a reactive oxygen species- and GSK3beta-dependent mechanism. Br. J. Haematol. 2021, 192, 747–760. [Google Scholar] [CrossRef]
- Zhu, J.L.; Lu, Z.S.; Wu, Y.; Xu, Y.J.; Yao, J.; Wang, X.X.; Wang, D.C. Copper, iron, zinc and aluminum induce the apoptosis of SH-SY5Y cells through Akt/GSK-3β signaling pathways. Pract. Geriatr. 2017, 31, 723–727. [Google Scholar]
- Guo, C.; Wang, P.; Zhong, M.-L.; Wang, T.; Huang, X.-S.; Li, J.-Y.; Wang, Z.-Y. Deferoxamine inhibits iron induced hippocampal tau phosphorylation in the Alzheimer transgenic mouse brain. Neurochem. Int. 2013, 62, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Fan, Y.; Wu, P.; Huang, Z.; Li, X.; Zhao, L.; Ji, Y.; Zhu, M. Parkinson’s Disease Dementia: Synergistic Effects of Alpha-Synuclein, Tau, Beta-Amyloid, and Iron. Front. Aging Neurosci. 2021, 13, 743754. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Cellular iron: Ferroportin is the only way out. Cell Metab. 2005, 1, 155–157. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, W.; Chen, J.; Liu, S.; Wang, M.; Yang, L.; Chen, C.; Qi, M.; Xu, Y.; Qiao, Z.; et al. Nuclear Exosome Targeting Complex Core Factor Zcchc8 Regulates the Degradation of LINE1 RNA in Early Embryos and Embryonic Stem Cells. Cell Rep. 2019, 29, 2461–2472. [Google Scholar] [CrossRef]
- Ai, Z.; Shao, J.; Wu, Y.; Yu, M.; Du, J.; Shi, X.; Shi, X.; Zhang, Y.; Guo, Z. CHIR99021 enhances Klf4 Expression through beta-Catenin Signaling and miR-7a Regulation in J1 Mouse Embryonic Stem Cells. PLoS ONE 2016, 11, e0150936. [Google Scholar] [CrossRef]
- Kim, K.; Mitra, S.; Wu, G.; Berka, V.; Song, J.; Yu, Y.; Poget, S.; Wang, D.-N.; Tsai, A.-L.; Zhou, M. Six-Transmembrane Epithelial Antigen of Prostate 1 (STEAP1) Has a Single b Heme and Is Capable of Reducing Metal Ion Complexes and Oxygen. Biochemistry 2016, 55, 6673–6684. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.J.; Frey, A.G.; Palenchar, D.J.; Achar, S.; Bullough, K.Z.; Vashisht, A.; Wohlschlegel, J.A.; Philpott, C.C. A PCBP1-BolA2 chaperone complex delivers iron for cytosolic [2Fe-2S] cluster assembly. Nat. Chem. Biol. 2019, 15, 872–881. [Google Scholar] [CrossRef]
- Patel, S.J.; Protchenko, O.; Shakoury-Elizeh, M.; Baratz, Z.; Jadhav, S.; Philpott, C.C. The iron chaperone and nucleic acid–binding activities of poly(rC)-binding protein 1 are separable and independently essential. Proc. Natl. Acad. Sci. USA 2021, 118, e2104666118. [Google Scholar] [CrossRef]
- He, X.J.; Zheng, W.W.; Jia, L.L.; Li, H.; Xu, L.M.; Zeng, J.W.; Xu, H.F.; Zheng, C.S.; Ye, J.X.; Wu, G.W.; et al. Study on the Mechanism of Tiaogu Tablet Regulating Wnt/β-catenin Signaling Pathway to Delay Articular Cartilage Degeneration in Osteoarthritis. Rheum. Arthritis 2019, 8, 5–9. [Google Scholar]
- Bartels, P.L.; Stodola, J.L.; Burgers, P.M.J.; Barton, J.K. A Redox Role for the [4Fe4S] Cluster of Yeast DNA Polymerase delta. J. Am. Chem. Soc. 2017, 139, 18339–18348. [Google Scholar] [CrossRef]
- Aulicino, F.; Pedone, E.; Sottile, F.; Lluis, F.; Marucci, L.; Cosma, M.P. Canonical Wnt Pathway Controls mESC Self-Renewal Through Inhibition of Spontaneous Differentiation via β-Catenin/TCF/LEF Functions. Stem Cell Rep. 2020, 15, 646–661. [Google Scholar] [CrossRef] [PubMed]
- Rafalski, V.A.; Mancini, E.; Brunet, A. Energy metabolism and energy-sensing pathways in mammalian embryonic and adult stem cell fate. J. Cell Sci. 2012, 125 Pt 23, 5597–5608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shyh-Chang, N.; Ng, H.H. The metabolic programming of stem cells. Genes Dev. 2017, 31, 336–346. [Google Scholar] [CrossRef]
