
Advances in Toxoplasma gondii Vaccines: Current Strategies and Challenges for Vaccine Development


Abstract
:1. Introduction
2. Vaccine Platforms against T. gondii
2.1. DNA and Vectored Vaccines
2.2. Protein and Recombinant Subunit Vaccines
2.3. Nanoparticles, Virus-Like Particle Vaccines
2.4. Live-Attenuated Vaccines
2.5. Carbohydrate Vaccines
3. Limitations and Challenges to Vaccine Design
4. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Halonen, S.K.; Weiss, L.M. Toxoplasmosis. Handb. Clin. Neurol. 2013, 114, 125–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montazeri, M.; Mehrzadi, S.; Sharif, M.; Sarvi, S.; Tanzifi, A.; Aghayan, S.A.; Daryani, A. Drug Resistance in Toxoplasma gondii. Front. Microbiol. 2018, 9, 2587. [Google Scholar] [CrossRef] [Green Version]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of Toxoplasmosis: Historical Perspective, Animal Models, and Current Clinical Practice. Clin. Microbiol. Rev. 2018, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.L.; Zhang, N.Z.; Li, T.T.; He, J.J.; Elsheikha, H.M.; Zhu, X.Q. Advances in the Development of Anti-Toxoplasma gondii Vaccines: Challenges, Opportunities, and Perspectives. Trends Parasitol. 2019, 35, 239–253. [Google Scholar] [CrossRef]
- Gallant, J. Get Rich Quick with Old Generic Drugs! The Pyrimethamine Pricing Scandal. Open Forum Infect. Dis. 2015, 2, ofv177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rémy, V.; Zöllner, Y.; Heckmann, U. Vaccination: The cornerstone of an efficient healthcare system. J. Mark. Access Health Policy 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, J.P. Toxoplasmosis in sheep—The last 20 years. Vet. Parasitol. 2009, 163, 1–14. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, H. Moving towards improved vaccines for Toxoplasma gondii. Expert Opin. Biol. Ther. 2018, 18, 273–280. [Google Scholar] [CrossRef]
- Sonaimuthu, P.; Ching, X.T.; Fong, M.Y.; Kalyanasundaram, R.; Lau, Y.L. Induction of Protective Immunity against Toxoplasmosis in BALB/c Mice Vaccinated with Toxoplasma gondii Rhoptry-1. Front. Microbiol. 2016, 7, 808. [Google Scholar] [CrossRef]
- Alizadeh, P.; Ahmadpour, E.; Daryani, A.; Kazemi, T.; Spotin, A.; Mahami-Oskouei, M.; Flynn, R.J.; Azadi, Y.; Rajabi, S.; Sandoghchian, S. IL-17 and IL-22 elicited by a DNA vaccine encoding ROP13 associated with protection against Toxoplasma gondii in BALB/c mice. J. Cell. Physiol. 2019, 234, 10782–10788. [Google Scholar] [CrossRef]
- Li, X.Z.; Lv, L.; Zhang, X.; Anchang, K.Y.; Abdullahi, A.Y.; Tu, L.; Wang, X.; Xia, L.; Zhang, X.X.; Feng, W.; et al. Recombinant canine adenovirus type-2 expressing TgROP16 provides partial protection against acute Toxoplasma gondii infection in mice. Infect. Genet. Evol. 2016, 45, 447–453. [Google Scholar] [CrossRef]
- Rashid, I.; Moiré, N.; Héraut, B.; Dimier-Poisson, I.; Mévélec, M.N. Enhancement of the protective efficacy of a ROP18 vaccine against chronic toxoplasmosis by nasal route. Med. Microbiol. Immunol. 2017, 206, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.B.; Zhou, D.H.; Zou, Y.; Chen, K.; Liu, Q.; Wang, J.L.; Zhu, X.Q.; Zhao, G.H. Vaccination with a DNA vaccine encoding Toxoplasma gondii ROP54 induces protective immunity against toxoplasmosis in mice. Acta Trop. 2017, 176, 427–432. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.; Wang, M.; Xie, Q.; Li, P.; Zuo, S.; Kong, L.; Wang, C.; Wang, S. Immune Protection of Rhoptry Protein 21 (ROP21) of Toxoplasma gondii as a DNA Vaccine Against Toxoplasmosis. Front. Microbiol. 2018, 9, 909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ching, X.T.; Fong, M.Y.; Lau, Y.L. Evaluation of the Protective Effect of Deoxyribonucleic Acid Vaccines Encoding Granule Antigen 2 and 5 Against Acute Toxoplasmosis in BALB/c Mice. Am. J. Trop. Med. Hyg. 2017, 96, 1441–1447. [Google Scholar] [CrossRef]
- Hu, L.Y.; Zhang, N.Z.; Zhang, F.K.; Wang, M.; Gao, Q.; Wang, J.L.; Zhu, X.Q. Resistance to Chronic Toxoplasma gondii Infection Induced by a DNA Vaccine Expressing GRA16. Biomed. Res. Int. 2017, 2017, 1295038. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Lou, D.; Ding, J.; Zhuo, X.; Ding, H.; Kong, Q.; Lu, S. GRA24-Based DNA Vaccine Prolongs Survival in Mice Challenged with a Virulent Toxoplasma gondii Strain. Front. Immunol. 2019, 10, 418. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.Y.; Lu, J.; Zhang, N.Z.; Elsheikha, H.M.; Hou, J.L.; Guo, H.T.; Zhu, X.Q. Immunization with plasmid DNA expressing Heat Shock Protein 40 confers prophylactic protection against chronic Toxoplasma gondii infection in Kunming mice. Parasite 2018, 25, 37. [Google Scholar] [CrossRef]
