Phenolic Constitution, Phytochemical and Macronutrient Content in Three Species of Microgreens as Modulated by Natural Fiber and Synthetic Substrates

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Standards Preparation
2.2. Plant Material and Climate Chamber Conditions
2.3. Growing Substrates Physicochemical Characterization
2.4. Harvesting Schedule, Sampling, Growth Analysis and Colorimeter Measurement
2.5. Mineral Analysis, Nitrate, Total Chlorophyll and Total Ascorbic Acid
2.6. Carotenoids and Polyphenols Extraction and Quantification
2.7. Statistics
3. Results and Discussion
3.1. Substrate Physicochemical Properties
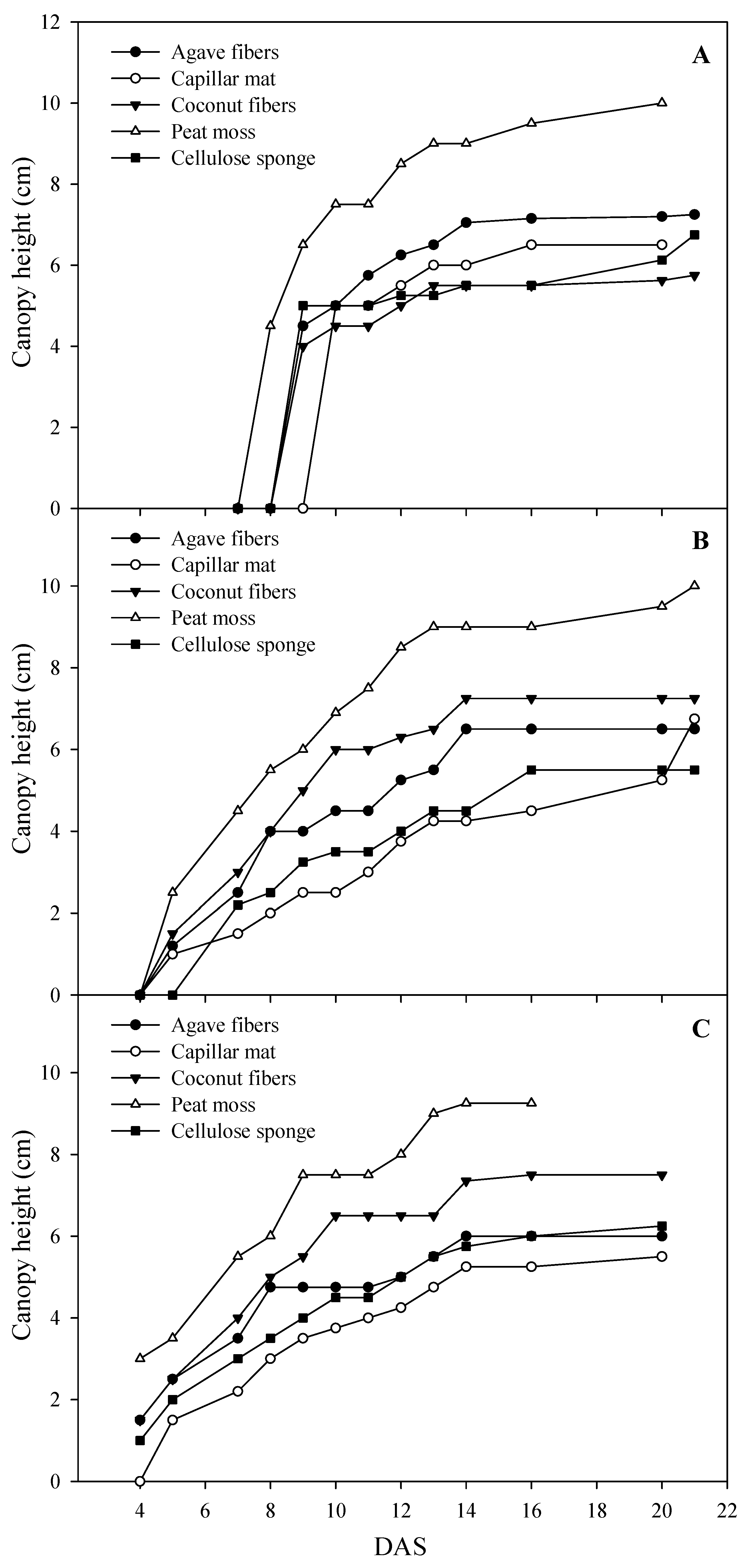
3.2. Fresh Biomass Yield, Dry Matter Content, Canopy Height and Color
3.3. Nitrate Concentration and Mineral Composition
3.4. Chlorophyll, Carotenoid and Ascorbate Content
3.5. Phenolic Composition
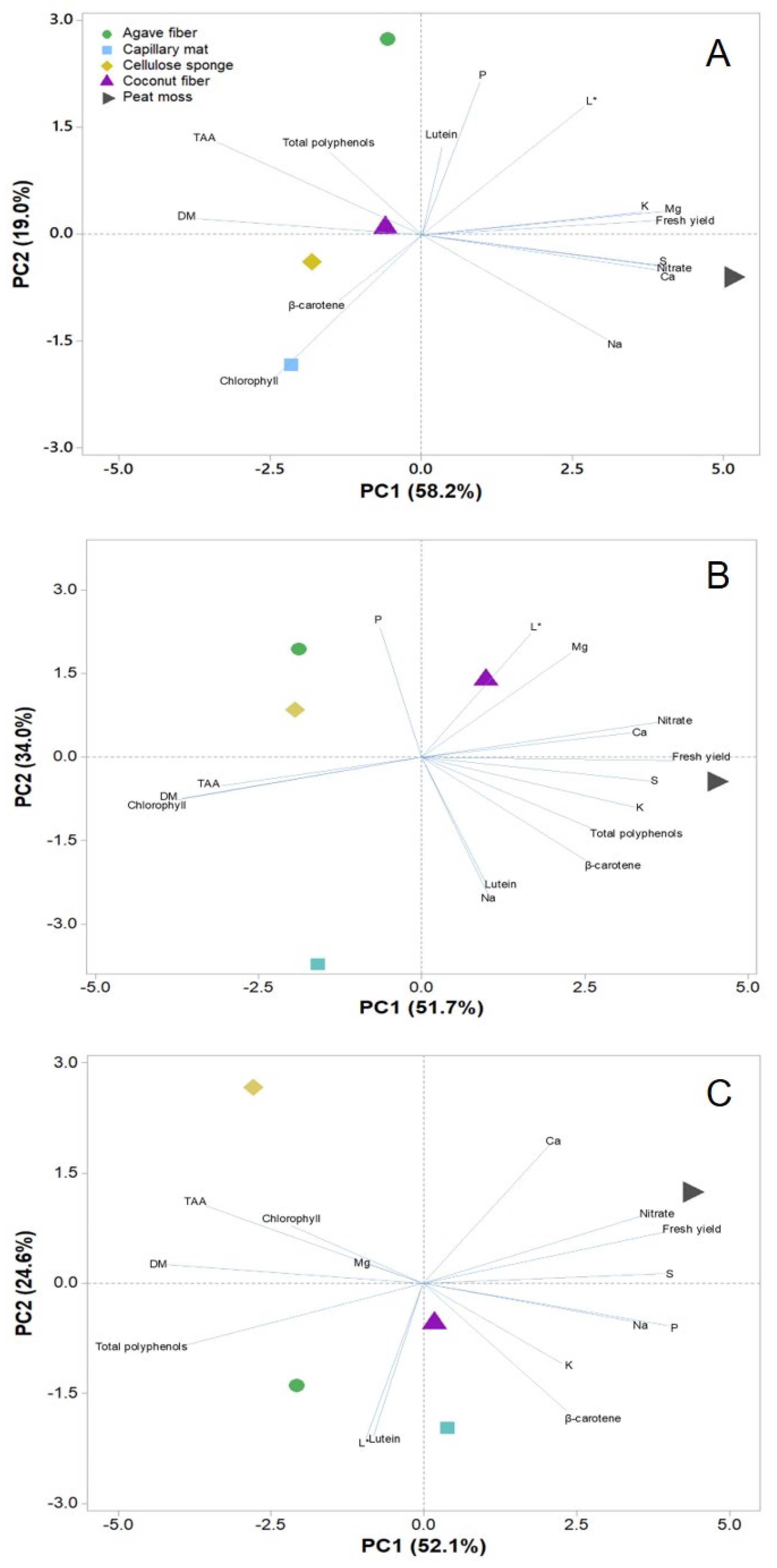
3.6. Principal Component Analysis of Nutritional and Functional Quality Parameters in Response to Substrate for Coriander, Kohlrabi and Pak Choi Microgreens
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Poiroux-Gonord, F.; Bidel, L.P.; Fanciullino, A.L.; Gautier, H.; Lauri-Lopez, F.; Urban, L. Health benefits of vitamins and secondary metabolites of fruits and vegetables and prospects to increase their concentrations by agronomic approaches. J. Agric. Food Chem. 2010, 58, 12065–12082. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; de Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Xiao, Z.; Lester, G.E.; Luo, Y.; Wang, Q. Assessment of vitamin and carotenoid concentrations of emerging food products: Edible microgreens. J. Agric. Food Chem. 2012, 60, 7644–7651. [Google Scholar] [CrossRef]
- Xiao, Z.; Lester, G.E.; Park, E.; Saftner, R.A.; Luo, Y.; Wang, Q. Evaluation and correlation of sensory attributes and chemical compositions of emerging fresh produce: Microgreens. Postharvest Biol. Technol. 2015, 110, 140–148. [Google Scholar] [CrossRef]
- Di Gioia, F.; Mininni, C.; Santamaria, P. How to grow microgreens. In Microgreens: Novel Fresh and Functional Food to Explore All the Value of Biodiversity; di Gioia, F., Santamaria, P., Eds.; Eco-Logica Srl: Bari, Italy, 2015; pp. 51–79. [Google Scholar]
- Pinto, E.; Almeida, A.A.; Aguiar, A.A.; Ferreira, I.M.P.L.V.O. Comparison between the mineral profile and nitrate content of microgreens and mature lettuces. J. Food Comp. Anal. 2015, 37, 38–43. [Google Scholar] [CrossRef]
- Choe, U.; Yu, L.L.; Wang, T.T.Y. The science behind microgreens as an exciting new food for the 21st century. J. Agric. Food Chem. 2018, 66, 11519–11530. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, A.; Orlandini, A.; Bulgari, R.; Ferrante, A.; Bruschi, P. Antioxidant and mineral composition of three wild leafy species: A comparison between microgreens and baby greens. Foods 2019, 8, 487. [Google Scholar] [CrossRef] [Green Version]
- Kyriacou, M.C.; Rouphael, Y.; di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; de Pascale, S. Micro-scale vegetable production and the rise of microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; de Pascale, S.; Kyratzis, A.; Rouphael, Y. Microgreens as a component of space life support systems: A cornucopia of functional food. Front. Plant Sci. 2017, 8, 1587. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Sakalauskiené, S.; Samuoliené, G.; Jankauskienė, J.; Viršilė, A.; Novičkovas, A.; Sirtautas, R.; Miliauskienė, J.; Vaštakaitė, V.; Dabašinskas, L.; et al. The effects of LED illumination spectra and intensity on carotenoid content in Brassicaceae microgreens. Food Chem. 2015, 173, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; de Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Genotype-specific modulatory effects of select spectral bandwidths on the nutritive and phytochemical composition of microgreens. Front. Plant Sci. 2019, 10, 1501. [Google Scholar] [CrossRef]
- Samuoliene, G.; Brazaitytė, A.; Viršilė, A.; Jankauskienė, J.; Sakalauskiene, S.; Duchovskis, P. Red light-dose or wavelength-dependent photoresponse of antioxidants in herb microgreens. PLoS ONE 2016, 11, e0163405. [Google Scholar] [CrossRef]
