Liver Injury Induced by Carbon Tetrachloride in Mice Is Prevented by the Antioxidant Capacity of Anji White Tea Polyphenols


Abstract
:1. Introduction
2. Materials and Methods
2.1. AJWTP Extraction
2.2. Determination of Anji White Tea Polyphenol (AJWTP) Content
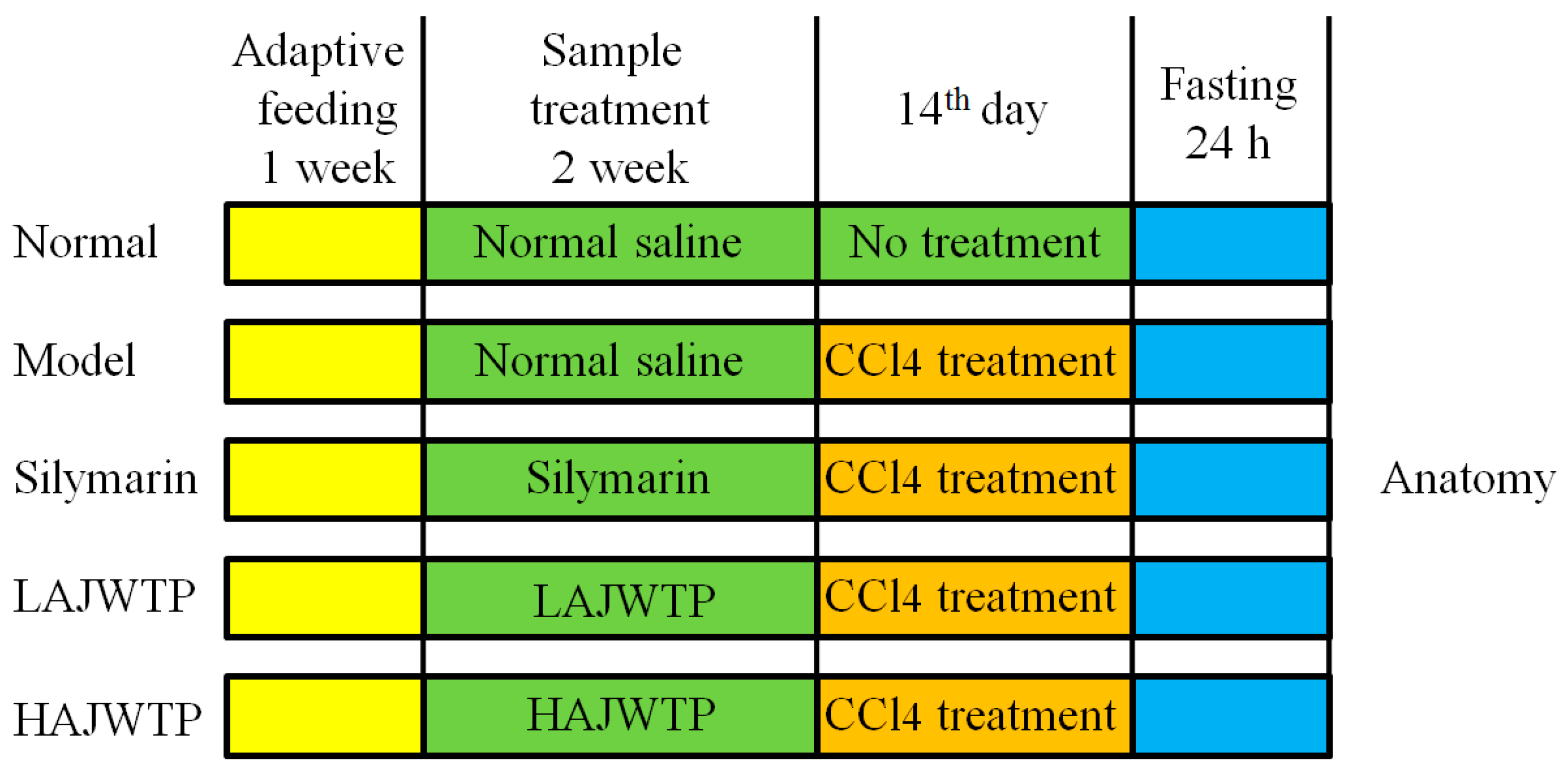
2.3. Animal Experiments
2.4. Pathological Observation of Liver Tissue in Mice
2.5. Determination of Aspartate Aminotransferase (AST), Alanine Aminotransferase (ALT), Triglyceride (TG), and Total Cholesterol (TC) Serum Levels in Mice
2.6. Determination of Malondialdehyde (MDA) Content and SOD and GSH-Px Activities in the Serum and Liver Tissues in Mice
2.7. Quantitative PCR (qPCR) Assay
2.8. Western Blot
2.9. Statistical Analysis
3. Results
3.1. Content of AJWTP
3.2. Liver Index of Mice
3.3. Serum AST, ALT, TG, and TC Levels in Mice
3.4. Serum and Hapatic MDA Content and SOD and GSH-Px Activities in Mice
3.5. Pathological Observation of Hepatic Tissue
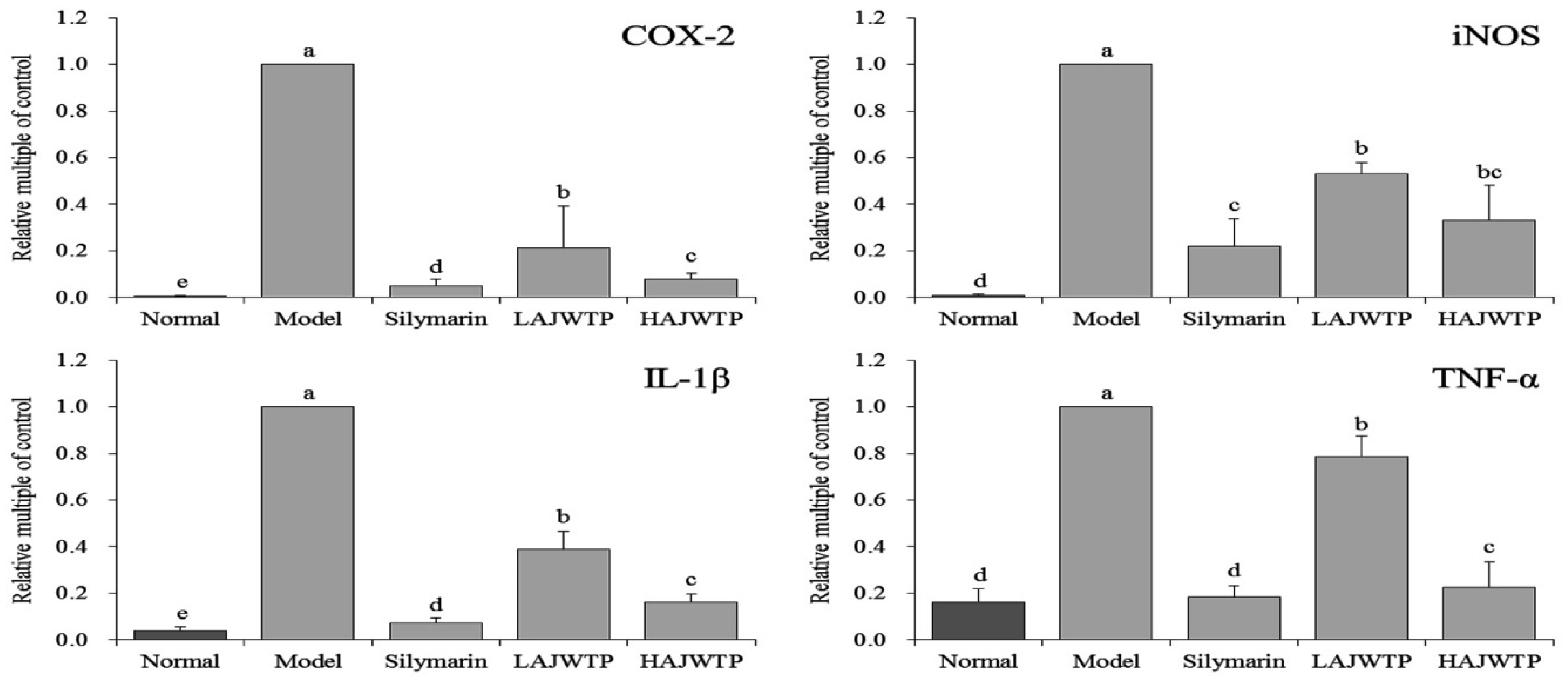
3.6. Cyclooxygenase-2 (COX-2), Inducible Nitric Oxide Synthase (iNOS), Interleukin-1 beta (IL-1β), and Tumor Necrosis Factor alpha (TNF-α) mRNA Expression in the Hepatic Tissue of Mice
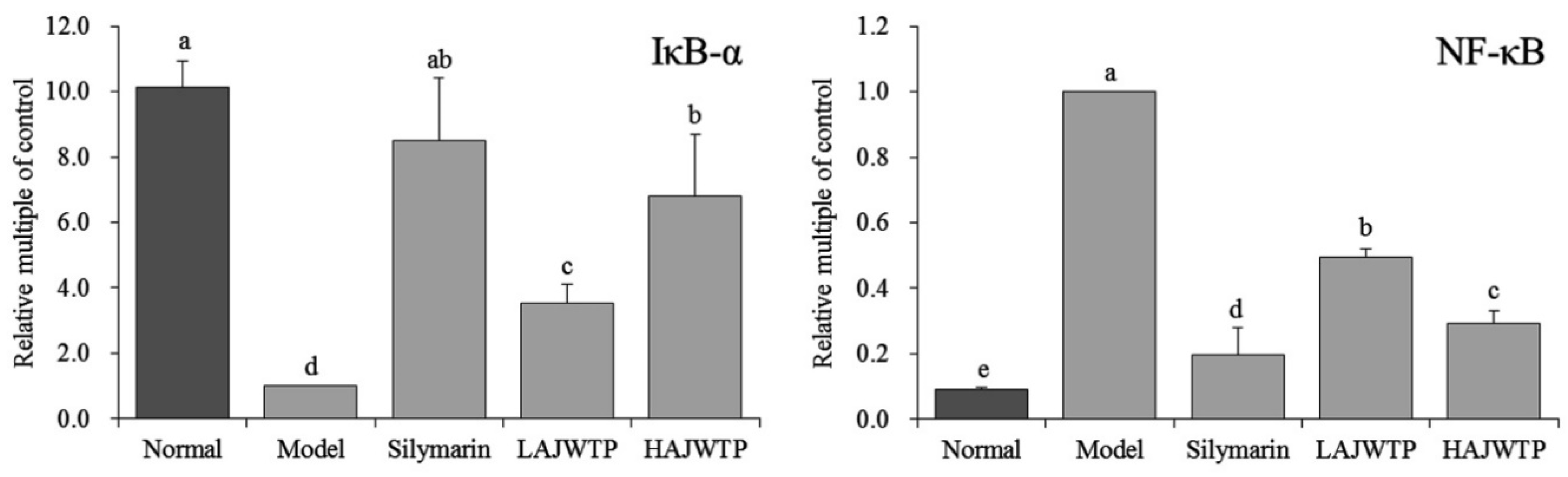
3.7. Nuclear Factor κ-Light-Chain-Enhancer of Activated B Cells (NF-κB) and Nuclear Factor of Kappa Light Polypeptide Gene Enhancer in B-cell Inhibitor Alpha (IκB-α) mRNA Expression in the Hepatic Tissue of Mice
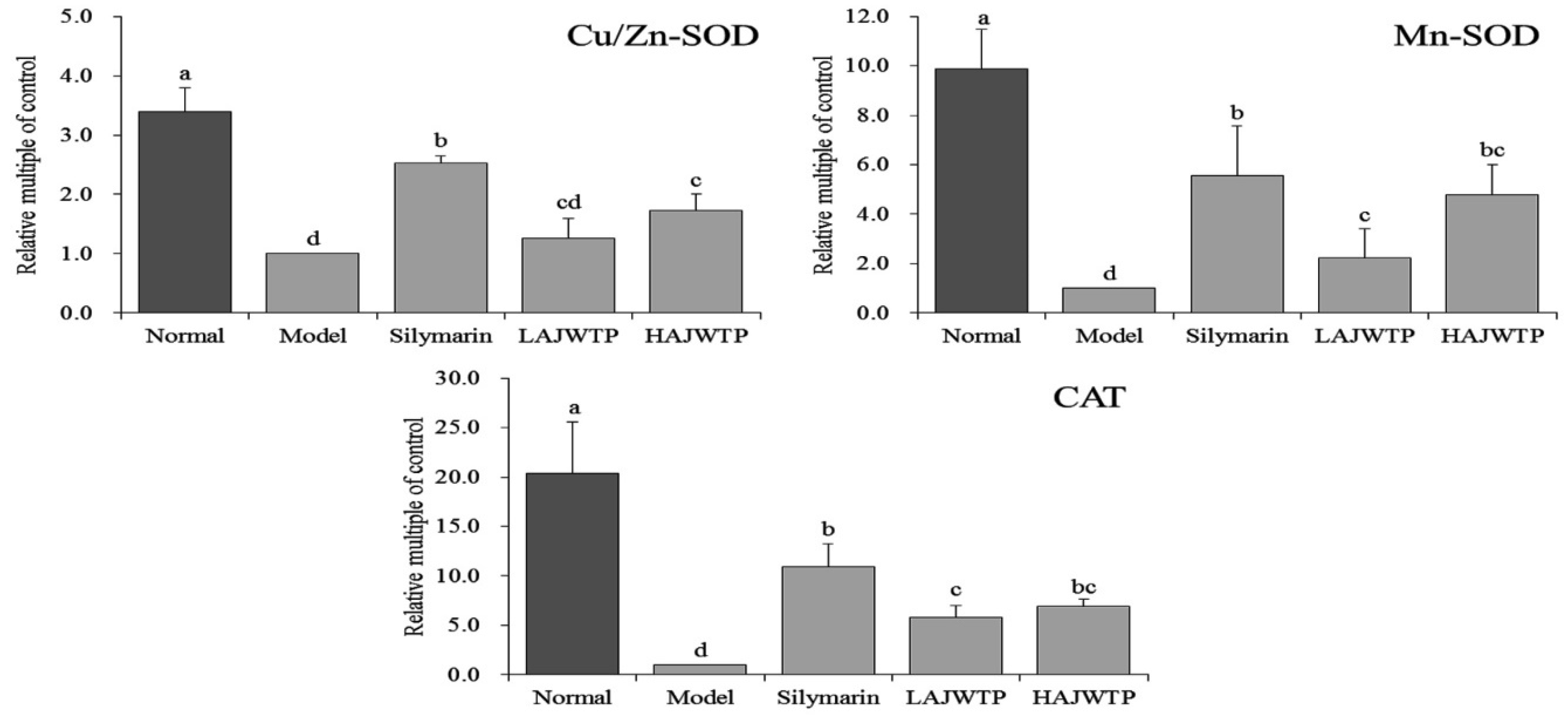
3.8. Cu/Zn-SOD, Mn-SOD, and CAT mRNA Expression in Hepatic Tissue of Mice
3.9. COX-2 and NF-κB Protein Expression in the Hepatic Tissue of Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lee, C.H.; Park, S.W.; Kim, Y.S.; Kang, S.S.; Kim, J.A.; Lee, S.H.; Lee, S.M. Protective mechanism of glycyrrhizin on acute liver injury induced by carbon tetrachloride in mice. Biol. Pharm. Bull. 2007, 30, 1898–1904. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.W.; Boll, M.; Stampfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef] [PubMed]
