
Comparative Study of Polycaprolactone Electrospun Fibers and Casting Films Enriched with Carbon and Nitrogen Sources and Their Potential Use in Water Bioremediation


 , ,
, ,  , and
, and 
Abstract
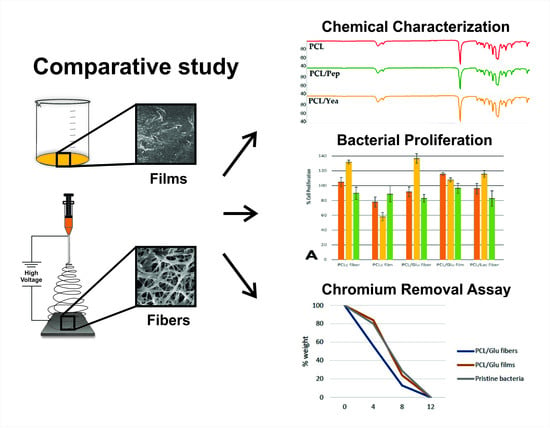
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Polymeric Blends
2.3. Films and Fibers
2.3.1. Electrospinning
2.3.2. Film Production
2.4. Fourier Transform Infrared Spectroscopy (FTIR)
2.5. Differential Scanning Calorimetry (DSC) and Thermogravimetric Analysis (TGA)
2.6. Culture Medium Preparation, Inoculation and Adjustment
2.7. Bacterial Cell Growth Assay
2.8. Biofilm Formation Observation by Scanning Electron Microscopy (SEM)
2.9. Chromium Removal Assay
2.10. Statistical Analysis
3. Results
3.1. Fourier Transform Infrared Spectroscopy (FTIR)
3.2. Differential Scanning Calorimetry (DSC) and Thermogravimetric Analysis (TGA)
3.3. Bacterial Cell Growth Assay
3.4. Biofilm Formation Observation by Scanning Electron Microscopy (SEM)
3.5. Chromium Removal Assay
4. Discussion
4.1. Fourier Transform Infrared Spectroscopy
4.2. Thermogravimetric Assay and Differential Scanning Calorimetry
4.3. Bacterial Cell Growth Assay
4.4. Biofilm Formation Observation by Scanning Electron Microscopy (SEM)
4.5. Chromium Removal Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lee, Y.-S.; Arinzeh, T.L. Electrospun Nanofibrous Materials for Neural Tissue Engineering. Polymers 2011, 3, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Guihard, P.; Danger, Y.; Brounais, B.; David, E.; Brion, R.; Delecrin, J.; Richards, C.D.; Chevalier, S.; Rédini, F.; Heymann, D.; et al. Induction of Osteogenesis in Mesenchymal Stem Cells by Activated Monocytes/Macrophages Depends on Oncostatin M Signaling. Stem Cells 2012, 30, 762–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal, B.B.J.; Wakabayashi, N.; Oyama, K.; Kamiya, H.; Braghirolli, D.I.; Pranke, P. Vascular Tissue Engineering: Polymers and Methodologies for Small Caliber Vascular Grafts. Front. Cardiovasc. Med. 2021, 7, 592361. [Google Scholar] [CrossRef] [PubMed]
- Aviss, K.; Gough, J.; Downes, S. Aligned electrospun polymer fibres for skeletal muscle regeneration. Eur. Cells Mater. 2010, 19, 193–204. [Google Scholar] [CrossRef]
- Abrigo, M.; McArthur, S.L.; Kingshott, P. Electrospun Nanofibers as Dressings for Chronic Wound Care: Advances, Challenges, and Future Prospects. Macromol. Biosci. 2014, 14, 772–792. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Suárez, A.S.; Dastager, S.G.; Bogdanchikova, N.; Grande, D.; Pestryakov, A.; García-Ramos, J.C.; Pérez-González, G.L.; Juárez-Moreno, K.; Toledano-Magaña, Y.; Smolentseva, E.; et al. Electrospun Fibers and Sorbents as a Possible Basis for Effective Composite Wound Dressings. Micromachines 2020, 11, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarreal-Gómez, L.J.; Vera-Graziano, R.; Vega-Rios, M.R.; Pineda-Camacho, J.L.; Mier-Maldonado, P.A.; Almanza-Reyes, H.; Bravo, J.M.C. In Vivo Biocompatibility of Dental Scaffolds for Tissue Regeneration. Adv. Mater. Res. 2014, 976, 191–195. [Google Scholar] [CrossRef]