- Zheng, B.; Ding, J.; Lou, D.; Tong, Q.; Zhuo, X.; Ding, H.; Kong, Q.; Lu, S. The Virulence-Related MYR1 Protein of Toxoplasma gondii as a Novel DNA Vaccine against Toxoplasmosis in Mice. Front. Microbiol. 2019, 10, 734. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.Y.; Chen, K.; Wang, J.L.; Yang, B.; Zhu, X.Q. Evaluation of protective immunity induced by recombinant calcium-dependent protein kinase 1 (TgCDPK1) protein against acute toxoplasmosis in mice. Microb. Pathog. 2019, 133, 103560. [Google Scholar] [CrossRef]
- Chen, K.; Wang, J.L.; Huang, S.Y.; Yang, W.B.; Zhu, W.N.; Zhu, X.Q. Immune responses and protection after DNA vaccination against Toxoplasma gondii calcium-dependent protein kinase 2 (TgCDPK2). Parasite 2017, 24, 41. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.Z.; Gao, Q.; Wang, M.; Hou, J.L.; Zhang, F.K.; Hu, L.Y.; Zhu, X.Q. Protective Efficacy Against Acute and Chronic Toxoplasma gondii Infection Induced by Immunization with the DNA Vaccine TgDOC2C. Front. Microbiol. 2018, 9, 2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadpour, E.; Sarvi, S.; Soteh, M.B.H.; Sharif, M.; Rahimi, M.T.; Valadan, R.; Tehrani, M.; Khalilian, A.; Montazeri, M.; Daryani, A. Evaluation of the immune response in BALB/c mice induced by a novel DNA vaccine expressing GRA14 against Toxoplasma gondii. Parasite Immunol. 2017, 39. [Google Scholar] [CrossRef]
- Ahmadpour, E.; Sarvi, S.; Soteh, M.B.H.; Sharif, M.; Rahimi, M.T.; Valadan, R.; Tehrani, M.; Khalilian, A.; Montazeri, M.; Fasihi-Ramandi, M.; et al. Enhancing immune responses to a DNA vaccine encoding Toxoplasma gondii GRA14 by calcium phosphate nanoparticles as an adjuvant. Immunol. Lett. 2017, 185, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Zhou, J.; Zhao, Y.H.; Wang, L. DNA vaccine ROP29 from Toxoplasma gondii containing R848 enhances protective immunity in mice. Parasite Immunol. 2018, 40, e12578. [Google Scholar] [CrossRef]
- Foroutan, M.; Barati, M.; Ghaffarifar, F. Enhancing immune responses by a novel multi-epitope ROP8 DNA vaccine plus interleukin-12 plasmid as a genetic adjuvant against acute Toxoplasma gondii infection in BALB/c mice. Microb. Pathog. 2020, 147, 104435. [Google Scholar] [CrossRef]
- Mavi, S.A.; Modarressi, M.H.; Mohebali, M.; Shojaee, S.; Zeraati, H.; Teimouri, A.; Keshavarz, H. Assessment of the immunogenicity and protective efficiency of a novel dual-promoter DNA vaccine, harboring SAG1 and GRA7 genes, from RH strain of Toxoplasma gondii in BALB/c mice. Infect. Drug Resist. 2019, 12, 2519–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.C.; He, Y.; Liu, J.F.; Chen, J. Adjuvantic cytokine IL-33 improves the protective immunity of cocktailed DNA vaccine of ROP5 and ROP18 against Toxoplasma gondii infection in mice. Parasite 2020, 27, 26. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Li, J.; Xiao, T.; Huang, X.D.; Wang, L.J.; Huang, B.C.; Yin, K.; Liu, G.Z.; Xu, C.; Wei, Q.K. Protective immunity induced by a DNA vaccine cocktail expressing TgSAG1, TgROP2, and the genetic adjuvant HBsAg against Toxoplasma gondii infection. Microb. Pathog. 2020, 147, 104441. [Google Scholar] [CrossRef]
- Pagheh, A.S.; Daryani, A.; Alizadeh, P.; Hassannia, H.; Oliveira, S.M.R.; Kazemi, T.; Rezaei, F.; Pereira, M.L.; Ahmadpour, E. Protective effect of a DNA vaccine cocktail encoding ROP13 and GRA14 with Alum nano-adjuvant against Toxoplasma gondii infection in mice. Int. J. Biochem. Cell Biol. 2021, 132, 105920. [Google Scholar] [CrossRef]
- Zhang, N.Z.; Gao, Q.; Wang, M.; Elsheikha, H.M.; Wang, B.; Wang, J.L.; Zhang, F.K.; Hu, L.Y.; Zhu, X.Q. Immunization with a DNA Vaccine Cocktail Encoding TgPF, TgROP16, TgROP18, TgMIC6, and TgCDPK3 Genes Protects Mice Against Chronic Toxoplasmosis. Front. Immunol. 2018, 9, 1505. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.P.; Liu, W.G.; Xu, Q.M.; Zhu, X.Q.; Chen, J. Evaluation of immune protection against Toxoplasma gondii infection in mice induced by a multi-antigenic DNA vaccine containing TgGRA24, TgGRA25 and TgMIC6. Parasite 2019, 26, 58. [Google Scholar] [CrossRef] [Green Version]