- Ingawale, D.K.; Mandlik, S.K.; Naik, S.R. Models of hepatotoxicity and the underlying cellular, biochemical and immunological mechanism(s): A critical discussion. Environ. Toxicol. Pharmacol. 2014, 37, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Pramyothin, P.; Janthasoot, W.; Pongnimitprasert, N.; Phrukudom, S.; Ruangrungsi, N. Hepatotoxic effect of (+)usnic acid from Usnea siamensis Wainio in rats, isolated rat hepatocytes and isolated rat liver mitochondria. J. Ethnopharmacol. 2004, 90, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Manibusan, M.K.; Odin, M.; Eastmond, D.A. Postulated carbon tetrachloride mode of action: A review. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2007, 25, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Lettéron, P.; Labbe, G.; Degott, C.; Berson, A.; Fromenty, B.; Delaforge, M.; Larrey, D.; Pessayre, D. Mechanism for the protective effects of silymarin against carbon tetrachloride-induced lipid peroxidation and hepatotoxicity in mice. Evidence that silymarin acts both as an inhibitor of metabolic activation and as a chain-breaking antioxidant. Biochem. Pharmacol. 1990, 39, 2027–2034. [Google Scholar] [CrossRef]
- Li, H.Y.; Xia, J.Q.; Yang, L.Y.; Zhong, R.Z. Plant polyphenols: Antioxidant capacity and application in animal production. China J. Anim. Nutr. 2013, 25, 2529–2534. [Google Scholar]