- Villarreal-Gómez, L.J.; Cornejo-Bravo, J.M.; Vera-Graziano, R.; Grande, D.; Jesús, V.-G.L.; Manuel, C.-B.J.; Ricardo, V.-G.; Daniel, G. Electrospinning as a Powerful Technique for Biomedical Applications: A Critically Selected Survey. J. Biomater. Sci. Polym. Ed. 2016, 27, 157–176. [Google Scholar] [CrossRef]
- Velasco Barraza, R.D.; Álvarez Suarez, A.S.; Villareal-Gómez, L.J.; Paz González, J.A.; Iglesias, A.L.; Vera Graziano, R. Designing a Low-Cost Electrospinning Device for Practical Learning in a Bioengineering Biomaterials Course. Rev. Mex. Ing. Biomed. 2016, 37, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, N.; Kundu, S.C. Electrospinning: A fascinating fiber fabrication technique. Biotechnol. Adv. 2010, 28, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Subrahmanya, T.M.; Bin Arshad, A.; Lin, P.T.; Widakdo, J.; K, M.H.; Austria, H.F.M.; Hu, C.-C.; Lai, J.-Y.; Hung, W.-S. A review of recent progress in polymeric electrospun nanofiber membranes in addressing safe water global issues. RSC Adv. 2021, 11, 9638–9663. [Google Scholar] [CrossRef]
- Massaglia, G.; Sacco, A.; Chiodoni, A.; Pirri, C.F.; Quaglio, M. Living Bacteria Directly Embedded into Electrospun Nanofibers: Design of New Anode for Bio-Electrochemical Systems. Nanomaterials 2021, 11, 3088. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A. Electrospun Nanofibers as Carriers of Microorganisms, Stem Cells, Proteins, and Nucleic Acids in Therapeutic and Other Applications. Front. Bioeng. Biotechnol. 2020, 8, 130. [Google Scholar] [CrossRef]
- Massaglia, G.; Frascella, F.; Chiadò, A.; Sacco, A.; Marasso, S.L.; Cocuzza, M.; Pirri, C.F.; Quaglio, M. Electrospun Nanofibers: From Food to Energy by Engineered Electrodes in Microbial Fuel Cells. Nanomaterials 2020, 10, 523. [Google Scholar] [CrossRef] [Green Version]
- Damasceno, R.; Roggia, I.; Pereira, C.; De Sá, E. Rhizobia survival in seeds coated with polyvinyl alcohol (PVA) electrospun nanofibres. Can. J. Microbiol. 2013, 59, 716–719. [Google Scholar] [CrossRef]
- Gafri, H.F.S.; Zuki, F.M.; Aroua, M.K.; Hashim, N.A. Mechanism of bacterial adhesion on ultrafiltration membrane modified by natural antimicrobial polymers (chitosan) and combination with activated carbon (PAC). Rev. Chem. Eng. 2019, 35, 421–443. [Google Scholar] [CrossRef]
- Velasco-Barraza, R.D.; Vera-Graziano, R.; López-Maldonado, E.A.; Oropeza-Guzmán, M.T.; Dastager, S.G.; Álvarez-Andrade, A.; Iglesias, A.L.; Villarreal-Gómez, L.J. Study of nanofiber scaffolds of PAA, PAA/CS, and PAA/ALG for its potential use in biotechnological applications. Int. J. Polym. Mater. Polym. Biomater. 2017, 67, 800–807. [Google Scholar] [CrossRef]
- Moffa, M.; Pasanisi, D.; Scarpa, E.; Marra, A.R.; Alifano, P.; Pisignano, D. Secondary Metabolite Production from Industrially Relevant Bacteria is Enhanced by Organic Nanofibers. Biotechnol. J. 2017, 12, 1700313. [Google Scholar] [CrossRef]
- Aliah, N.N.; Ansari, M.N.M. Thermal analysis on characterization of polycaprolactone (PCL)-chitosan scaffold for tissue engineering. Int. J. Sci. Res. Eng. Technol. 2017, 6, 2278–2882. [Google Scholar]