- Samuolienė, G.; Viršilė, A.; Brazaitytė, A.; Jankauskienė, J.; Sakalauskienė, S.; Vaštakaitė, V.; Novičkovas, A.; Viškelienė, A.; Sasnauskas, A.; Duchovskis, P. Blue light dosage affects carotenoids and tocopherols in microgreens. Food Chem. 2017, 228, 50–56. [Google Scholar] [CrossRef]
- Craver, J.K.; Gerovac, J.R.; Lopez, R.G.; Kopsell, D.A. Light intensity and light quality from sole-source light-emitting diodes impact phytochemical concentrations within Brassica microgreens. J. Am. Soc. Hortic. Sci. 2017, 142, 3–12. [Google Scholar] [CrossRef]
- Vaštakaitė, V.; Viršilė, A.; Brazaitytė, A.; Samuolienė, G.; Jankauskienė, J.; Novičkovas, A.; Duchovskis, P. Pulsed light emitting diodes for a higher phytochemical level in microgreens. J. Agric. Food Chem. 2017, 65, 6529–6534. [Google Scholar] [CrossRef]
- Alrifai, O.; Hao, X.; Marcone, M.F.; Tsao, R. Current review of the modulatory effects of LED lights on photosynthesis of secondary metabolites and future perspectives of microgreen vegetables. J. Agric. Food Chem. 2019, 67, 6075–6090. [Google Scholar] [CrossRef]
- De la Fuente, B.; López-García, G.; Máñez, V.; Alegría, A.; Barberá, R.; Cilla, A. Evaluation of the bioaccessibility of antioxidant bioactive compounds and minerals of four genotypes of Brassicaceae Microgreens. Foods 2019, 8, 250. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Rausch, S.R.; Luo, Y.; Sun, J.; Yu, L.; Wang, Q.; Chen, P.; Yu, L.; Stommel, J.R. Microgreens of Brassicaceae: Genetic diversity of phytochemical concentrations and antioxidant capacity. LWT 2019, 101, 731–737. [Google Scholar] [CrossRef]
- Di Gioia, F.; De Bellis, P.; Mininni, C.; Santamaria, P.; Serio, F. Physicochemical, agronomical and microbiological evaluation of alternative growing media for the production of rapini (Brassica rapa L.) microgreens. J. Sci. Food Agric. 2017, 97, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Muchjajib, U.; Muchjajib, S.; Suknikom, S.; Butsai, J. Evaluation of organic media alternatives for the production of microgreens in Thailand. Acta Hortic. 2015, 1102, 157–162. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; de Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1. [Google Scholar] [CrossRef]
- Kampfenkel, K.; van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Kim, H.J.; Fonseca, J.M.; Choi, J.H.; Kubota, C.; Kwon, D.Y. Salt in irrigation water affects the nutritional and visual properties of Romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef]
- Llorach, R.; Martínez-Sánchez, A.; Tomás-Barberán, F.A.; Gil, M.I.; Ferreres, F. Characterisation of polyphenols and antioxidant properties of five lettuce varieties and escarole. Food Chem. 2008, 108, 1028–1038. [Google Scholar] [CrossRef]
- Abad, M.; Noguera, P.; Bures, S. National inventory of organic wastes for use as growing media for ornamental potted plant production: Case study in Spain. Bioresour. Technol. 2011, 77, 197–200. [Google Scholar] [CrossRef]
- Barrett, G.E.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems—A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Savvas, D. Modern developments in the use of inorganic media for greenhouse vegetable and flower production. Acta Hort. 2007, 819, 73–86. [Google Scholar] [CrossRef]
- Schmilewski, G. The role of peat in assuring the quality of growing media. Mires Peat 2008, 3, 1–8. [Google Scholar]
- Sánchez-Monedero, M.A.; Roig, A.; Cegarra, J.; Bernal, M.P.; Noguera, P.; Abad, M.; Antón, A. Composts as media constituents for vegetable transplant production. Compost Sci. Utilization 2004, 12, 161–168. [Google Scholar] [CrossRef]
- Shannon, M.C.; Grieve, C.M. Tolerance of vegetable crops to salinity. Sci. Hortic. 1998, 78, 5–38. [Google Scholar] [CrossRef]
- Okkaoğlu, H.; Sönmez, Ç.; Şimşek, A.Ö.; Bayram, E. Effect of salt stress on some agronomical characteristics and essential oil content of coriander (Coriandrum sativum L.) cultivars. J. Appl. Biol. Sci. JABS 2015, 9, 21–24. [Google Scholar]
- Bulgari, R.; Baldi, A.; Ferrante, A.; Lenzi, A. Yield and quality of basil, Swiss chard, and rocket microgreens grown in a hydroponic system. N. Z. J. Crop Hortic. Sci. 2017, 45, 119–129. [Google Scholar] [CrossRef]