- Carluccio, M.A.; Siculella, L.; Ancora, M.A.; Massaro, M.; Scoditti, E.; Storelli, C.; Visioli, F.; Distante, A.; de Caterina, R. Olive oil and red wine antioxidant polyphenols inhibit endothelial activation: Antiatherogenic properties of Mediterranean diet phytochemicals. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Rana, S.; Patial, V.; Gupta, M.; Bhushan, S.; Padwad, Y.S. Antioxidant and hepatoprotective effect of polyphenols from apple pomace extract via apoptosis inhibition and Nrf2 activation in mice. Hum. Exp. Toxicol. 2016, 35, 1264–1275. [Google Scholar] [CrossRef]
- Delwing-Dal Magro, D.; Roecker, R.; Junges, G.M.; Rodrigues, A.F.; Delwing-de Lima, D.; da Cruz, J.G.P.; Wyse, A.T.S.; Pitz, H.S.; Zeni, A.L.B. Protective effect of green tea extract against proline-induced oxidative damage in the rat kidney. Biomed. Pharmacother. 2016, 83, 1422–1427. [Google Scholar] [CrossRef]
- Zhao, X.; Li, G.; Hu, Y.; Yi, R.; Song, J. Improvement effects and mechanism research of polyphenol extracts from Kudingcha on carbon tetrachloride induced hepatic damage in mice. Sci. Technol. Food Ind. 2018, 39, 289–295. [Google Scholar]
- Zheng, X. Progress in research on anti-oxidation resistence of tea polyphenols and its protection effect of experimental liver injury. Chin. J. Clin. Ration. Drug Use 2011, 4, 126–128. [Google Scholar]
- Xu, L.; Shi, P.T.; Fu, X.S.; Cui, H.F.; Ye, Z.H.; Cai, C.B.; Yu, X.P. Protected geographical indication identification of a Chinese green tea (Anji-white) by near-infrared spectroscopy and chemometric class modeling techniques. J. Spectrosc. 2013, 2013, 1–8. [Google Scholar] [CrossRef]
- Lin, Y.; Xiao, Z.; Tian, B.; Hao, N.; Wang, J. Callus proliferation and accumulation of tea polyphenols of Anji Baicha. J. Hebei Univ. (Nat. Sci. Ed.) 2017, 37, 614–620. [Google Scholar]
- Li, Z.; He, J.; Liu, Q.; Yang, Y.; Li, C. The antioxidant effect of polyphenols of Anji white tea on D-galactose induced aging model mice. J. Guiyang Coll. Tradit. Chin. Med. 2016, 38, 21–24. [Google Scholar]