- França, D.C.; Bezerra, E.B.; de Morais, D.D.D.S.; Araújo, E.M.; Wellen, R.M.R. Hydrolytic and Thermal Degradation of PCL and PCL/Bentonite Compounds. Mater. Res. 2016, 19, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Kregiel, D. Advances in biofilm control for food and beverage industry using organo-silane technology: A review. Food Control 2014, 40, 32–40. [Google Scholar] [CrossRef]
- Turki, Y.; Mehri, I.; Lajnef, R.; Ben Rejab, A.; Khessairi, A.; Cherif, H.; Ouzari, H.; Hassen, A. Biofilms in bioremediation and wastewater treatment: Characterization of bacterial community structure and diversity during seasons in municipal wastewater treatment process. Environ. Sci. Pollut. Res. 2017, 24, 3519–3530. [Google Scholar] [CrossRef] [PubMed]
- Gamarra, N.N.; Villena, G.K.; Gutiérrez-Correa, M. Cellulase production by Aspergillus niger in biofilm, solid-state, and submerged fermentations. Appl. Microbiol. Biotechnol. 2010, 87, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Verma, N.; Banerjee, B.; Vibha, K.; Haque, S.; Tripathi, C.K.M. Enzymatic degradation of bacterial biofilms using Aspergillus clavatus MTCC 1323. Microbiology 2015, 84, 59–64. [Google Scholar] [CrossRef]
- Zou, M.; Liu, D. A systematic characterization of the distribution, biofilm-forming potential and the resistance of the biofilms to the CIP processes of the bacteria in a milk powder processing factory. Food Res. Int. 2018, 113, 316–326. [Google Scholar] [CrossRef]
- Blana, V.; Georgomanou, A.; Giaouris, E. Assessing biofilm formation by Salmonella enterica serovar Typhimurium on abiotic substrata in the presence of quorum sensing signals produced by Hafnia alvei. Food Control 2017, 80, 83–91. [Google Scholar] [CrossRef]
- Dasgupta, D.; Ghosh, R.; Sengupta, T.K. Biofilm-Mediated Enhanced Crude Oil Degradation by Newly Isolated Pseudomonas Species. ISRN Biotechnol. 2013, 2013, 250749. [Google Scholar] [CrossRef] [Green Version]
- Rossi, E.; Cimdins, A.; Lüthje, P.; Brauner, A.; Sjöling, A.; Landini, P.; Römling, U. “It’s a gut feeling”–Escherichia colibiofilm formation in the gastrointestinal tract environment. Crit. Rev. Microbiol. 2017, 44, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, A.G.; Kaliwal, B.B. Bioremediation of Methomyl by Escherichia coli. In Toxicity and Biodegradation Testing; Bidoia, E., Montagnolli, R., Eds.; Humana Press; Methods in pharmacology and toxicology; New York, NY, USA, 2017; pp. 75–86. [Google Scholar] [CrossRef]
- Khanafer, M.; Al-Awadhi, H.; Radwan, S. Coliform Bacteria for Bioremediation of Waste Hydrocarbons. BioMed Res. Int. 2017, 2017, 1838072. [Google Scholar] [CrossRef] [Green Version]
- Oaikhena, E.E.; Makaije, D.B.; Denwe, S.D.; Namadi, M.M.; Haroun, A.A. Bioremediation Potentials of Heavy Metal Tolerant Bacteria Isolated from Petroleum Refinery Effluent. Am. J. Environ. Prot. 2016, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Abatenh, E.; Gizaw, B.; Tsegaye, Z.; Wassie, M. The Role of Microorganisms in Bioremediation- A Review. Open J. Environ. Biol. 2017, 2, 038–046. [Google Scholar] [CrossRef] [Green Version]