- Carillo, P.; Raimondi, G.; Kyriacou, M.C.; Pannico, A.; El-Nakhel, C.; Cirillo, V.; Colla, G.; de Pascale, S.; Rouphael, Y. Morpho-physiological and homeostatic adaptive responses triggered by omeprazole enhance lettuce tolerance to salt stress. Sci. Hortic. 2019, 249, 22–30. [Google Scholar] [CrossRef]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. J. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Colla, G.; Kim, H.J.; Kyriacou, M.C.; Rouphael, Y. Nitrate in fruits and vegetables. Sci. Hortic. 2018, 237, 221–238. [Google Scholar] [CrossRef]
- Raviv, M.; Wallach, R.; Silber, A.; Bar-Tal, A. Substrates and their analysis. Hydroponic Prod. Veg. Ornam. 2002, 2, 25–105. [Google Scholar]
- Commission Regulation (EC) No. 1881/2006 of 19 December Setting Maximum Levels for Certain Contaminants in Foodstuffs; European Commission: Brussels, Belgium, 2006.
- EFSA. Opinion of the scientific panel on contaminants in the food chain on a request from the European commission to perform a scientific risk assessment on nitrate in vegetables. EFSA J. 2008, 689, 1–79. [Google Scholar]
- Gharibzahedi, S.M.T.; Jafari, S.M. The importance of minerals in human nutrition bioavailability, food fortification, processing effects and nanoencapsulation. Trends Food Sci. Technol. 2017, 62, 119–132. [Google Scholar] [CrossRef]
- Levander, O.A. Fruit and vegetable contributions to dietary mineral intake in human health and disease. HortScience 1990, 25, 1486–1488. [Google Scholar] [CrossRef]
- Xiao, Z.; Codling, E.E.; Luo, Y.; Nou, X.; Lester, G.E.; Wang, Q. Microgreens of Brassicaceae: Mineral composition and content of 30 varieties. J. Food Compos. Anal. 2016, 49, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Neugart, S.; Baldermann, S.; Hanschen, F.S.; Klopsch, R.; Wiesner-Reinhold, M.; Schreiner, M. The intrinsic quality of brassicaceous vegetables: How secondary plant metabolites are affected by genetic, environmental, and agronomic factors. Sci. Hortic. 2018, 233, 460–478. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef]
- Kvansakul, J.; Rodriguez-Carmona, M.; Edgar, D.F.; Barker, F.M.; Koepcke, W.; Schalch, W.; Barbur, J.L. Supplementation with the carotenoids lutein or zeaxanthin improves human visual performance. Ophthalmic Physiol. Opt. 2006, 26, 362–371. [Google Scholar] [CrossRef]
- Saltveit, M.E.; Choi, Y.J.; Tomas-Barberan, F.A. Involvement of components of the phosopholipid-signalling pathway in wound-induced phenylpropanoid metabolism in lettuce (Lactuca sativa) leaf tissue. Physiol. Plant. 2005, 125, 345–355. [Google Scholar] [CrossRef]
- Vallejo, F.; García-Viguera, C.; Tomás-Barberán, F.A. Changes in broccoli (Brassica oleracea L. var. italica) health-promoting compounds with inflorescence development. J. Agric. Food Chem. 2003, 51, 3776–3782. [Google Scholar] [CrossRef]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef]
- Onakpoya, I.J.; Spencer, E.A.; Thompson, M.J.; Heneghan, C.J. The effect of chlorogenic acid on blood pressure: A systematic review and meta-analysis of randomized clinical trials. J. Hum. Hypertens. 2014, 29, 77–81. [Google Scholar] [CrossRef]
- Tajik, N.; Takik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017, 56, 2215–2244. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, B.D.; Stommel, J.R. Distribution of hydroxycinnamic acid conjugates in fruit of commercial eggplant (Solanum melongena L.) cultivars. J. Agric. Food Chem. 2003, 51, 3448–3454. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.S.; Escher, G.B.; do Carmo, M.V.; Azevedo, L.; Marques, M.B.; Daguer, H.; Oh, W.Y. A new analytical concept based on chemistry and toxicology for herbal extracts analysis: From phenolic composition to bioactivity. Food Res. Int. 2020. [Google Scholar] [CrossRef]
- Alfaifi, M.; Alsayari, A.; Gurusamy, N.; Louis, J.; Eldin Elbehairi, S.; Venkatesan, K.; Alboushnak, H. Analgesic, anti-Inflammatory, cytotoxic activity screening and UPLC-PDA-ESI-MS metabolites determination of bioactive fractions of Kleinia pendula. Molecules 2020, 25, 418. [Google Scholar] [CrossRef] [Green Version]
- Terao, J.; Yamaguchi, S.; Shirai, M.; Miyoshi, M.; Moon, J.H.; Oshima, S.; Inakuma, T.; Tsushida, T.; Kato, Y. Protection by quercetin and quercetin 3-O-β-D-glucuronide of peroxynitrite-induced antioxidant consumption in human plasma low-density lipoprotein. Free Radic. Res. 2001, 35, 925–931. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2011, 16, 251–280. [Google Scholar] [CrossRef]