- Qian, Y.; Zhang, J.; Fu, X.; Yi, R.; Sun, P.; Zou, M.; Long, X.; Zhao, X. Preventive effect of raw Liubao tea polyphenols on mouse gastric injuries induced by HCl/ethanol via anti-oxidative stress. Molecules 2018, 23, 2848. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, J.; Yi, R.; Mu, J.; Zhou, X.; Zhao, X. Preventive effect of alkaloids from Lotus plumule on acute liver injury in mice. Foods 2019, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, X.; Chen, B.; Long, X.; Mu, J.; Pan, Y.; Song, J.-L.; Zhao, X.; Yang, Z. Preventive effect of Lactobacillus plantarum CQPC10 on activated carbon induced constipation in Institute of Cancer Research (ICR) mice. Appl. Sci. 2018, 8, 1498. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, D.; Wang, H.; Di, L.; Zhou, X.; Xu, T.; Yang, X.; Liu, Y. The hepatoprotective and antifibrotic effects of Saururus chinensis against carbon tetrachloride induced hepatic fibrosis in rats. J. Ethnopharmacol. 2009, 126, 487–491. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Huang, R.; Zhang, S.; Zheng, L.; Wei, L.; He, M.; Zhou, Y.; Zhuo, L.; Huang, Q. Methyl helicterate protects against CCl4-induced liver injury in rats by inhibiting oxidative stress, NF-κB activation, Fas/FasL pathway and cytochrome P4502E1 level. Food Chem. Toxicol. 2012, 50, 3413–3420. [Google Scholar] [CrossRef]
- Dong, D.; Zhang, S.; Yin, L.; Tang, X.; Xu, Y.; Han, X.; Qi, Y.; Peng, J. Protective effects of the total saponins from Rosa laevigata Michx fruit against carbon tetrachloride-induced acute liver injury in mice. Food Chem. Toxicol. 2013, 62, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kim, H.H.; Ku, S.K. Hepatoprotective effects of Artemisiae capillaris herba and Picrorrhiza rhizoma combinations on carbon tetrachloride-induced subacute liver damage in rats. Nutr. Res. 2008, 28, 270–277. [Google Scholar] [CrossRef]