- Rashtchian, M.; Hivechi, A.; Bahrami, S.H.; Milan, P.B.; Simorgh, S. Fabricating alginate/poly (caprolactone) nanofibers with enhanced bio-mechanical properties via cellulose nanocrystal incorporation. Carbohydr. Polym. 2020, 233, 115873. [Google Scholar] [CrossRef] [PubMed]
- Nickless, E.M.; Holroyd, S.E.; Hamilton, G.; Gordon, K.C.; Wargent, J.J. Analytical method development using FTIR-ATR and FT-Raman spectroscopy to assay fructose, sucrose, glucose and dihydroxyacetone, in Leptospermum scoparium nectar. Vib. Spectrosc. 2016, 84, 38–43. [Google Scholar] [CrossRef]
- Wang, P.; Wang, N.; Pang, S.-F.; Zhang, Y.-H. Hygroscopicity of internally mixed particles glycine/NaNO3 studied by FTIR-ATR technique. J. Aerosol Sci. 2018, 116, 25–33. [Google Scholar] [CrossRef]
- Vélez, J.M.B.; Martínez, J.G.; Ospina, J.T.; Agudelo, S.O. Bioremediation potential of Pseudomonas genus isolates from residual water, capable of tolerating lead through mechanisms of exopolysaccharide production and biosorption. Biotechnol. Rep. 2021, 32, e00685. [Google Scholar] [CrossRef] [PubMed]
- Wieringa, P.; Truckenmuller, R.; Micera, S.; van Wezel, R.; Moroni, L. Tandem electrospinning for heterogeneous nanofiber patterns. Biofabrication 2020, 12, 025010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deforet, M.; van Ditmarsch, D.; Xavier, J.B. Cell-Size Homeostasis and the Incremental Rule in a Bacterial Pathogen. Biophys. J. 2015, 109, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Chan-Chan, L.H.; González-García, G.; Vargas-Coronado, R.F.; Cervantes-Uc, J.M.; Hernández-Sánchez, F.; Marcos-Fernandez, A.; Cauich-Rodríguez, J.V. Characterization of model compounds and poly(amide-urea) urethanes based on amino acids by FTIR, NMR and other analytical techniques. Eur. Polym. J. 2017, 92, 27–39. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska, M.; Masek, A. Saccharomyces Cerevisiae Cell Wall Components as Tools for Ochratoxin a Decontamination. Toxins 2015, 7, 1151–1162. [Google Scholar] [CrossRef] [Green Version]
- Shoja, M.; Shameli, K.; Ahmad, M.B.; Zakaria, Z. Preparation and characterization of poly (ε- caprolactone)/TiO2 microcomposites. Dig. J. Nanomater. Biostruct. 2015, 10, 471–477. [Google Scholar]
- Hadj-Hamou, A.S.; Metref, F.; Yahiaoui, F. Thermal stability and decomposition kinetic studies of antimicrobial PCL/nanoclay packaging films. Polym. Bull. 2017, 74, 3833–3853. [Google Scholar] [CrossRef]
- Bezerra, E.B.; França, D.C.; Morais, D.D.D.S.; Rosa, M.D.F.; Morais, J.P.S.; Araújo, E.M.; Wellen, R.M.R. Processing and Properties of PCL/Cotton Linter Compounds. Mater. Res. 2017, 20, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, J.; Li, R.; Lu, A.; Li, Y. Thermal and X-ray Diffraction Analysis of Lactose Polymorph. Procedia Eng. 2015, 102, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.R.; Varma, A.J. Thermal stability of cellulose and their nanoparticles: Effect of incremental increases in carboxyl and aldehyde groups. Carbohydr. Polym. 2014, 114, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Oshima, H.; Kinoshita, M. Effects of sugars on the thermal stability of a protein. J. Chem. Phys. 2013, 138, 245101. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, M.K.; Branton, A.; Trivedi, D.; Nayak, G.; Singh, R.; Jana, S. Physical, Spectroscopic and Thermal Characterization of Biofield Treated Fish Peptone. Eur. J. Biophys. 2015, 3, 51–58. [Google Scholar] [CrossRef]