- Chahal, J.S.; Khan, O.F.; Cooper, C.L.; McPartlan, J.S.; Tsosie, J.K.; Tilley, L.D.; Sidik, S.M.; Lourido, S.; Langer, R.; Bavari, S.; et al. Dendrimer-RNA nanoparticles generate protective immunity against lethal Ebola, H1N1 influenza, and Toxoplasma gondii challenges with a single dose. Proc. Natl. Acad. Sci. USA 2016, 113, E4133–E4142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Feng, F.; Lv, J.; Xie, Z.; Chen, J.; Zhang, L.; Li, W. Major Immunodominant Region of Hepatitis B Virus Core Antigen as a Delivery Vector to Improve the Immunogenicity of the Fusion Antigen ROP2-SAG1 Multiepitope from Toxoplasma gondii in Mice. Viral Immunol. 2017, 30, 508–515. [Google Scholar] [CrossRef]
- Zhang, D.; Jiang, N.; Chen, Q. Vaccination with recombinant adenoviruses expressing Toxoplasma gondii MIC3, ROP9, and SAG2 provide protective immunity against acute toxoplasmosis in mice. Vaccine 2019, 37, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Hu, Y.; Hua, Q.; Luo, F.; Xie, G.; Li, X.; Lin, J.; Wan, Y.; Ren, S.; Pan, C.; et al. Protective immune response in mice induced by a suicidal DNA vaccine encoding NTPase-II gene of Toxoplasma gondii. Acta Trop. 2017, 166, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Zheng, L.; Hu, Y.; Liu, S.; Wang, Y.; Xiong, Z.; Hu, X.; Tan, F. Induction of Protective Immunity against Toxoplasma gondii in Mice by Nucleoside Triphosphate Hydrolase-II (NTPase-II) Self-amplifying RNA Vaccine Encapsulated in Lipid Nanoparticle (LNP). Front. Microbiol. 2017, 8, 605. [Google Scholar] [CrossRef]
- Maeta, M.; Miura, N.; Tanaka, H.; Nakamura, T.; Kawanishi, R.; Nishikawa, Y.; Asano, K.; Tanaka, M.; Tamagawa, S.; Nakai, Y.; et al. Vitamin E Scaffolds of pH-Responsive Lipid Nanoparticles as DNA Vaccines in Cancer and Protozoan Infection. Mol. Pharm. 2020, 17, 1237–1247. [Google Scholar] [CrossRef]
- Wang, T.; Yin, H.; Li, Y.; Zhao, L.; Sun, X.; Cong, H. Vaccination with recombinant adenovirus expressing multi-stage antigens of Toxoplasma gondii by the mucosal route induces higher systemic cellular and local mucosal immune responses than with other vaccination routes. Parasite 2017, 24, 12. [Google Scholar] [CrossRef] [Green Version]
- El Bissati, K.; Chentoufi, A.A.; Krishack, P.A.; Zhou, Y.; Woods, S.; Dubey, J.P.; Vang, L.; Lykins, J.; Broderick, K.E.; Mui, E.; et al. Adjuvanted multi-epitope vaccines protect HLA-A*11:01 transgenic mice against Toxoplasma gondii. JCI Insight 2016, 1, e85955. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Ding, J.; Chen, X.; Yu, H.; Lou, D.; Tong, Q.; Kong, Q.; Lu, S. Immuno-Efficacy of a T. gondii Secreted Protein with an Altered Thrombospondin Repeat (TgSPATR) As a Novel DNA Vaccine Candidate against Acute Toxoplasmosis in BALB/c Mice. Front. Microbiol. 2017, 8, 216. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, L. SAG4 DNA and Peptide Vaccination Provides Partial Protection against T. gondii Infection in BALB/c Mice. Front. Microbiol. 2017, 8, 1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagheh, A.S.; Sarvi, S.; Gholami, S.; Asgarian-Omran, H.; Valadan, R.; Hassannia, H.; Ahmadpour, E.; Fasihi-Ramandie, M.; Dodangeh, S.; Hosseni-Khah, Z.; et al. Protective efficacy induced by DNA prime and recombinant protein boost vaccination with Toxoplasma gondii GRA14 in mice. Microb. Pathog. 2019, 134, 103601. [Google Scholar] [CrossRef]
- Vartak, A.; Sucheck, S.J. Recent Advances in Subunit Vaccine Carriers. Vaccines 2016, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Costa, A.; Zorgi, N.E.; do Nascimento, N.; Galisteo, A.J.J.; de Andrade, H.F.J. Gamma irradiation of Toxoplasma gondii protein extract improve immune response and protection in mice models. Biomed. Pharmacother. 2018, 106, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, A.; Nascimento, N.D.; Galisteo, A.J.; Passos, A.B.D.D.; de Andrade, H.F. Radiation effects on Toxoplasma antigens: Different immune responses of irradiated intact tachyzoites or soluble antigens in experimental mice models. Int. J. Radiat. Biol. 2020, 96, 697–706. [Google Scholar] [CrossRef]
- Rahman, M.; Devriendt, B.; Algaba, I.G.; Verhaegen, B.; Dorny, P.; Dierick, K.; Cox, E. QuilA-Adjuvanted T. gondii Lysate Antigens Trigger Robust Antibody and IFNγ+ T Cell Responses in Pigs Leading to Reduction in Parasite DNA in Tissues Upon Challenge Infection. Front. Immunol. 2019, 10, 2223. [Google Scholar] [CrossRef]