- Pan, Y.; Long, X.; Yi, R.; Zhao, X. Polyphenols in Liubao tea can prevent CCl4-induced hepatic damage in mice through its antioxidant capacities. Nutrients 2018, 10, 1280. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Qian, Y.; Li, G.J.; Tan, J. Preventive effects of the polysaccharide of Larimichthys crocea swim bladder on carbon tetrachloride (CCl4)-induced hepatic damage. Chin. J. Nat. Med. 2015, 13, 521–528. [Google Scholar] [CrossRef]
- Cai, L.; Chen, W.N.; Li, R.; Hu, C.M.; Lei, C.; Li, C.M. Therapeutic effect of acetazolamide, an aquaporin 1 inhibitor, on adjuvant-induced arthritis in rats by inhibiting NF-B signal pathway. Immunopharmacol. Immunotoxicol. 2018, 40, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.; Waibler, Z.; Pleli, T.; Pfeilschifter, J.; Mühl, H. Type I interferon supports inducible nitric oxide synthase in murine hepatoma cells and hepatocytes and during experimental acetaminophen-induced liver damage. Front. Immunol. 2017, 8, 890. [Google Scholar] [CrossRef]
- Araújo Júnior, R.F.; Garcia, V.B.; Leitão, R.F.; Brito, G.A.; Miguel Ede, C.; Guedes, P.M.; de Araújo, A.A. Carvedilol improves inflammatory response, oxidative stress and fibrosis in the alcohol-induced liver injury in rats by regulating Kuppfer cells and hepatic stellate cells. PLoS ONE 2016, 12, e0148868. [Google Scholar] [CrossRef] [PubMed]
- Sipes, I.G.; el Sisi, A.E.; Sim, W.W.; Mobley, S.A.; Earnest, D.L. Reactive oxygen species in the progression of CCl4-induced liver injury. Adv. Exp. Med. Biol. 1991, 283, 489–497. [Google Scholar] [PubMed]