- Kumar, R.A.; Vizhi, R.E.; Sivakumar, N.; Vijayan, N.; Babu, D.R. Crystal growth, optical and thermal studies of nonlinear optical γ-glycine single crystal grown from lithium nitrate. Int. J. Light Electron. Opt. 2012, 123, 409–413. [Google Scholar] [CrossRef]
- Ercan, I.; Kaygili, O.; Ates, T.; Gunduz, B.; Bulut, N.; Koytepe, S.; Ozcan, I. The effects of urea content on the structural, thermal and morphological properties of MgO nanopowders. Ceram. Int. 2018, 44, 14523–14527. [Google Scholar] [CrossRef]
- Doran, P.M. Advanced power cycles for coal-fired power plants based on calcium looping combustion: A techno-economic feasibility assessment. Appl. Energy 2020, 269, 114954. [Google Scholar] [CrossRef]
- Ebeid, E.-Z.; Zakaria, M. Thermal Analysis from Introductory Fundamentals to Advanced Applications, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 1, Available online: https://www.elsevier.com/books/thermal-analysis/ebeid/978-0-323-90191-8 (accessed on 15 December 2021).
- Persat, A.; Inclan, Y.F.; Engel, J.N.; Stone, H.A.; Gitai, Z. Type IV pili mechanochemically regulate virulence factors in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2015, 112, 7563–7568. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Cedillo, E.; Ortega-Lara, W.; Rocha-Pizaña, M.R.; Gutierrez-Uribe, J.A.; Elías-Zúñiga, A.; Rodríguez, C.A. Electrospun Polycaprolactone Fibrous Membranes Containing Ag, TiO2 and Na2Ti6O13 Particles for Potential Use in Bone Regeneration. Membranes 2019, 9, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruckh, T.T.; Oldinski, R.A.; Carroll, D.A.; Mikhova, K.; Bryers, J.D.; Popat, K.C. Antimicrobial effects of nanofiber poly (caprolactone) tissue scaffolds releasing rifampicin. J. Mater. Sci. Mater. Med. 2012, 23, 1411–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeli-Sardou, M.; Torkzadeh-Mahani, M.; Yaghoobi, M.M.; Dodel, M. Antibacterial and anti-biofilm investigation of electrospun PCL/gelatin/Lawsone nano fiber scaffolds against biofilm producing bacteria. Biomacromol. J. 2018, 4, 46–57. Available online: http://www.bmmj.org/article_34368.html (accessed on 10 December 2021).
- Pompa-Monroy, D.A.; Figueroa-Marchant, P.G.; Dastager, S.G.; Thorat, M.N.; Iglesias, A.L.; Miranda-Soto, V.; Pérez-González, G.L.; Villarreal-Gómez, L.J. Bacterial Biofilm Formation Using PCL/Curcumin Electrospun Fibers and Its Potential Use for Biotechnological Applications. Materials 2020, 13, 5556. [Google Scholar] [CrossRef]
- Priyadarsini, K.I. The Chemistry of Curcumin: From Extraction to Therapeutic Agent. Molecules 2014, 19, 20091–20112. [Google Scholar] [CrossRef] [Green Version]
- Himmler, M.; Garreis, F.; Paulsen, F.; Schubert, D.W.; Fuchsluger, T.A. Optimization of polycaprolactone-based nanofiber matrices for the cultivation of corneal endothelial cells. Sci. Rep. 2021, 11, 18858. [Google Scholar] [CrossRef]