- Olsen, H.; Aaby, K.; Borge, G.I. Characterization and quantification of flavonoids and hydroxycinnamic acids in curly kale (Brassica oleracea L. convar. acephala var. sabellica) by Hplc-Dadesi-Msn. J. Agric. Food Chem. 2009, 57, 2816–2825. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Func. Food 2018, 40, 68–75. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Giordano, M.; de Pascale, S.; Rouphael, Y. Macronutrient deprivation eustress elicits differential secondary metabolites in red and green-pigmented butterhead lettuce grown in closed soilless system. J. Sci. Food Agric. 2019, 99, 6962–6972. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Physicochemical Parameters | Substrate | Significance | ||||
|---|---|---|---|---|---|---|
| Agave Fiber | Capillary Mat | Cellulose Sponge | Coconut Fiber | Peat Moss | ||
| Bulk density (kg m−3) | 144.2 ± 4.49b | 240.7 ± 6.24a | 39.6 ± 0.23d | 112.4 ± 0.19c | 147.8 ± 1.15b | *** |
| Total pore space (% v/v) | 95.6 ± 0.28a | 86.4 ± 0.52d | 93.8 ± 0.42b | 90.8 ± 0.16c | 87.4 ± 0.16d | *** |
| Water-holding capacity (% v/v) | 76.6 ± 0.3b | 71.1 ± 0.5d | 80.2 ± 0.58a | 74 ± 0.12c | 58.8 ± 0.27e | *** |
| Air capacity (% v/v) | 19 ± 0.25b | 15.3 ± 0.21d | 13.7 ± 0.61e | 16.8 ± 0.07c | 28.7 ± 0.27a | *** |
| EC (μS cm−1) | 254 ± 0.50e | 1258 ± 8.00a | 702 ± 0.50c | 879 ± 1.00b | 282 ± 1.50d | *** |
| pH | 4.90 ± 0.005e | 5.71 ± 0.010b | 8.49 ± 0.010a | 5.34 ± 0.005d | 5.48 ± 0.005c | *** |
| NO3 (g kg−1 dw) | 9.07 ± 0.21a | 0.54 ± 0.11d | 1.16 ± 0.06c | 2.63 ± 0.18b | 0.24 ± 0.05d | *** |
| P (g kg−1 dw) | 0.67 ± 0.03c | 0.07 ± 0.00d | 0.01 ± 0.00e | 1.72 ± 0.00a | 1.04 ± 0.01b | *** |
| K (g kg−1 dw) | 15.00 ± 0.32b | 13.39 ± 0.03bc | 1.93 ± 0.92d | 81.41 ± 1.83a | 12.10 ± 0.88c | *** |
| Ca (g kg−1 dw) | 10.66 ± 0.02c | 3.44 ± 0.26e | 9.89 ± 0.09d | 16.67 ± 0.29b | 30.34 ± 0.19a | *** |
| Mg (g kg−1 dw) | 4.92 ± 0.95b | 2.75 ± 0.74b | 12.23 ± 0.59a | 5.73 ± 0.33b | 2.66 ± 1.17b | ** |
| S (g kg−1 dw) | 0.98 ± 0.14d | 9.15 ± 0.00a | 1.72 ± 0.01c | 1.10 ± 0.16d | 6.09 ± 0.09b | *** |
| NH4 (g kg−1 dw) | 2.12 ± 0.06b | 3.66 ± 0.05a | 0.20 ± 0.16c | 1.68 ± 0.05b | 0.42 ± 0.16c | *** |
| Na (g kg−1 dw) | 6.72 ± 0.08d | 21.73 ± 0.16a | 12.18 ± 0.37c | 19.80 ± 0.11b | 3.89 ± 0.29e | *** |
| Cl (g kg−1 dw) | 9.58 ± 0.05c | 6.42 ± 0.77d | 37.65 ± 0.40b | 77.52 ± 1.09a | 2.05 ± 0.06e | *** |
| Source of Variance | Yield (kg fw m−2) | Dry Weight (g m−2) | Dry Matter (%) | L* |
|---|---|---|---|---|
| Microgreens Species (M) | ||||
| Coriander | 1.31 ± 0.14b | 169.03 ± 11.06 | 13.59 ± 0.52a | 33.68 ± 0.30a |
| Kohlrabi | 1.80 ± 0.24a | 182.89 ± 17.63 | 10.71 ± 0.37b | 28.52 ± 0.26b |
| Pak choi | 1.86 ± 0.20a | 172.75 ± 8.18 | 10.12 ± 0.64b | 23.87 ± 0.27c |
| Substrate (S) | ||||
| Agave fiber | 1.18 ± 0.05c | 143.46 ± 8.92c | 12.16 ± 0.42b | 29.57 ± 1.39a |
| Capillary mat | 1.18 ± 0.12c | 134.73 ± 7.17c | 11.95 ± 0.68b | 27.98 ± 1.26c |
| Cellulose sponge | 1.31 ± 0.06c | 172.81 ± 5.22b | 13.30 ± 0.52a | 28.27 ± 1.57bc |
| Coconut fiber | 1.65 ± 0.18b | 178.71 ± 9.43b | 11.54 ± 0.94b | 28.72 ± 1.36abc |
| Peat moss | 2.96 ± 0.19a | 244.75 ± 17.17a | 8.43 ± 0.57c | 28.91 ± 1.64ab |
| M × S | ||||
| Coriander × Agave fiber | 1.28 ± 0.04ef | 175.33 ± 8.20cd | 13.66 ± 0.39b | 34.49 ± 0.19 |
| Coriander × Capillary mat | 0.80 ± 0.08g | 111.04 ± 8.59g | 13.91 ± 0.39ab | 32.55 ± 0.90 |
| Coriander × Cellulose sponge | 1.16 ± 0.12efg | 174.67 ± 14.95cd | 15.11 ± 0.32a | 33.63 ± 0.49 |
| Coriander × Coconut fiber | 1.02 ± 0.02fg | 153.66 ± 5.15def | 15.12 ± 0.74a | 33.27 ± 0.05 |
| Coriander × Peat moss | 2.27 ± 0.10b | 230.45 ± 13.00b | 10.12 ± 0.14ef | 34.49 ± 0.82 |
| Kohlrabi × Agave fiber | 1.08 ± 0.04fg | 121.70 ± 4.95fg | 11.29 ± 0.12de | 29.16 ± 0.48 |
| Kohlrabi × Capillary mat | 1.20 ± 0.02ef | 144.69 ± 8.70def | 12.07 ± 0.85cd | 27.21 ± 0.31 |
| Kohlrabi × Cellulose sponge | 1.41 ± 0.03ef | 164.00 ± 3.24de | 11.65 ± 0.10d | 28.32 ± 0.49 |
| Kohlrabi × Coconut fiber | 1.86 ± 0.23cd | 180.27 ± 20.61cd | 9.72 ± 0.42f | 28.93 ± 0.59 |
| Kohlrabi × Peat moss | 3.43 ± 0.18a | 303.77 ± 15.78a | 8.84 ± 0.05f | 28.98 ± 0.47 |