- Bonthius, D.J., Jr.; Winters, Z.; Karacay, B.; Bousquet, S.L.; Bonthius, D.J. Importance of genetics in fetal alcohol effects: Null mutation of the nNOS gene worsens alcohol-induced cerebellar neuronal losses and behavioral deficits. Neurotoxicology 2015, 46, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Hyun, D.H.; Jenner, P.; Halliwell, B. Effect of proteasome inhibition on cellular oxidative damage, antioxidant defences and nitric oxide production. J. Neurochem. 2001, 78, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Kosenko, E.A.; Tikhonova, L.A.; Alilova, G.A.; Montoliu, C.; Barreto, G.E.; Aliev, G.; Kaminsky, Y.G. Portacaval shunting causes differential mitochondrial superoxide production in brain regions. Free Radic. Biol. Med. 2017, 113, 109–118. [Google Scholar] [CrossRef]
- Selvaratnam, J.S.; Robaire, B. Effects of aging and oxidative stress on spermatozoa of superoxide-dismutase 1- and catalase-null mice. Biol. Reprod. 2016, 95, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, W.; Kedziora, J.; Zolynski, K.; Kedziora-Kornatowska, K.; Blaszczyk, J.; Witkowski, P.; Zieleniewski, J. Effect of long term bed rest in men on enzymatic antioxidative defence and lipid peroxidation in erythrocytes. J. Gravit. Physiol. 1998, 5, 163–164. [Google Scholar]
- Berndt, C.; Lillig, C.H. Glutathione, glutaredoxins, and iron. Antioxid. Redox. Signal. 2017, 27, 1235–1251. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Medina, J.P.; Zenteno-Savín, T.; Forman, H.J.; Crocker, D.E.; Ortiz1, R.M. Prolonged fasting increases glutathione biosynthesis in postweaned northern elephant seals. J. Exp. Biol. 2011, 214, 1294–1299. [Google Scholar] [CrossRef] [Green Version]
- Mihas, A.A.; Kanji, V.K.; Mihas, T.A.; Joseph, R.M.; Markov, A.K.; Heuman, D.M. Fructose diphosphate attenuates the acetaminophen-induced liver injury in the rat evidence for involvement of nitric oxide. Res. Commun. Mol. Pathol. Pharmacol. 2003, 113–114, 253–266. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence |
|---|---|
| COX-2 | Forward: 5′-GGTGCCTGGTCTGATGATG-3′ |