- Roozbehani, M.; Falak, R.; Mohammadi, M.; Hemphill, A.; Razmjou, E.; Meamar, A.R.; Masoori, L.; Khoshmirsafa, M.; Moradi, M.; Gharavi, M.J. Characterization of a multi-epitope peptide with selective MHC-binding capabilities encapsulated in PLGA nanoparticles as a novel vaccine candidate against Toxoplasma gondii infection. Vaccine 2018, 36, 6124–6132. [Google Scholar] [CrossRef]
- Guo, J.; Sun, X.; Yin, H.; Wang, T.; Li, Y.; Zhou, C.; Zhou, H.; He, S.; Cong, H. Chitosan Microsphere Used as an Effective System to Deliver a Linked Antigenic Peptides Vaccine Protect Mice Against Acute and Chronic Toxoplasmosis. Front. Cell. Infect. Microbiol. 2018, 8, 163. [Google Scholar] [CrossRef]
- Fereig, R.M.; Kuroda, Y.; Terkawi, M.A.; Mahmoud, M.E.; Nishikawa, Y. Immunization with Toxoplasma gondii peroxiredoxin 1 induces protective immunity against toxoplasmosis in mice. PLoS ONE 2017, 12, e0176324. [Google Scholar] [CrossRef]
- Liu, K.; Wen, H.; Cai, H.; Wu, M.; An, R.; Chu, D.; Yu, L.; Shen, J.; Chen, L.; Du, J. Protective Effect Against Toxoplasmosis in BALB/c Mice Vaccinated with Toxoplasma gondii Macrophage Migration Inhibitory Factor. Front. Microbiol. 2019, 10, 813. [Google Scholar] [CrossRef]
- Liu, Z.; Yuan, F.; Yang, Y.; Yin, L.; Liu, Y.; Wang, Y.; Zheng, K.; Cao, J. Partial protective immunity against toxoplasmosis in mice elicited by recombinant Toxoplasma gondii malate dehydrogenase. Vaccine 2016, 34, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yin, L.; Li, Y.; Yuan, F.; Zhang, X.; Ma, J.; Liu, H.; Wang, Y.; Zheng, K.; Cao, J. Intranasal immunization with recombinant Toxoplasma gondii actin depolymerizing factor confers protective efficacy against toxoplasmosis in mice. BMC Immunol. 2016, 17, 37. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; An, R.; Chen, Y.; Chen, T.; Wen, H.; Yan, Q.; Shen, J.; Chen, L.; Du, J. Vaccination with recombinant Toxoplasma gondii CDPK3 induces protective immunity against experimental toxoplasmosis. Acta Trop. 2019, 199, 105148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, Y.; Li, H.; Song, X.; Ma, Z.; Lu, H.; Liu, S.; Zhao, Y.; Tan, M.; Wang, S.; et al. Identification of Toxoplasma gondii Tyrosine Hydroxylase (TH) Activity and Molecular Immunoprotection against Toxoplasmosis. Vaccines 2020, 8, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.Y.; Guo, H.T.; Calderón-Mantilla, G.; He, J.J.; Wang, J.L.; Bonev, B.B.; Zhu, X.Q.; Elsheikha, H.M. Immunostimulatory efficacy and protective potential of putative TgERK7 protein in mice experimentally infected by Toxoplasma gondii. Int. J. Med. Microbiol. 2020, 310, 151432. [Google Scholar] [CrossRef] [PubMed]
- Dimier-Poisson, I.; Carpentier, R.; N’Guyen, T.T.; Dahmani, F.; Ducournau, C.; Betbeder, D. Porous nanoparticles as delivery system of complex antigens for an effective vaccine against acute and chronic Toxoplasma gondii infection. Biomaterials 2015, 50, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhou, T.; Luo, Y.; Dong, L.; Li, C.; Liu, J.; Luo, J.; Yan, R.; Xu, L.; Song, X.; et al. Modulation Effects of Toxoplasma gondii Histone H2A1 on Murine Macrophages and Encapsulation with Polymer as a Vaccine Candidate. Vaccines 2020, 8, 731. [Google Scholar] [CrossRef]
- Chuang, S.C.; Chung, Y.C.; Yang, C.D. Protective immunity against toxoplasmosis in mice induced by single-dose immunization with rSAG1/2 protein released from poly(lactide-co-glycolide) microparticles. Parasite 2017, 24, 5. [Google Scholar] [CrossRef] [Green Version]
- Czarnewski, P.; Araújo, E.C.B.; Oliveira, M.C.; Mineo, T.W.P.; Silva, N.M. Recombinant TgHSP70 Immunization Protects against Toxoplasma gondii Brain Cyst Formation by Enhancing Inducible Nitric Oxide Expression. Front. Cell. Infect. Microbiol. 2017, 7, 142. [Google Scholar] [CrossRef] [Green Version]