| Pak choi × Agave fiber | 1.17 ± 0.13efg | 133.33 ± 8.20efg | 11.54 ± 0.50d | 25.05 ± 0.68 |
| Pak choi × Capillary mat | 1.53 ± 0.17de | 148.44 ± 6.60def | 9.86 ± 0.76f | 24.19 ± 0.46 |
| Pak choi × Cellulose sponge | 1.37 ± 0.08ef | 179.77 ± 5.31cd | 13.14 ± 0.35bc | 22.85 ± 0.39 |
| Pak choi × Coconut fiber | 2.08 ± 0.13bc | 202.19 ± 5.12bc | 9.76 ± 0.43f | 23.97 ± 0.49 |
| Pak choi × Peat moss | 3.16 ± 0.14a | 200.02 ± 16.64bc | 6.31 ± 0.32g | 23.27 ± 0.11 |
| Significance | ||||
| Microgreens species (M) | *** | ns | *** | *** |
| Substrate (S) | *** | *** | *** | ** |
| M × S | *** | *** | *** | ns |
| Source of Variance | Nitrate (mg kg−1 fw) | P (g kg−1 dw) | K (g kg−1 dw) | Ca (g kg−1 dw) | Mg (g kg−1 dw) | S (g kg−1 dw) | Na (g kg−1 dw) |
|---|---|---|---|---|---|---|---|
| Microgreens species (M) | |||||||
| Coriander | 398.5 ± 80.0c | 4.14 ± 0.25b | 15.60 ± 0.77b | 6.46 ± 1.26c | 4.27 ± 0.21 | 0.98 ± 0.17c | 0.48 ± 0.10c |
| Kohlrabi | 500.1 ± 59.6b | 3.93 ± 0.18b | 10.88 ± 0.69c | 12.98 ± 1.21a | 4.25 ± 0.21 | 3.10 ± 0.22b | 1.78 ± 0.37b |
| Pak choi | 601.6 ± 52.9a | 4.92 ± 0.29a | 21.20 ± 1.31a | 9.58 ± 0.73b | 4.22 ± 0.11 | 4.03 ± 0.52a | 2.68 ± 0.47a |
| Substrate (S) | |||||||
| Agave fiber | 361.3 ± 48.5c | 4.68 ± 0.32a | 13.69 ± 1.52c | 8.04 ± 1.15c | 4.51 ± 0.19b | 2.23 ± 0.37bc | 0.44 ± 0.09d |
| Capillary mat | 324.3 ± 52.7c | 3.95 ± 0.38bc | 16.32 ± 1.94b | 6.12 ± 0.71d | 3.45 ± 0.16d | 2.64 ± 0.52b | 3.16 ± 0.66a |
| Cellulose sponge | 396.1 ± 60.7c | 3.70 ± 0.27c | 11.97 ± 1.29c | 9.43 ± 1.29b | 3.93 ± 0.14c | 1.82 ± 0.36c | 0.64 ± 0.14d |
| Coconut fiber | 530.5 ± 71.3b | 4.82 ± 0.20a | 19.06 ± 2.21a | 7.70 ± 0.89c | 4.42 ± 0.08b | 2.07 ± 0.38c | 1.25 ± 0.28c |
| Peat moss | 888.0 ± 31.8a | 4.49 ± 0.41ab | 18.43 ± 1.41a | 17.07 ± 1.16a | 4.92 ± 0.22a | 4.77 ± 0.80a | 2.75 ± 0.53b |
| M × S | |||||||
| Coriander × Agave fiber | 236.8 ± 25.2ef | 5.17 ± 0.92ab | 15.62 ± 1.50efg | 3.94 ± 0.29h | 4.26 ± 0.49cde | 0.80 ± 0.10d | 0.17 ± 0.01g |
| Coriander × Capillary mat | 186.5 ± 43.2f | 3.53 ± 0.22cd | 13.82 ± 1.07efg | 3.58 ± 0.42h | 3.61 ± 0.12f | 0.81 ± 0.09d | 0.69 ± 0.03def |
| Coriander × Cellulose sponge | 255.4 ± 15.9ef | 3.20 ± 0.06cd | 12.18 ± 0.19gh | 4.39 ± 0.10h | 3.70 ± 0.09ef | 0.48 ± 0.05d | 0.14 ± 0.00g |
| Coriander × Coconut fiber | 333.1 ± 49.0def | 4.65 ± 0.24abc | 16.77 ± 1.03def | 4.58 ± 0.31h | 4.27 ± 0.17cde | 0.62 ± 0.04d | 0.33 ± 0.02fg |
| Coriander × Peat moss | 980.6 ± 37.2a | 4.15 ± 0.11bcd | 19.61 ± 0.60cd | 15.80 ± 0.68b | 5.53 ± 0.19a | 2.19 ± 0.09c | 1.09 ± 0.04de |
| Kohlrabi × Agave fiber | 301.3 ± 6.3ef | 4.53 ± 0.20abc | 8.03 ± 0.26i | 11.79 ± 0.25de | 4.87 ± 0.16abc | 2.71 ± 0.08c | 0.36 ± 0.02efg |
| Kohlrabi × Capillary mat | 276.4 ± 4.6ef | 3.06 ± 0.13d | 11.54 ± 0.48hi | 8.31 ± 0.30f | 2.89 ± 0.08g | 2.79 ± 0.11c | 4.01 ± 0.34b |
| Kohlrabi × Cellulose sponge | 399.8 ± 15.6cde | 3.95 ± 0.30bcd | 7.99 ± 0.36i | 12.84 ± 0.21cd | 3.90 ± 0.05def | 2.67 ± 0.19c | 0.71 ± 0.10defg |
| Kohlrabi × Coconut fiber | 729.0 ± 64.9b | 4.64 ± 0.10abc | 13.07 ± 0.83fgh | 10.60 ± 0.23e | 4.49 ± 0.11bcd | 2.63 ± 0.20c | 1.21 ± 0.14d |
| Kohlrabi × Peat moss | 793.8 ± 24.8ab | 3.46 ± 0.12cd | 13.76 ± 0.56efg | 21.35 ± 1.12a | 5.08 ± 0.07ab | 4.71 ± 0.09b | 2.59 ± 0.06c |
| Pak choi × Agave fiber | 545.8 ± 31.6c | 4.32 ± 0.36bcd | 17.43 ± 0.64de | 8.37 ± 0.45f | 4.39 ± 0.21cde | 3.17 ± 0.26c | 0.78 ± 0.05defg |
| Pak choi × Capillary mat | 509.9 ± 59.6cd | 5.28 ± 0.52ab | 23.59 ± 1.70b | 6.49 ± 0.11g | 3.83 ± 0.22def | 4.31 ± 0.34b | 4.79 ± 0.55a |
| Pak choi × Cellulose sponge | 533.0 ± 156.4c | 3.96 ± 0.77bcd | 15.74 ± 2.19efg | 11.06 ± 0.15e | 4.19 ± 0.39cde | 2.32 ± 0.36c | 1.07 ± 0.08def |
| Pak choi × Coconut fiber | 529.5 ± 123.2c | 5.19 ± 0.54ab | 27.33 ± 1.40a | 7.92 ± 0.56f | 4.52 ± 0.14bcd | 2.95 ± 0.24c | 2.22 ± 0.21c |
| Pak choi × Peat moss | 889.6 ± 37.4ab | 5.85 ± 0.69a | 21.91 ± 2.39bc | 14.08 ± 0.15c | 4.16 ± 0.24cde | 7.41 ± 0.93a | 4.56 ± 0.54ab |
| Significance | |||||||
| Microgreens species (M) | *** | *** | *** | *** | ns | *** | *** |
| Substrate (S) | *** | ** | *** | *** | *** | *** | *** |
| M × S | *** | ** | ** | *** | *** | *** | *** |
| Source of Variance | Total Chlorophyll (mg kg−1 fw) | Lutein (mg kg−1 dw) | β-Carotene (mg kg−1 dw) | Total Ascorbic Acid (mg ascorbic acid kg−1 dw) |