| Reverse: 5′-TGCTGGTTTGGAATAGTTGCT-3′ | |
| iNOS | Forward: 5′-GTTCTCAGCCCAACAATACAAGA-3′ |
| Reverse: 5′-GTGGACGGGTCGATGTCAC-3′ | |
| IL-1β | Forward: 5′-CTCCATGAGCTTTGTACAAGG-3′ |
| Reverse: 5′-TGCTGATGTACCAGTTGGGG-3′ | |
| TNF-α | Forward: 5′-GACCCTCAGACTCAGATCATCCTTCT-3′ |
| Reverse: 5′-ACGCTGGCTCAGCCACTC-3′ | |
| NF-κB | Forward: 5′-ATGGCAGACGATGATCCCTAC-3′ |
| Reverse: 5′ CGGAATCGAAATCCCCTCTGTT-3′ | |
| IκB-α | Forward: 5′-TGAAGGACGAGGAGTACGAGC-3′ |
| Reverse: 5′-TGCAGGAACGAGTCTCCGT-3′ | |
| Cu/Zn-SOD | Forward: 5′-AACCAGTTGTGTTGTCAGGAC-3′ |
| Reverse: 5′-CCACCATGTTTCTTAGAGTGAGG-3′ | |
| Mn-SOD | Forward: 5′-CAGACCTGCCTTACGACTATGG-3′ |
| Reverse: 5′-CTCGGTGGCGTTGAGATTGTT-3′ | |
| CAT | Forward: 5′-GGAGGCGGGAACCCAATAG-3′ |
| Reverse: 5′-GTGTGCCATCTCGTCAGTGAA-3′ | |
| GAPDH | Forward: 5′-AGGTCGGTGTGAACGGATTTG-3′ |
| Reverse: 5′-GGGGTCGTTGATGGCAACA-3′ |
| Group | Body Weight (g) | Liver Weight (g) | Liver Index |
|---|---|---|---|
| Normal | 38.776 ± 5.485 a | 1.422 ± 0.107 c | 3.651 ± 0.361 e |
| Model | 39.351 ± 2.036 a | 2.278 ± 0.158 a | 5.618 ± 0.339 a |
| Silymarin | 39.257 ± 2.890 a | 1.587 ± 0.155 b | 4.046 ± 0.045 d |
| LAJWTP | 38.223 ± 2.557 a | 2.011 ± 0.127 a | 5.082 ± 0.092 b |
| HAJWTP | 37.432 ± 3.994 a | 1.578 ± 0.085 b | 4.459 ± 0.033 c |
| Group | AST (Karmen/mL) | ALT (Karmen/mL) | TC (mg/dL) | TG (µmol/dL) |
|---|---|---|---|---|
| Normal | 29.49 ± 3.09 e | 18.30 ± 0.84 e | 101.67 ± 28.43 b | 58.23 ± 2.40 b |
| Model | 303.65 ± 15.65 a | 361.55 ± 1.69 a | 165.00 ± 47.7 a | 67.71 ± 10.65 a |
| Silymarin | 155.30 ± 6.06 d | 42.74 ± 15.08 d | 105.00 ± 25.98 b | 63.99 ± 3.14 ab |
| LAJWTP | 256.75 ± 4.28 b | 294.49 ± 0.82 b | 130.00 ± 20.00 ab | 66.37 ± 0.68 a |
| HAJWTP | 170.48 ± 5.28 c | 112.60 ± 33.11 c | 113.33 ± 30.14 b | 64.29 ± 1.61 ab |
| Group | SOD (U/mL) | GSH-Px (U/mL) | MDA (nmol/mL) |