- Ching, X.T.; Fong, M.Y.; Lau, Y.L. Evaluation of Immunoprotection Conferred by the Subunit Vaccines of GRA2 and GRA5 against Acute Toxoplasmosis in BALB/c Mice. Front. Microbiol. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Bastos, L.M.; Macêdo, A.G.; Silva, M.V.; Santiago, F.M.; Ramos, E.L.; Santos, F.A.; Pirovani, C.P.; Goulart, L.R.; Mineo, T.W.; Mineo, J.R. Toxoplasma gondii-Derived Synthetic Peptides Containing B- and T-Cell Epitopes from GRA2 Protein Are Able to Enhance Mice Survival in a Model of Experimental Toxoplasmosis. Front. Cell. Infect. Microbiol. 2016, 6, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatkowska, J.; Dzitko, K.; Ferra, B.T.; Holec-Gąsior, L.; Kawka, M.; Dziadek, B. The Immunogenic and Immunoprotective Activities of Recombinant Chimeric T. gondii Proteins Containing AMA1 Antigen Fragments. Vaccines 2020, 8, 724. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Song, X.; Kong, X.; Zhang, N.; Qu, S.; Zhu, W.; Yang, Y.; Wang, Q. Immunization with Toxoplasma gondii aspartic protease 3 increases survival time of infected mice. Acta Trop. 2017, 171, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.J.; Xiao, T.; Xu, C.; Li, J.; Liu, G.Z.; Yin, K.; Cui, Y.; Wei, Q.K.; Huang, B.C.; Sun, H. Protective immune response against Toxoplasma gondii elicited by a novel yeast-based vaccine with microneme protein 16. Vaccine 2018, 36, 3943–3948. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, E.F.; Corigliano, M.G.; Albarracín, R.M.; Sander, V.A.; Legarralde, A.; Bengoa-Luoni, S.A.; Clemente, M. Plant Hsp90 is a novel adjuvant that elicits a strong humoral and cellular immune response against B- and T-cell epitopes of a Toxoplasma gondii SAG1 peptide. Parasites Vectors 2019, 12, 140. [Google Scholar] [CrossRef] [PubMed]
- Lakhrif, Z.; Moreau, A.; Hérault, B.; Di-Tommaso, A.; Juste, M.; Moiré, N.; Dimier-Poisson, I.; Mévélec, M.N.; Aubrey, N. Targeted Delivery of Toxoplasma gondii Antigens to Dendritic Cells Promote Immunogenicity and Protective Efficiency against Toxoplasmosis. Front. Immunol. 2018, 9, 317. [Google Scholar] [CrossRef] [Green Version]
- Arcon, N.; Picchio, M.S.; Fenoy, I.M.; Moretta, R.E.; Soto, A.S.; Sibilia, M.D.P.; Sánchez, V.R.; Prato, C.A.; Tribulatti, M.V.; Goldman, A.; et al. Synergistic effect of GRA7 and profilin proteins in vaccination against chronic Toxoplasma gondii infection. Vaccine 2021, 39, 933–942. [Google Scholar] [CrossRef]
- Lung, P.; Yang, J.; Li, Q. Nanoparticle formulated vaccines: Opportunities and challenges. Nanoscale 2020, 12, 5746–5763. [Google Scholar] [CrossRef]
- El Bissati, K.; Zhou, Y.; Paulillo, S.M.; Raman, S.K.; Karch, C.P.; Roberts, C.W.; Lanar, D.E.; Reed, S.; Fox, C.; Carter, D.; et al. Protein nanovaccine confers robust immunity against Toxoplasma. NPJ Vaccines 2017, 2, 24. [Google Scholar] [CrossRef] [Green Version]
- El Bissati, K.; Zhou, Y.; Paulillo, S.M.; Raman, S.K.; Karch, C.P.; Reed, S.; Estes, A.; Estes, A.; Lykins, J.; Burkhard, P.; et al. Engineering and characterization of a novel Self Assembling Protein for Toxoplasma peptide vaccine in HLA-A*11:01, HLA-A*02:01 and HLA-B*07:02 transgenic mice. Sci. Rep. 2020, 10, 16984. [Google Scholar] [CrossRef]
- Ducournau, C.; Nguyen, T.T.; Carpentier, R.; Lantier, I.; Germon, S.; Précausta, F.; Pisella, P.J.; Leroux, H.; Van Langendonck, N.; Betbeder, D.; et al. Synthetic parasites: A successful mucosal nanoparticle vaccine against Toxoplasma congenital infection in mice. Futur. Microbiol. 2017, 12, 393–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducournau, C.; Moiré, N.; Carpentier, R.; Cantin, P.; Herkt, C.; Lantier, I.; Betbeder, D.; Dimier-Poisson, I. Effective Nanoparticle-Based Nasal Vaccine Against Latent and Congenital Toxoplasmosis in Sheep. Front. Immunol. 2020, 11, 2183. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Kato, K. Inorganic nanoparticles kill Toxoplasma gondii via changes in redox status and mitochondrial membrane potential. Int. J. Nanomed. 2017, 12, 1647–1661. [Google Scholar] [CrossRef] [Green Version]
- Adeyemi, O.S.; Molefe, N.I.; Awakan, O.J.; Nwonuma, C.O.; Alejolowo, O.O.; Olaolu, T.; Maimako, R.F.; Suganuma, K.; Han, Y.; Kato, K. Metal nanoparticles restrict the growth of protozoan parasites. Artif. Cells Nanomed. Biotechnol. 2018, 46, S86–S94. [Google Scholar] [CrossRef] [Green Version]
- Costa, I.N.; Ribeiro, M.; Franco, P.S.; da Silva, R.J.; de Araújo, T.E.; Milián, I.C.B.; Luz, L.C.; Guirelli, P.M.; Nakazato, G.; Mineo, J.R.; et al. Biogenic Silver Nanoparticles Can Control Toxoplasma gondii Infection in Both Human Trophoblast Cells and Villous Explants. Front. Microbiol. 2020, 11, 623947. [Google Scholar] [CrossRef]
- Kushnir, N.; Streatfield, S.J.; Yusibov, V. Virus-like particles as a highly efficient vaccine platform: Diversity of targets and production systems and advances in clinical development. Vaccine 2012, 31, 58–83. [Google Scholar] [CrossRef]