- Zheng, H.; Ho, P.-Y.; Jiang, M.; Tang, B.; Liu, W.; Li, D.; Yu, X.; Kleckner, N.E.; Amir, A.; Liu, C. Interrogating the Escherichia coli cell cycle by cell dimension perturbations. Proc. Natl. Acad. Sci. USA 2016, 113, 15000–15005. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Zhang, Z.; Shi, H. Cell Size Is Coordinated with Cell Cycle by Regulating Initiator Protein DnaA in E. coli. Biophys J. 2020, 119, 2537–2557. [Google Scholar] [CrossRef]
- Monteiro, J.M.; Fernandes, P.B.; Vaz, F.; Pereira, A.R.; Tavares, A.C.; Ferreira, M.T.; Pereira, P.M.; Veiga, H.; Kuru, E.; VanNieuwenhze, M.S.; et al. Cell shape dynamics during the Staphylococcal cell cycle. Nat. Commun. 2015, 6, 8055. [Google Scholar] [CrossRef]
- Rnjak-Kovacina, J.; Weiss, A.S. Increasing the Pore Size of Electrospun Scaffolds. Tissue Eng. Part B Rev. 2011, 17, 365–372. [Google Scholar] [CrossRef]
- Yang, S.-Y.; Asmatulu, R.; Ceylan, M. Effects of gentamicin-loaded PCL nanofibers on growth of Gram-positive and Gram-negative bacteria. Int. J. Appl. Microbiol. Biotechnol. Res. 2017, 5, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Gautam, S.; Chou, C.-F.; Dinda, A.K.; Potdar, P.D.; Mishra, N.C. Fabrication and characterization of PCL/gelatin/chitosan ternary nanofibrous composite scaffold for tissue engineering applications. J. Mater. Sci. 2013, 49, 1076–1089. [Google Scholar] [CrossRef]
- Chu, E.K.; Kilic, O.; Cho, H.; Groisman, A.; Levchenko, A. Self-induced mechanical stress can trigger biofilm formation in uropathogenic Escherichia coli. Nat. Commun. 2018, 9, 4087. [Google Scholar] [CrossRef] [PubMed]
- Oura, H.; Tashiro, Y.; Toyofuku, M.; Ueda, K.; Kiyokawa, T.; Ito, S.; Takahashi, Y.; Lee, S.; Nojiri, H.; Nakajima-Kambe, T.; et al. Inhibition of Pseudomonas aeruginosa Swarming Motility by 1-Naphthol and Other Bicyclic Compounds Bearing Hydroxyl Groups. Appl. Environ. Microbiol. 2015, 81, 2808–2818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsogiannis, K.A.G.; Vladisavljević, G.T.; Georgiadou, S. Porous electrospun polycaprolactone (PCL) fibres by phase separation. Eur. Polym. J. 2015, 69, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Walker, G.; Price, S.; Yang, X.; Li, J.; Bunt, C. Electrospun Membranes as a Porous Barrier for Molecular Transport: Membrane Characterization and Release Assessment. Pharmaceutics 2021, 13, 916. [Google Scholar] [CrossRef]
- Fernandes, G.; Bastos, M.C.; De Vargas, J.P.R.; Le Guet, T.; Clasen, B.; Dos Santos, D.R. The use of epilithic biofilms as bioaccumulators of pesticides and pharmaceuticals in aquatic environments. Ecotoxicology 2020, 29, 1293–1305. [Google Scholar] [CrossRef]
- Berlanga, M.; Guerrero, R. Living together in biofilms: The microbial cell factory and its biotechnological implications. Microb. Cell Factories 2016, 15, 165. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, S.; Kaya, T.; Aslim, B.; Tan, S. Removal and reduction of chromium by Pseudomonas sp. and their correlation to rhamnolipid production. J. Hazard. Mater. 2012, 231–232, 64–69. [Google Scholar] [CrossRef]