|---|---|---|---|
| Normal | 218.71 ± 10.66 a | 221.85 ± 12.08 a | 3.39 ± 0.41 e |
| Model | 59.72 ± 4.32 e | 70.89 ± 5.25 e | 37.65 ± 1.71 a |
| Silymarin | 148.38 ± 7.59 b | 155.29 ± 7.16 b | 13.57 ± 0.89 d |
| LAJWTP | 79.81 ± 5.50 d | 82.55 ± 5.73 d | 28.99 ± 1.62 b |
| HAJWTP | 122.36 ± 6.63 c | 138.71 ± 4.39 c | 18.37 ± 1.30 c |
| Group | SOD (U/mg prot) | GSH-Px (U/mg prot) | MDA (nmol/mg prot) |
|---|---|---|---|
| Normal | 93.57 ± 5.87 a | 192.67 ± 8.71 a | 1.03 ± 0.12 e |
| Model | 21.32 ± 3.88 e | 48.36 ± 4.30 e | 9.15 ± 0.43 a |
| Silymarin | 79.35 ± 4.79 b | 125.77 ± 6.68 b | 3.41 ± 0.25 d |
| LAJWTP | 40.98 ± 3.87 d | 73.58 ± 5.29 d | 7.05 ± 0.28 b |
| HAJWTP | 66.72 ± 5.05 c | 112.33 ± 4.21 c | 4.32 ± 0.21 c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Yang, Z.; Zhang, J.; Mu, J.; Zhou, X.; Zhao, X. Liver Injury Induced by Carbon Tetrachloride in Mice Is Prevented by the Antioxidant Capacity of Anji White Tea Polyphenols. Antioxidants 2019, 8, 64. https://doi.org/10.3390/antiox8030064
Wang R, Yang Z, Zhang J, Mu J, Zhou X, Zhao X. Liver Injury Induced by Carbon Tetrachloride in Mice Is Prevented by the Antioxidant Capacity of Anji White Tea Polyphenols. Antioxidants. 2019; 8(3):64. https://doi.org/10.3390/antiox8030064
Chicago/Turabian StyleWang, Ranran, Zhiqing Yang, Jing Zhang, Jianfei Mu, Xianrong Zhou, and Xin Zhao. 2019. "Liver Injury Induced by Carbon Tetrachloride in Mice Is Prevented by the Antioxidant Capacity of Anji White Tea Polyphenols" Antioxidants 8, no. 3: 64. https://doi.org/10.3390/antiox8030064
APA StyleWang, R., Yang, Z., Zhang, J., Mu, J., Zhou, X., & Zhao, X. (2019). Liver Injury Induced by Carbon Tetrachloride in Mice Is Prevented by the Antioxidant Capacity of Anji White Tea Polyphenols. Antioxidants, 8(3), 64. https://doi.org/10.3390/antiox8030064