- Chu, K.B.; Quan, F.S. Virus-Like Particle Vaccines against Respiratory Viruses and Protozoan Parasites. Curr. Top. Microbiol. Immunol. 2021. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, A.R.; Lee, S.H.; Quan, F.S. Cross-protection induced by Toxoplasma gondii virus-like particle vaccine upon intraperitoneal route challenge. Acta Trop. 2016, 164, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lee, S.H.; Kim, A.R.; Quan, F.S. Virus-Like Nanoparticle Vaccine Confers Protection against Toxoplasma gondii. PLoS ONE 2016, 11, e0161231. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.J.; Chu, K.B.; Lee, S.H.; Kim, M.J.; Park, H.; Jin, H.; Moon, E.K.; Quan, F.S. Toxoplasma gondii virus-like particle vaccination alleviates inflammatory response in the brain upon T. gondii infection. Parasite Immunol. 2020, 42, e12716. [Google Scholar] [CrossRef]
- Kang, H.J.; Lee, S.H.; Kim, M.J.; Chu, K.B.; Lee, D.H.; Chopra, M.; Choi, H.J.; Park, H.; Jin, H.; Quan, F.S. Influenza Virus-Like Particles Presenting both Toxoplasma gondii ROP4 and ROP13 Enhance Protection against T. gondii Infection. Pharmaceutics 2019, 11, 342. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Chu, K.B.; Kang, H.J.; Quan, F.S. Virus-like particles containing multiple antigenic proteins of Toxoplasma gondii induce memory T cell and B cell responses. PLoS ONE 2019, 14, e0220865. [Google Scholar] [CrossRef]
- Kim, M.J.; Lee, S.H.; Kang, H.J.; Chu, K.B.; Park, H.; Jin, H.; Moon, E.K.; Kim, S.S.; Quan, F.S. Virus-like particle vaccine displaying Toxoplasma gondii apical membrane antigen 1 induces protection against T. gondii ME49 infection in mice. Microb. Pathog. 2020, 142, 104090. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Chu, K.B.; Kim, M.J.; Lee, S.H.; Park, H.; Jin, H.; Moon, E.K.; Quan, F.S. Protective immunity induced by CpG ODN-adjuvanted virus-like particles containing Toxoplasma gondii proteins. Parasite Immunol. 2021, 43, e12799. [Google Scholar] [CrossRef]
- Kang, H.J.; Chu, K.B.; Kim, M.J.; Park, H.; Jin, H.; Lee, S.H.; Moon, E.K.; Quan, F.S. Evaluation of CpG-ODN-Adjuvanted Toxoplasma gondii Virus-Like Particle Vaccine upon One, Two, and Three Immunizations. Pharmaceutics 2020, 12, 989. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, H.J.; Lee, D.H.; Kang, S.M.; Quan, F.S. Virus-like particle vaccines expressing Toxoplasma gondii rhoptry protein 18 and microneme protein 8 provide enhanced protection. Vaccine 2018, 36, 5692–5700. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhou, A.; Sun, X.; Sha, W.; Ai, K.; Pan, G.; Zhou, C.; Zhou, H.; Cong, H.; He, S. Immunogenicity of a Virus-Like-Particle Vaccine Containing Multiple Antigenic Epitopes of Toxoplasma gondii Against Acute and Chronic Toxoplasmosis in Mice. Front. Immunol. 2019, 10, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Kim, A.R.; Lee, D.H.; Rubino, I.; Choi, H.J.; Quan, F.S. Protection induced by virus-like particles containing Toxoplasma gondii microneme protein 8 against highly virulent RH strain of Toxoplasma gondii infection. PLoS ONE 2017, 12, e0175644. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, H.J.; Lee, D.H.; Quan, F.S. Protective Immunity Induced by Incorporating Multiple Antigenic Proteins of Toxoplasma gondii Into Influenza Virus-Like Particles. Front. Immunol. 2018, 9, 3073. [Google Scholar] [CrossRef]
- Zhuo, X.; Du, K.; Ding, H.; Lou, D.; Zheng, B.; Lu, S. A Carbamoyl Phosphate Synthetase II (CPSII) Deletion Mutant of Toxoplasma gondii Induces Partial Protective Immunity in Mice. Front. Microbiol. 2020, 11, 616688. [Google Scholar] [CrossRef]
- Yang, J.; Yang, C.; Qian, J.; Li, F.; Zhao, J.; Fang, R. Toxoplasma gondii α-amylase deletion mutant is a promising vaccine against acute and chronic toxoplasmosis. Microb. Biotechnol. 2020, 13, 2057–2069. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Tang, D.; Yang, C.; Yang, J.; Fang, R. Toxoplasma gondii ADSL Knockout Provides Excellent Immune Protection against a Variety of Strains. Vaccines 2020, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.B.; Wang, J.L.; Gui, Q.; Zou, Y.; Chen, K.; Liu, Q.; Liang, Q.L.; Zhu, X.Q.; Zhou, D.H. Immunization With a Live-Attenuated RH:ΔNPT1 Strain of Toxoplasma gondii Induces Strong Protective Immunity Against Toxoplasmosis in Mice. Front. Microbiol. 2019, 10, 1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.L.; Elsheikha, H.M.; Zhu, W.N.; Chen, K.; Li, T.T.; Yue, D.M.; Zhang, X.X.; Huang, S.Y.; Zhu, X.Q. Immunization with Toxoplasma gondii GRA17 Deletion Mutant Induces Partial Protection and Survival in Challenged Mice. Front. Immunol. 2017, 8, 730. [Google Scholar] [CrossRef] [Green Version]