- Nagini, S.; Sophia, J.; Mishra, R. Glycogen synthase kinases: Moonlighting proteins with theranostic potential in cancer. Semin. Cancer Biol. 2019, 56, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Su, K.; Sun, X.; Jiang, Y.; Wang, L.; Hu, C.; Zhang, C.; Lu, M.; Du, X.; Xing, B. Sec62 promotes stemness and chemoresistance of human colorectal cancer through activating Wnt/beta-catenin pathway. J. Exp. Clin. Cancer Res. 2021, 40, 132. [Google Scholar] [CrossRef]
- Narusaka, T.; Ohara, T.; Noma, K.; Nishiwaki, N.; Katsura, Y.; Kato, T.; Sato, H.; Tomono, Y.; Kikuchi, S.; Tazawa, H.; et al. Nanog is a promising chemoresistant stemness marker and therapeutic target by iron chelators for esophageal cancer. Int. J. Cancer 2021, 149, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.S.; Moran, E.; Sanchez, M. Cellular Iron Metabolism—The IRP/IRE Regulatory Network. Iron Metab. 2012, 2, 25–58. [Google Scholar]
- Hubert, R.S.; Vivanco, I.; Chen, E.; Rastegar, S.; Leong, K.; Mitchell, S.C.; Madraswala, R.; Zhou, Y.; Kuo, J.; Raitano, A.B.; et al. STEAP: A prostate-specific cell-surface antigen highly expressed in human prostate tumors. Proc. Natl. Acad. Sci. USA 1999, 96, 14523–14528. [Google Scholar] [CrossRef]
- Lagunas-Rangel, F.A. KDM6B (JMJD3) and its dual role in cancer. Biochimie 2021, 184, 63–71. [Google Scholar] [CrossRef]
- Liu, J.; Gao, L.; Zhan, N.; Xu, P.; Yang, J.; Yuan, F.; Xu, Y.; Cai, Q.; Geng, R.; Chen, Q. Hypoxia induced ferritin light chain (FTL) promoted epithelia mesenchymal transition and chemoresistance of glioma. J. Exp. Clin. Cancer Res. 2020, 39, 137. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Singh, P.; Srivastava, A. Iron: Key player in cancer and cell cycle? J. Trace Elem. Med. Biol. 2020, 62, 126582. [Google Scholar] [CrossRef]
- Troadec, M.-B.; Courselaud, B.; Détivaud, L.; Haziza-Pigeon, C.; Leroyer, P.; Brissot, P.; Loréal, O. Iron overload promotes Cyclin D1 expression and alters cell cycle in mouse hepatocytes. J. Hepatol. 2006, 44, 391–399. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Gene Accession Number) | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| TfR1 (NM_001357298.1) | GAGTATCACTTCCTGTCGCCCTATG | GCTGAGAGAGTGTGAGAGCCAGAGC |
| FPN1 (NM_016917.2) | TTCCGCACTTTCCGAGATG | AGTCAAAGCCCAGGACTGTCA |
| FtH (NM_010239.2) | TGCCATCAACCGCCAGATCAAC | TCTTCAGAGCCACATCATCT CGGTC |
| FtL (NM_010240.2) | CAACCATCTGACCAACCTCCGCAG | AAAGAGATACTCGCCCAGAGATCC |
| Oct4 (NM_013633.3) | GAGGAGTCCCAGGACATGAA | AGATGGTGGTCTGGCTGAAC |
| Sox2 (NM_011443.4) | CTGCAGTACAACTCCATGACCAG | GGACTTGACCACAGAGCCCAT |
| Steap1 (NM_027399.3) | GGTCGCCATTACCCTCTTGG | GGTATGAGAGACTGTAAACAGCG |
| Bola2 (NM_175103.3) | GAACTCAGCGCCGATTACCTC | CAGTGGCTTTCCCTCGAACTT |
| Kdm6bos (NM_001017426.2) | AGTGAGGAAGCCGTATGCTG | AGCCCCATAGTTCCGTTTGTG |
| β-actin (NM_007393.5) | AGGCCCAGAGCAAGAGAGGTA | TCTCCATGTCGTCCCAGTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.; He, Y.; Jin, X.; Xie, J.; Yu, P.; Gao, G.; Chang, S.; Zhang, J.; Chang, Y.-Z. CHIR99021 Maintenance of the Cell Stemness by Regulating Cellular Iron Metabolism. Antioxidants 2023, 12, 377. https://doi.org/10.3390/antiox12020377
Han Y, He Y, Jin X, Xie J, Yu P, Gao G, Chang S, Zhang J, Chang Y-Z. CHIR99021 Maintenance of the Cell Stemness by Regulating Cellular Iron Metabolism. Antioxidants. 2023; 12(2):377. https://doi.org/10.3390/antiox12020377
Chicago/Turabian StyleHan, Yingying, Yong He, Xiaofang Jin, Jiayi Xie, Peng Yu, Guofen Gao, Shiyang Chang, Jianhua Zhang, and Yan-Zhong Chang. 2023. "CHIR99021 Maintenance of the Cell Stemness by Regulating Cellular Iron Metabolism" Antioxidants 12, no. 2: 377. https://doi.org/10.3390/antiox12020377
APA StyleHan, Y., He, Y., Jin, X., Xie, J., Yu, P., Gao, G., Chang, S., Zhang, J., & Chang, Y.-Z. (2023). CHIR99021 Maintenance of the Cell Stemness by Regulating Cellular Iron Metabolism. Antioxidants, 12(2), 377. https://doi.org/10.3390/antiox12020377