|---|---|---|---|---|
| Microgreens species (M) | ||||
| Coriander | 13.63 ± 0.38a | 98.6 ± 8.8b | 325.1 ± 38.1a | 121.40 ± 5.77c |
| Kohlrabi | 8.85 ± 0.37b | 73.2 ± 5.9c | 183.1 ± 15.5c | 199.87 ± 8.77a |
| Pak choi | 14.33 ± 0.59a | 122.6 ± 7.5a | 236.8 ± 19.0b | 177.12 ± 5.38b |
| Substrate (S) | ||||
| Agave fiber | 12.59 ± 0.90ab | 109.5 ± 15.7a | 233.2 ± 31.6b | 179.79 ± 12.16a |
| Capillary mat | 13.13 ± 0.89a | 112.3 ± 9.8a | 351.1 ± 56.0a | 173.37 ± 13.46a |
| Cellulose sponge | 13.07 ± 1.17a | 70.4 ± 8.2b | 153.8 ± 11.0c | 187.39 ± 16.08a |
| Coconut fiber | 11.16 ± 0.89b | 103.3 ± 8.6a | 250.3 ± 28.9b | 150.35 ± 9.77b |
| Peat moss | 11.39 ± 1.17b | 95.1 ± 11.0a | 253.2 ± 23.9b | 139.75 ± 14.03b |
| M × S | ||||
| Coriander × Agave fiber | 12.70 ± 1.04 | 121.0 ± 23.5 | 312.9 ± 36.0bc | 146.16 ± 3.23bcd |
| Coriander × Capillary mat | 15.03 ± 0.84 | 99.4 ± 15.8 | 533.3 ± 103.1a | 128.92 ± 4.93cd |
| Coriander × Cellulose sponge | 13.76 ± 0.24 | 57.8 ± 7.2 | 169.1 ± 18.6def | 128.01 ± 6.92cd |
| Coriander × Coconut fiber | 13.78 ± 0.18 | 120.4 ± 13.2 | 355.2 ± 25.4b | 116.41 ± 8.80d |
| Coriander × Peat moss | 12.89 ± 1.22 | 94.3 ± 19.3 | 255.0 ± 33.3bcd | 87.52 ± 7.88e |
| Kohlrabi × Agave fiber | 9.68 ± 0.32 | 58.0 ± 7.3 | 134.9 ± 16.7ef | 219.14 ± 19.37a |
| Kohlrabi × Capillary mat | 10.03 ± 0.99 | 101.9 ± 16.5 | 234.5 ± 29.3bcd | 217.77 ± 13.18a |
| Kohlrabi × Cellulose sponge | 9.35 ± 0.30 | 52.2 ± 4.2 | 122.6 ± 15.9f | 226.75 ± 11.88a |
| Kohlrabi × Coconut fiber | 7.85 ± 0.23 | 73.8 ± 6.5 | 180.8 ± 18.6cde | 158.42 ± 11.07bc |
| Kohlrabi × Peat moss | 7.34 ± 0.92 | 79.8 ± 8.2 | 242.7 ± 22.9bcd | 177.27 ± 7.44b |
| Pak choi × Agave fiber | 15.40 ± 0.50 | 149.4 ± 13.3 | 252.0 ± 46.8bcd | 174.08 ± 5.81b |
| Pak choi × Capillary mat | 14.34 ± 0.77 | 135.5 ± 14.8 | 285.6 ± 24.9bcd | 173.42 ± 1.85b |
| Pak choi × Cellulose sponge | 16.09 ± 2.10 | 101.2 ± 3.6 | 169.6 ± 10.9def | 207.41 ± 13.28a |
| Pak choi × Coconut fiber | 11.86 ± 0.50 | 115.7 ± 4.7 | 214.9 ± 22.1cde | 176.22 ± 1.28b |
| Pak choi × Peat moss | 13.94 ± 1.19 | 111.2 ± 27.8 | 261.9 ± 71.7bcd | 154.45 ± 8.26bc |
| Significance | ||||
| Microgreens species (M) | *** | *** | *** | *** |
| Substrate (S) | * | ** | *** | *** |
| M × S | ns | ns | * | * |
| Source of variance | Caffeic acid | Caffeic acid hexoside | Chlorogenic acid | p-Coumaroylquinic acid | Dicaffeoylquinic acid | Ferulic acid | Feruloyl quinic acid | Feruloylglycoside | Kaempferol-3-O-(caffeoyl) sophoroside-7-O-glucoside | Kaempferol-3-O-(feruloyl) sophoroside-7-O-glucoside | Kaempferol-3-O-(synapil) sophoroside-7-O-glucoside | Kaempferol-3-O-rutinoside | Luteolina-3-O-rutinoside | Quercetin sophoroside | Quercetin-3-O-(feruloyl) sophoroside-7-O-glucoside | Quercetin-3-O-sophoroside-7-O-glucoside | Quercetina-3-O-glucoside | Quercetina-3-O-glucuronide | Rosmarinic acid | Rutin | Total polyphenols |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (µg g−1 dw) | (mg g−1 dw) | |
| Microgreens species (M) | |||||||||||||||||||||
| Coriander | 125.84 ± 16.7a | 88.53 ± 4.1a | 6367 ± 549b | 64.85 ± 2.11a | 33.52 ± 1.22a | 20.55 ± 0.04a | 533.8 ± 20.6a | 93.14 ± 2.6c | nd | 319.0 ± 44.7a | 86.9 ± 14c | 566.5 ± 51a | 563 ± 51.3a | 83.99 ± 6.0a | nd | 4.89 ± 0.08c | 381.4 ± 23.9a | 791.1 ± 48a | 20.41 ± 0.08c | 5109 ± 367b | 15.25 ± 0.57ab |
| Kohlrabi | 40.34 ± 4.0b | 38.24 ± 4.5b | 6776 ± 516b | 22.99 ± 0.20c | 19.72 ± 0.03b | 19.55 ± 0.01c | 78.95 ± 3.3c | 154.79 ± 17.0b | 25.47 ± 1.44 b | 169.9 ± 6.3b | 291.7 ± 10b | 114.1 ± 3b | 164 ± 21.0b | 4.40 ± 0.1c | 10.79 ± 0.40b | 30.45 ± 1.08b | 222.0 ± 8.3b | nd | 20.64 ± 0.03b | 8052 ± 735a | 16.26 ± 0.78a |
| Pak choi | 32.71 ± 1.1c | 38.69 ± 3.1b | 9026 ± 837a | 25.90 ± 0.62b | 19.59 ± 0.03b | 20.24 ± 0.03b | 117.66 ± 8.4b | 314.81 ± 50.7a | 35.56 ± 2.25a | 113.4 ± 8.4c | 862.5 ± 40a | 128.4 ± 5b | 146 ± 8.3b | 10.80 ± 0.3b | 45.50 ± 2.93a | 71.93 ± 4.02a | 95.4 ± 6.0c | nd | 20.78 ± 0.05a | 3096 ± 365c | 14.22 ± 0.78b |
| Substrate (S) | |||||||||||||||||||||
| Agave fiber | 48.79 ± 7.2d | 55.07 ± 8.1b | 9947 ± 1095a | 40.40 ± 4.78ab | 23.75 ± 1.39b | 20.12 ± 0.11b | 281.26 ± 54.2a | 84.61 ± 4.6d | 16.68 ± 0.89c | 136.4 ± 5.3c | 363.4 ± 70c | 265.3 ± 43a | 238 ± 47.6c | 43.05 ± 12.6a | 18.55 ± 3.14e | 21.99 ± 3.37c | 216.3 ± 40.7bc | 754.1 ± 46bc | 20.52 ± 0.07b | 4027 ± 581d | 16.11 ± 0.71a |