- Lagal, V.; Dinis, M.; Cannella, D.; Bargieri, D.; Gonzalez, V.; Andenmatten, N.; Meissner, M.; Tardieux, I. AMA1-Deficient Toxoplasma gondii Parasites Transiently Colonize Mice and Trigger an Innate Immune Response that Leads to Long-Lasting Protective Immunity. Infect. Immun. 2015, 83, 2475–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, B.A.; Bzik, D.J. Nonreplicating, cyst-defective type II Toxoplasma gondii Vaccine Strains Stimulate Protective Immunity against Acute and Chronic Infection. Infect. Immun. 2015, 83, 2148–2155. [Google Scholar] [CrossRef] [Green Version]
- Xia, N.; Zhou, T.; Liang, X.; Ye, S.; Zhao, P.; Yang, J.; Zhou, Y.; Zhao, J.; Shen, B. A Lactate Fermentation Mutant of Toxoplasma Stimulates Protective Immunity against Acute and Chronic Toxoplasmosis. Front. Immunol. 2018, 9, 1814. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.L.; Li, T.T.; Elsheikha, H.M.; Chen, K.; Cong, W.; Yang, W.B.; Bai, M.J.; Huang, S.Y.; Zhu, X.Q. Live Attenuated Pru:Δcdpk2 Strain of Toxoplasma gondii Protects Against Acute, Chronic, and Congenital Toxoplasmosis. J. Infect. Dis. 2018, 218, 768–777. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.L.; Liang, Q.L.; Li, T.T.; He, J.J.; Bai, M.J.; Cao, X.Z.; Elsheikha, H.M.; Zhu, X.Q. Toxoplasma gondii tkl1 Deletion Mutant Is a Promising Vaccine against Acute, Chronic, and Congenital Toxoplasmosis in Mice. J. Immunol. 2020, 204, 1562–1570. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.L.; Sun, L.X.; Elsheikha, H.M.; Cao, X.Z.; Nie, L.B.; Li, T.T.; Li, T.S.; Zhu, X.Q.; Wang, J.L. RHΔgra17Δnpt1 Strain of Toxoplasma gondii Elicits Protective Immunity Against Acute, Chronic and Congenital Toxoplasmosis in Mice. Microorganisms 2020, 8, 352. [Google Scholar] [CrossRef] [Green Version]
- Le Roux, D.; Djokic, V.; Morisse, S.; Chauvin, C.; Doré, V.; Lagrée, A.C.; Voisin, D.; Villain, Y.; Grasset-Chevillot, A.; Boursin, F.; et al. Evaluation of immunogenicity and protection of the Mic1-3 knockout Toxoplasma gondii live attenuated strain in the feline host. Vaccine 2020, 38, 1457–1466. [Google Scholar] [CrossRef]
- Ramakrishnan, C.; Maier, S.; Walker, R.A.; Rehrauer, H.; Joekel, D.E.; Winiger, R.R.; Basso, W.U.; Grigg, M.E.; Hehl, A.B.; Deplazes, P.; et al. An experimental genetically attenuated live vaccine to prevent transmission of Toxoplasma gondii by cats. Sci. Rep. 2019, 9, 1474. [Google Scholar] [CrossRef]
- Maeda, Y.; Kinoshita, T. Structural remodeling, trafficking and functions of glycosylphosphatidylinositol-anchored proteins. Prog. Lipid Res. 2011, 50, 411–424. [Google Scholar] [CrossRef]
- Götze, S.; Reinhardt, A.; Geissner, A.; Azzouz, N.; Tsai, Y.-H.; Kurucz, R.; Silva, D.V.; Seeberger, P.H. Investigation of the protective properties of glycosylphosphatidylinositol-based vaccine candidates in a Toxoplasma gondii mouse challenge model. Glycobiology 2015, 25, 984–991. [Google Scholar] [CrossRef] [Green Version]
- Teimouri, A.; Azami, S.J.; Keshavarz, H.; Esmaeili, F.; Alimi, R.; Mavi, S.A.; Shojaee, S. Anti-Toxoplasma activity of various molecular weights and concentrations of chitosan nanoparticles on tachyzoites of RH strain. Int J. Nanomed. 2018, 13, 1341–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myhr, A.I. DNA Vaccines: Regulatory Considerations and Safety Aspects. Curr. Issues Mol. Biol. 2017, 22, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Robinson, H.L.; Pertmer, T.M. DNA vaccines for viral infections: Basic studies and applications. Adv. Virus Res. 2000, 55, 1–74. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Saade, F.; Petrovsky, N. The future of human DNA vaccines. J. Biotechnol. 2012, 162, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Effio, C.L.; Hubbuch, J. Next generation vaccines and vectors: Designing downstream processes for recombinant protein-based virus-like particles. Biotechnol. J. 2015, 10, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Astronomo, R.D.; Burton, D.R. Carbohydrate vaccines: Developing sweet solutions to sticky situations? Nat. Rev. Drug Discov. 2010, 9, 308–324. [Google Scholar] [CrossRef] [Green Version]