- Arisah, F.M.; Amir, A.F.; Ramli, N.; Ariffin, H.; Maeda, T.; Hassan, M.A.; Yusoff, M.Z.M. Bacterial Resistance against Heavy Metals in Pseudomonas aeruginosa RW9 Involving Hexavalent Chromium Removal. Sustainability 2021, 13, 9797. [Google Scholar] [CrossRef]
- Dogan, N.M.; Kantar, C.; Gulcan, S.; Dodge, C.J.; Yilmaz, B.C.; Mazmanci, M.A. Chromium (VI) Bioremoval by Pseudomonas Bacteria: Role of Microbial Exudates for Natural Attenuation and Biotreatment of Cr(VI) Contamination. Environ. Sci. Technol. 2011, 45, 2278–2285. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Maiti, A. Study of simultaneous bioremediation of mixed reactive dyes and Cr(VI) containing wastewater through designed experiments. Environ. Monit. Assess. 2019, 191, 766. [Google Scholar] [CrossRef] [PubMed]
- Orandi, S.; Lewis, D.M.; Moheimani, N.R. Biofilm establishment and heavy metal removal capacity of an indigenous mining algal-microbial consortium in a photo-rotating biological contactor. J. Ind. Microbiol. Biotechnol. 2012, 39, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.-H.; Hong, J.-P.; Zhang, Q.; Jin, D.-S.; Gao, C.-H. Contrast in soil microbial metabolic functional diversity to fertilization and crop rotation under rhizosphere and non-rhizosphere in the coal gangue landfill reclamation area of Loess Hills. PLoS ONE 2020, 15, e0229341. [Google Scholar] [CrossRef]
- Chang, J.-S.; Yoon, I.-H.; Lee, J.-H.; Kim, K.-R.; An, J.; Kim, K.-W. Arsenic detoxification potential of aox genes in arsenite-oxidizing bacteria isolated from natural and constructed wetlands in the Republic of Korea. Environ. Geochem. Health 2010, 32, 95–105. [Google Scholar] [CrossRef]
- Tariq, M.; Waseem, M.; Rasool, M.H.; Zahoor, M.A.; Hussain, I. Isolation and molecular characterization of the indigenous Staphylococcus aureus strain K1 with the ability to reduce hexavalent chromium for its application in bioremediation of metal-contaminated sites. PeerJ 2019, 7, e7726. [Google Scholar] [CrossRef] [Green Version]
- Noureddine, N.; Benhammadi, S.; Kara, F.; Aguedal, H.; Iddou, A.; Juhna, T. Purification of Contaminated Water with Chromium (VI) Using Pseudomonas aeruginosa. Key Eng. Mater. 2016, 721, 143–148. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Label | Cs a Solutions | Sample Label | Ns b Solutions |
|---|---|---|---|
| PCL | PCL (Control) | PCL | PCL (Control) |
| PCL/Glu | PCL and Glucose | PCL/Pep | PCL and Peptone |
| PCL/Lac | PCL and Lactose | PCL/Gly | PCL and Glycine |
| PCL/Fru | PCL and Fructose | PCL/Yea | PCL and Yeast Extract |
| PCL/Gal | PCL and Galactose | PCL/Ure | PCL and Urea |
| Sample | Onset Degradation Temperature (Ti) 5% | Critical Degradation Temperature 50% | Final Degradation Temperature 100% | Sample | Onset Degradation Temperature (Ti) 5% | Critical Degradation Temperature 50% | Final Degradation Temperature 100% |
|---|---|---|---|---|---|---|---|
| PCL | 336.23 °C | 388.39 °C | 412.96 °C | PCL/Pep | 342.86 °C | 386.89 °C | 570.56 °C |
| PCL/Glu | 347.51 °C | 393.64 °C | 418.74 °C | PCL/Gly | 333.24 °C | 388.38 °C | 417.52 °C |
| PCL/Fru | 341.56 °C | 392.50 °C | 416.46 °C | PCL/Yea | 305.02 °C | 384.62 °C | 427.38 °C |
| PCL/Lac | 349.87 °C | 392.48 °C | 417.08 °C | PCL/Ure | 273.16 °C | 389.51 °C | 548.68 °C |