| Capillary mat | 56.56 ± 4.7c | 49.05 ± 5.9bc | 8625 ± 780b | 42.01 ± 5.66a | 22.98 ± 1.14b | 20.08 ± 0.10b | 260.1 ± 48.2ab | 107.30 ± 9.6c | 32.90 ± 1.03b | 180.8 ± 18.1b | 512.5 ± 125a | 321.6 ± 80a | 340 ± 76.5ab | 26.88 ± 6.8b | 35.98 ± 6.7b | 33.97 ± 6.30b | 239.2 ± 33.1ab | 1057.9 ± 90a | 20.46 ± 0.07b | 5061 ± 390bc | 16.32 ± 0.39a |
| Cellulose sponge | 119.46 ± 31.6a | 49.87 ± 6.9bc | 8757 ± 579b | 32.47 ± 3.17d | 24.74 ± 1.70b | 20.05 ± 0.09b | 210.8 ± 42.2c | 272.90 ± 53.0b | 34.61 ± 1.06ab | 143.1 ± 13.6c | 446.2 ± 85b | 156.9 ± 20b | 223 ± 12.9c | 44.30 ± 13.1a | 22.35 ± 3.53d | 35.45 ± 6.10b | 274.8 ± 35.4a | 580.6 ± 66c | 20.61 ± 0.05b | 4584 ± 586cd | 15.65 ± 0.44a |
| Coconut fiber | 40.81 ± 7.1e | 43.28 ± 9.6c | 5290 ± 466c | 37.85 ± 5.06bc | 26.98 ± 2.94a | 20.22 ± 0.13a | 229.8 ± 60.8bc | 94.75 ± 3.4d | 30.94 ± 3.59b | 363.1 ± 71.1a | 362.0 ± 63c | 304.8 ± 73a | 260 ± 79.0bc | 28.56 ± 7.3b | 39.51 ± 9.02a | 43.40 ± 9.53a | 171.1 ± 17.1c | 617.5 ± 98c | 20.95 ± 0.09a | 5646 ± 543b | 13.24 ± 0.75b |
| Peat moss | 65.86 ± 4.4b | 78.49 ± 3.2a | 4331 ± 500d | 36.84 ± 5.21c | 22.95 ± 1.18b | 20.11 ± 0.11b | 235.5 ± 54.7bc | 378.32 ± 60.8a | 37.45 ± 4.35a | 180.3 ± 23.7 b | 384.5 ± 66c | 299.7 ± 74a | 394 ± 63.3a | 22.53 ± 5.7b | 24.34 ± 3.96c | 43.97 ± 8.71a | 263.4 ± 35.4ab | 945.6 ± 84ab | 20.51 ± 0.07b | 7776 ± 1377a | 14.91 ± 1.68a |
| M × S | |||||||||||||||||||||
| Coriander × Agave fiber | 86.11 ± 9.7b | 100.90 ± 5.7a | 8244 ± 1149de | 67.41 ± 2.82b | 31.82 ± 0.32bc | 20.53 ± 0.08bc | 587.9 ± 25.4a | 86.11 ± 2.4efg | nd | 116.2 ± 2.7f | 99.8 ± 15g | 505.2 ± 40b | 502 ± 39.8bc | 114.72 ± 7.9a | nd | 5.11 ± 0.17h | 430.1 ± 49.5a | 754.1 ± 46b | 20.18 ± 0.04h | 5169 ± 344cde | 16.94 ± 1.35bcd |
| Coriander × Capillary mat | 78.25 ± 0.5bc | 82.48 ± 2.6a | 4807 ± 762gh | 73.92 ± 4.53a | 29.45 ± 0.88c | 20.38 ± 0.10cd | 537.0 ± 19.9a | 84.89 ± 5.2fg | nd | 283.6 ± 1.9b | 55.0 ± 14g | 741.7 ± 108a | 738 ± 108.4a | 64.98 ± 5.4bc | nd | 4.80 ± 0.17h | 378.8 ± 42.1a | 1057.9 ± 90a | 20.18 ± 0.07h | 6741 ± 813bc | 15.80 ± 0.80bcdef |
| Coriander x Cellulose sponge | 303.39 ± 3.0a | 83.80 ± 9.9a | 9850 ± 679cd | 49.45 ± 3.88c | 34.60 ± 0.61b | 20.46 ± 0.04cd | 434.1 ± 53.1b | 82.59 ± 1.6fg | nd | 137.8 ± 2.7ef | 94.5 ± 26g | 250.4 ± 37c | 243 ± 37.3de | 117.17 ± 12.3a | nd | 5.28 ± 0.14h | 428.9 ± 68.1a | 580.6 ± 66c | 20.49 ± 0.13ef | 2800 ± 474fg | |
| Coriander × Coconut fiber | 76.75 ± 10.6bc | 89.20 ± 17.0a | 4113 ± 728h | 66.64 ± 3.34b | 41.98 ± 4.55a | 20.73 ± 0.10a | 560.7 ± 69.8a | 102.25 ± 3.5ef | nd | 769.1 ± 44.9a | 50.4 ± 16g | 644.4 ± 139ab | 641 ± 139.2ab | 69.25 ± 6.9b | nd | 4.66 ± 0.15h | 235.4 ± 18.0b | 617.5 ± 98c | 20.95 ± 0.24ab | 5427 ± 996cde | 13.55 ± 1.93efg |
| Coriander × Peat moss | 84.70 ± 1.4b | 86.24 ± 3.3a | 4822 ± 852gh | 66.81 ± 2.72b | 29.78 ± 0.47c | 20.66 ± 0.09ab | 549.5 ± 28.6a | 109.85 ± 5.6e | nd | 288.1 ± 5.8b | 134.8 ± 54g | 690.8 ± 103a | 689 ± 102.8a | 53.83 ± 6.2c | nd | 4.57 ± 0.18h | 434.0 ± 40.3a | 945.6 ± 84a | 20.2 ± 0.11gh | 5406 ± 530cde | 14.44 ± 0.43defg |
| Kohlrabi × Agave fiber | 23.73 ± 0.3gh | 28.60 ± 0.5cd | 5846 ± 164gh | 22.95 ± 0.11e | 19.72 ± 0.02d | 19.49 ± 0.00f | 76.37 ± 2.8de | 62.60 ± 1.1g | 15.96 ± 1.75e | 130.8 ± 4.1ef | 228.3 ± 11f | 127.8 ± 3c | 84.6 ± 18.5ef | 4.22 ± 0.1d | 8.13 ± 0.19h | 22.14 ± 0.71g | 150.5 ± 3.8cd | nd | 20.5 ± 0.03def | 6158 ± 11cd | 13.05 ± 0.13fg |
| Kohlrabi × Capillary mat | 59.64 ± 1.9d | 36.52 ± 1.0c | 11974 ± 601b | 24.77 ± 0.21de | 19.95 ± 0.02d | 19.54 ± 0.01f | 97.49 ± 4.3de | 159.55 ± 9.4d | 33.24 ± 0.80c | 138.8 ± 3.9ef | 258.4 ± 4ef | 100.2 ± 3c | 154 ± 3.7ef | 4.41 ± 0.1d | 13.89 ± 0.69f | 30.48 ± 1.64ef | 262.4 ± 8.1b | nd | 20.4 ± 0.04fgh | 4181 ± 128ef | 17.59 ± 0.59bc |
| Kohlrabi × Cellulose sponge | 30.18 ± 0.9efg | 29.33 ± 0.6cd | 5724 ± 200gh | 22.50 ± 0.16e | 19.81 ± 0.02d | 19.55 ± 0.01f | 68.37 ± 3.2de | 158.42 ± 3.7d | 33.94 ± 0.74c | 213.5 ± 3.5c | 343.2 ± 18de | 109.8 ± 5c | 207 ± 10.0def | 4.28 ± 0.1d | 10.94 ± 0.50g | 34.65 ± 1.42de | 245.5 ± 7.0b | nd | 20.7 ± 0.04bcdef | 7852 ± 202b | |