- Yeh, M.T.; Bujaki, E.; Dolan, P.T.; Smith, M.; Wahid, R.; Konz, J.; Weiner, A.J.; Bandyopadhyay, A.S.; Van Damme, P.; De Coster, I.; et al. Engineering the Live-Attenuated Polio Vaccine to Prevent Reversion to Virulence. Cell Host Microbe 2020, 27, 736–751. [Google Scholar] [CrossRef]
- Di Cristina, M.; Carruthers, V.B. New and emerging uses of CRISPR/Cas9 to genetically manipulate apicomplexan parasites. Parasitology 2018, 145, 1119–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A. Building a better malaria vaccine. Nature 2019, 575, S51–S54. [Google Scholar] [CrossRef] [Green Version]
- Howe, D.K.; Sibley, L.D. Toxoplasma gondii Comprises Three Clonal Lineages: Correlation of Parasite Genotype with Human Disease. J. Infect. Dis. 1995, 172, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Innes, E.A.; Hamilton, C.; Garcia, J.L.; Chryssafidis, A.; Smith, D. A one health approach to vaccines against Toxoplasma gondii. Food Waterborne Parasitol. 2019, 15, e00053. [Google Scholar] [CrossRef] [PubMed]
- Elbez-Rubinstein, A.; Ajzenberg, D.; Dardé, M.; Cohen, R.; Dumètre, A.; Yera, H.; Gondon, E.; Janaud, J.; Thulliez, P. Congenital Toxoplasmosis and Reinfection during Pregnancy: Case Report, Strain Characterization, Experimental Model of Reinfection, and Review. J. Infect. Dis. 2009, 199, 280–285. [Google Scholar] [CrossRef]


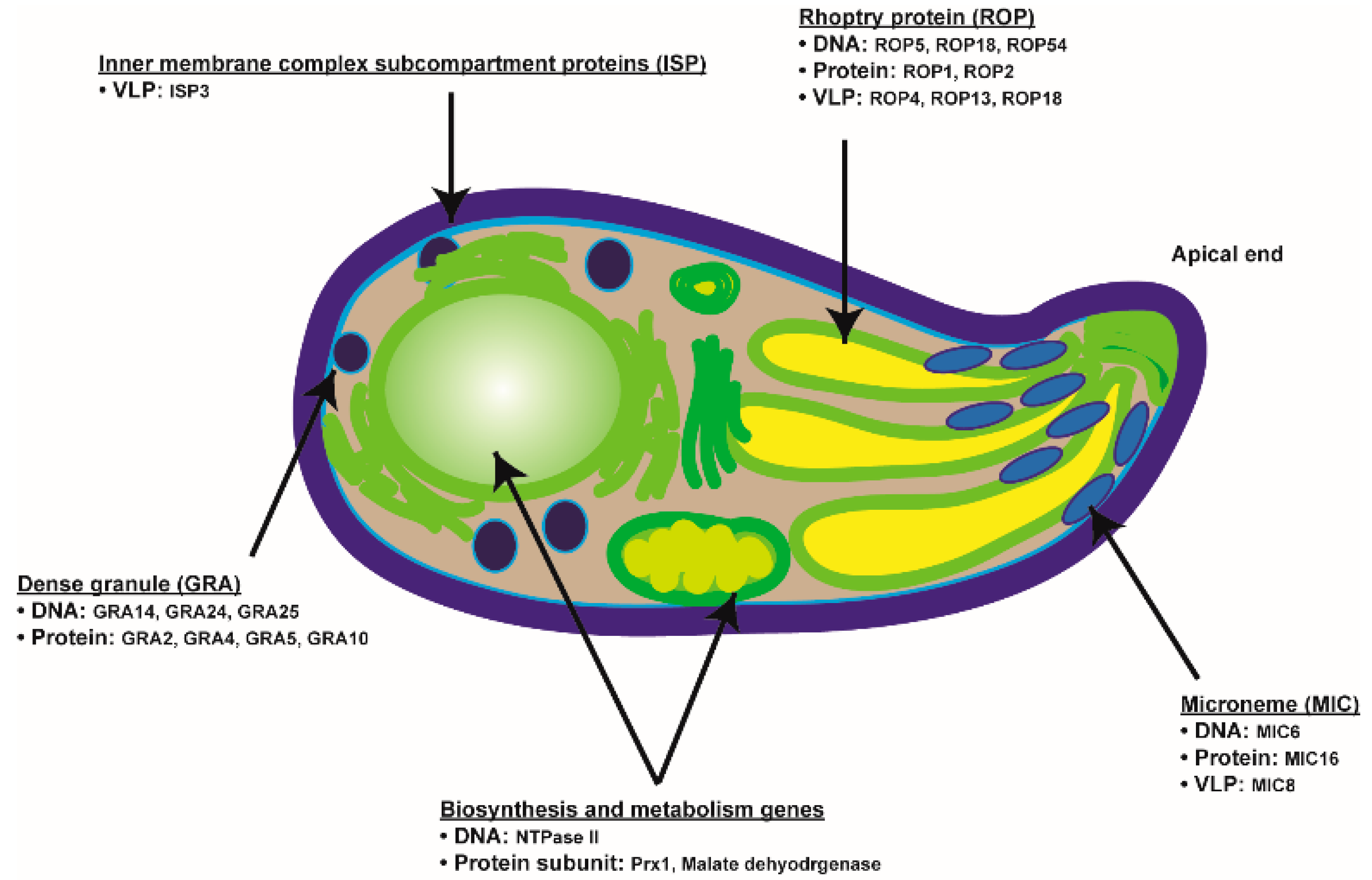{kind=link}
| Vaccine Platform | Animal Model | Antibody Response | Cellular Immune Response | Parasite Burden Reduction | Survival | Reference |
|---|---|---|---|---|---|---|
| DNA vaccine | Mouse | |||||
| ROP54 | + | ++ | + | RH: 15 days | [13] | |
| GRA16 | + | ++ | + | RH: 10 days | [16] | |
| HSP40 | + | + | + | RH: 10 days | [18] | |
| DOC2C | + | ++ | + | RH: >15 days | [22] | |
| GRA14 | ++ | + | ++ | RH: >8 days | [23,24] | |
| ROP5+ROP18 | +++ | ++ | ++ | RH: >28 days | [28] | |
| GRA24+GRA25+MIC6 | ++ | +++ | ++ | RH: >20 days | [32] | |
| NTPase II | + | ++ | + | RH: 18 days, 20% | [36,37] |
| Vaccine Platform | Animal Model | Antibody Response | Cellular Immune Response | Parasite Burden Reduction | Survival | Reference |
|---|---|---|---|---|---|---|
| Protein subunit | ||||||
| Multiantigenic epitopes | ++ | + | + | RH: 16 days | [48] | |
| Multi-epitope | ++ | + | ++ | RH: 21 days | [49] | |
| Prx1 | +++ | + | + | PLK: >60% | [50] | |
| MIF | Mouse | +++ | + | ++ | RH: >30% | [51] |
| Malate dehydrogenase | ++ | ++ | ++ | RH: 50% | [52] | |
| Actin depolF | ++ | + | + | RH: 40% | [53] | |
| CDPK3 | +++ | + | + | RH: 16 days | [54] | |
| Tyrosine hydroxylase | +++ | ++ | + | RH: 18 days; PRU: 50% | [55] | |
| ERK7 | + | ++ | + | RH: 19 days; PRU: 50% | [56] |
| Vaccine Platform | Animal Model | Antibody Response | Cellular Immune Response | Parasite Burden Reduction | Survival | Reference |
|---|---|---|---|---|---|---|
| Virus-like particles | ||||||
| ROP4, ROP13 | ++ | ++ | +++ | ME49: 100% | [82] | |
| IMC, ROP18, MIC8 | Mouse | ++ | + | +++ | ME49: 100% | [85,86] |
| ROP18+MIC8 | ++ | + | ++ | GT1: 17 days | [87] | |
| Multi-antigenic epitope | + | + | + | RH: 20 days | [88] |
| Vaccine Platform | Animal Model | Antibody Response | Cellular Immune Response | Parasite Burden Reduction | Survival | Reference |
|---|---|---|---|---|---|---|
| Live-attenuated | ||||||
| Δα-amylase | ++ | ++ | ++ | RH, ME49, VEG, C7719, | [92] | |
| WH1: >90% | ||||||
| ΔAdsl | Mouse | ++ | +++ | ++ | RH, VEG, ME49: 100% | [93] |
| ΔNpt1 | ++ | +++ | +++ | RH, PYS: 100% | [94] | |
| ΔGra17 | ++ | + | +++ | RH, PRU, PYS, TgC7: 100% | [95] | |
| ΔCdpk2 | ++ | + | +++ | RH, PRU, PYS, TgC7: 100% | [99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, K.-B.; Quan, F.-S. Advances in Toxoplasma gondii Vaccines: Current Strategies and Challenges for Vaccine Development. Vaccines 2021, 9, 413. https://doi.org/10.3390/vaccines9050413
Chu K-B, Quan F-S. Advances in Toxoplasma gondii Vaccines: Current Strategies and Challenges for Vaccine Development. Vaccines. 2021; 9(5):413. https://doi.org/10.3390/vaccines9050413
Chicago/Turabian StyleChu, Ki-Back, and Fu-Shi Quan. 2021. "Advances in Toxoplasma gondii Vaccines: Current Strategies and Challenges for Vaccine Development" Vaccines 9, no. 5: 413. https://doi.org/10.3390/vaccines9050413
APA StyleChu, K. -B., & Quan, F. -S. (2021). Advances in Toxoplasma gondii Vaccines: Current Strategies and Challenges for Vaccine Development. Vaccines, 9(5), 413. https://doi.org/10.3390/vaccines9050413