| PCL/Gal | 346.33 °C | 392.35 °C | 416.20 °C |
| Sample | Melting Temperature | Decomposition Temperature | Sample | Melting Temperature | Decomposition Temperature |
|---|---|---|---|---|---|
| PCL | 63.55 °C | 397.88 °C | PCL/Pep | 62.92 °C | 395.64 °C |
| PCL/Glu | 62.44 °C | 398.79 °C | PCL/Gly | 61.02 °C | 396.62 °C |
| PCL/Fru | 61.99 °C | 398.28 °C | PCL/Yea | 61.54 °C | 393.37 °C |
| PCL/Lac | 61.61 °C | 397.28 °C | PCL/Ure | 60.73 °C | 395.54 °C |
| PCL/Gal | 61.97 °C | 398.32 °C |
| Sample | Average ± SD (nm) | Sample | Average ± SD (nm) |
|---|---|---|---|
| PCL | 581 ± 129 | PCL/Pep | 480 ± 146 |
| PCL/Glu | 313 ± 089 | PCL/Gly | 473 ± 124 |
| PCL/Fru | 558 ± 162 | PCL/Yea | 766 ± 290 |
| PCL/Lac | 301 ± 072 | PCL/Ure | 332 ± 083 |
| PCL/Gal | 408 ± 143 |
| Organism | Time of Incubation (h) | Removed Chromium (%) | Reference |
|---|---|---|---|
| P. aeruginosa | 48 | 24 | [72] |
| P. aeruginosa 99 | 48 | 45 | [72] |
| P. stutzeri T3 | 48 | 33 | [72] |
| P. aeruginosa RW 9 | 8 | 82 | [73] |
| P. aeruginosa P16 | 45 | 37 | [74] |
| P. aeruginosa 4442 | 8 | 22 | [75] |
| P. aeruginosa | 8 | 71 | Our study |
| PCL/Glu fibers/P. aeruginosa | 8 | 87 | Our study |
| PCL/Glu films/P. aeruginosa | 8 | 76 | Our study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pompa-Monroy, D.A.; Iglesias, A.L.; Dastager, S.G.; Thorat, M.N.; Olivas-Sarabia, A.; Valdez-Castro, R.; Hurtado-Ayala, L.A.; Cornejo-Bravo, J.M.; Pérez-González, G.L.; Villarreal-Gómez, L.J. Comparative Study of Polycaprolactone Electrospun Fibers and Casting Films Enriched with Carbon and Nitrogen Sources and Their Potential Use in Water Bioremediation. Membranes 2022, 12, 327. https://doi.org/10.3390/membranes12030327
Pompa-Monroy DA, Iglesias AL, Dastager SG, Thorat MN, Olivas-Sarabia A, Valdez-Castro R, Hurtado-Ayala LA, Cornejo-Bravo JM, Pérez-González GL, Villarreal-Gómez LJ. Comparative Study of Polycaprolactone Electrospun Fibers and Casting Films Enriched with Carbon and Nitrogen Sources and Their Potential Use in Water Bioremediation. Membranes. 2022; 12(3):327. https://doi.org/10.3390/membranes12030327
Chicago/Turabian StylePompa-Monroy, Daniella Alejandra, Ana Leticia Iglesias, Syed Gulam Dastager, Meghana Namdeo Thorat, Amelia Olivas-Sarabia, Ricardo Valdez-Castro, Lilia Angélica Hurtado-Ayala, José Manuel Cornejo-Bravo, Graciela Lizeth Pérez-González, and Luis Jesús Villarreal-Gómez. 2022. "Comparative Study of Polycaprolactone Electrospun Fibers and Casting Films Enriched with Carbon and Nitrogen Sources and Their Potential Use in Water Bioremediation" Membranes 12, no. 3: 327. https://doi.org/10.3390/membranes12030327
APA StylePompa-Monroy, D. A., Iglesias, A. L., Dastager, S. G., Thorat, M. N., Olivas-Sarabia, A., Valdez-Castro, R., Hurtado-Ayala, L. A., Cornejo-Bravo, J. M., Pérez-González, G. L., & Villarreal-Gómez, L. J. (2022). Comparative Study of Polycaprolactone Electrospun Fibers and Casting Films Enriched with Carbon and Nitrogen Sources and Their Potential Use in Water Bioremediation. Membranes, 12(3), 327. https://doi.org/10.3390/membranes12030327