| Kohlrabi × Coconut fiber | 17.01 ± 0.5h | 12.96 ± 0.4d | 4176 ± 223h | 21.86 ± 0.27e | 19.49 ± 0.00d | 19.50 ± 0.00f | 55.75 ± 2.1e | 78.18 ± 2.8g | 19.56 ± 1.25e | 166.8 ± 2.9de | 357.3 ± 12d | 128.4 ± 4c | 28.1 ± 0.7f | 4.58 ± 0.1d | 9.65 ± 0.23gh | 28.82 ± 0.94f | 197.8 ± 4.9bc | nd | 20.7 ± 0.06bcde | 6601 ± 41bcd | 11.96 ± 0.26g |
| Kohlrabi × Peat moss | 71.12 ± 1.4c | 83.79 ± 1.4a | 6162 ± 134fg | 22.87 ± 0.13e | 19.65 ± 0.01d | 19.64 ± 0.01f | 96.79 ± 2.9de | 315.19 ± 15.9c | 24.66 ± 1.74d | 199.7 ± 7.4cd | 271.3 ± 5def | 104.3 ± 2c | 350 ± 10.6cd | 4.52 ± 0.0d | 11.34 ± 0.42g | 36.16 ± 1.80d | 253.8 ± 7.0b | nd | 20.8 ± 0.03bc | 15467 ± 660a | 23.53 ± 0.64a |
| Pak choi × Agave fiber | 36.53 ± 0.7ef | 35.70 ± 0.7c | 15750 ± 462a | 30.84 ± 0.22d | 19.71 ± 0.01d | 20.33 ± 0.03d | 179.5 ± 2.2c | 105.13 ± 5.0ef | 17.40 ± 0.50e | 162.3 ± 5.9de | 762.1 ± 31c | 163.0 ± 2c | 129 ± 11.2ef | 10.22 ± 0.1d | 28.96 ± 0.27e | 38.71 ± 1.45d | 68.2 ± 1.2d | nd | 20.8 ± 0.03bc | 753 ± 35h | 18.33 ± 0.49b |
| Pak choi × Capillary mat | 31.80 ± 0.4efg | 28.16 ± 0.6cd | 9093 ± 256cde | 27.33 ± 0.38de | 19.54 ± 0.00d | 20.30 ± 0.06d | 145.9 ± 5.1cd | 77.46 ± 1.6g | 32.56 ± 1.99c | 120.0 ± 11.6f | 1224.0 ± 44a | 122.8 ± 7c | 129 ± 18.3ef | 11.23 ± 0.4d | 58.07 ± 1.53b | 66.64 ± 4.02c | 76.4 ± 1.9d | nd | 20.8 ± 0.03bcd | 4261 ± 157ef | 15.57 ± 0.25cdef |
| Pak choi × Cellulose sponge | 24.80 ± 0.2fgh | 36.49 ± 6.6c | 10697 ± 296bc | 25.45 ± 0.50de | 19.81 ± 0.01d | 20.13 ± 0.01e | 129.8 ± 3.2cde | 577.69 ± 14.7b | 35.29 ± 2.05c | 78.0 ± 5.0g | 900.9 ± 65b | 110.7 ± 1c | 219 ± 7.6de | 11.46 ± 0.9d | 33.77 ± 1.61d | 66.42 ± 2.05c | 150.1 ± 6.1cd | nd | 20.7 ± 0.04cdef | 3100 ± 140fg | 16.26 ± 0.42bcde |
| Pak choi × Coconut fiber | 28.66 ± 0.6fgh | 27.68 ± 0.6cd | 7580 ± 253ef | 25.05 ± 0.46de | 19.49 ± 0.00d | 20.42 ± 0.02cd | 72.90 ± 1.7de | 103.81 ± 3.7ef | 42.32 ± 1.82b | 153.5 ± 6.7ef | 678.4 ± 16c | 141.5 ± 8c | 112 ± 4.0ef | 11.85 ± 0.6d | 69.36 ± 1.26a | 96.72 ± 3.75a | 80.0 ± 2.6d | nd | 21.2 ± 0.05a | 4912 ± 1308de | 14.20 ± 1.19defg |
| Pak choi × Peat moss | 41.77 ± 0.4e | 65.44 ± 6.4b | 2009 ± 87i | 20.83 ± 0.10e | 19.41 ± 0.00d | 20.03 ± 0.02e | 60.23 ± 2.0e | 709.93 ± 13.4a | 50.24 ± 3.84a | 53.0 ± 3.0g | 747.3 ± 20c | 103.9 ± 1c | 144 ± 2.6ef | 9.25 ± 0.6d | 37.34 ± 1.11c | 91.17 ± 1.51b | 102.3 ± 10.3d | nd | 20.4 ± 0.04efg | 2454 ± 56g | 6.76 ± 0.13h |
| Significance | |||||||||||||||||||||
| Microgreens species (M) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Substrate (S) | *** | *** | *** | *** | *** | ** | ** | *** | *** | *** | *** | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| M × S | *** | *** | *** | *** | *** | *** | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ** | *** | *** | *** | *** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Palladino, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Phenolic Constitution, Phytochemical and Macronutrient Content in Three Species of Microgreens as Modulated by Natural Fiber and Synthetic Substrates. Antioxidants 2020, 9, 252. https://doi.org/10.3390/antiox9030252
Kyriacou MC, El-Nakhel C, Pannico A, Graziani G, Soteriou GA, Giordano M, Palladino M, Ritieni A, De Pascale S, Rouphael Y. Phenolic Constitution, Phytochemical and Macronutrient Content in Three Species of Microgreens as Modulated by Natural Fiber and Synthetic Substrates. Antioxidants. 2020; 9(3):252. https://doi.org/10.3390/antiox9030252
Chicago/Turabian StyleKyriacou, Marios C., Christophe El-Nakhel, Antonio Pannico, Giulia Graziani, Georgios A. Soteriou, Maria Giordano, Mario Palladino, Alberto Ritieni, Stefania De Pascale, and Youssef Rouphael. 2020. "Phenolic Constitution, Phytochemical and Macronutrient Content in Three Species of Microgreens as Modulated by Natural Fiber and Synthetic Substrates" Antioxidants 9, no. 3: 252. https://doi.org/10.3390/antiox9030252
APA StyleKyriacou, M. C., El-Nakhel, C., Pannico, A., Graziani, G., Soteriou, G. A., Giordano, M., Palladino, M., Ritieni, A., De Pascale, S., & Rouphael, Y. (2020). Phenolic Constitution, Phytochemical and Macronutrient Content in Three Species of Microgreens as Modulated by Natural Fiber and Synthetic Substrates. Antioxidants, 9(3), 252. https://doi.org/10.3390/antiox9030252