Possibly Invasive New Bioenergy Crop Silphium perfoliatum: Growth and Reproduction Are Promoted in Moist Soil



Abstract
:1. Introduction
2. Materials and Methods
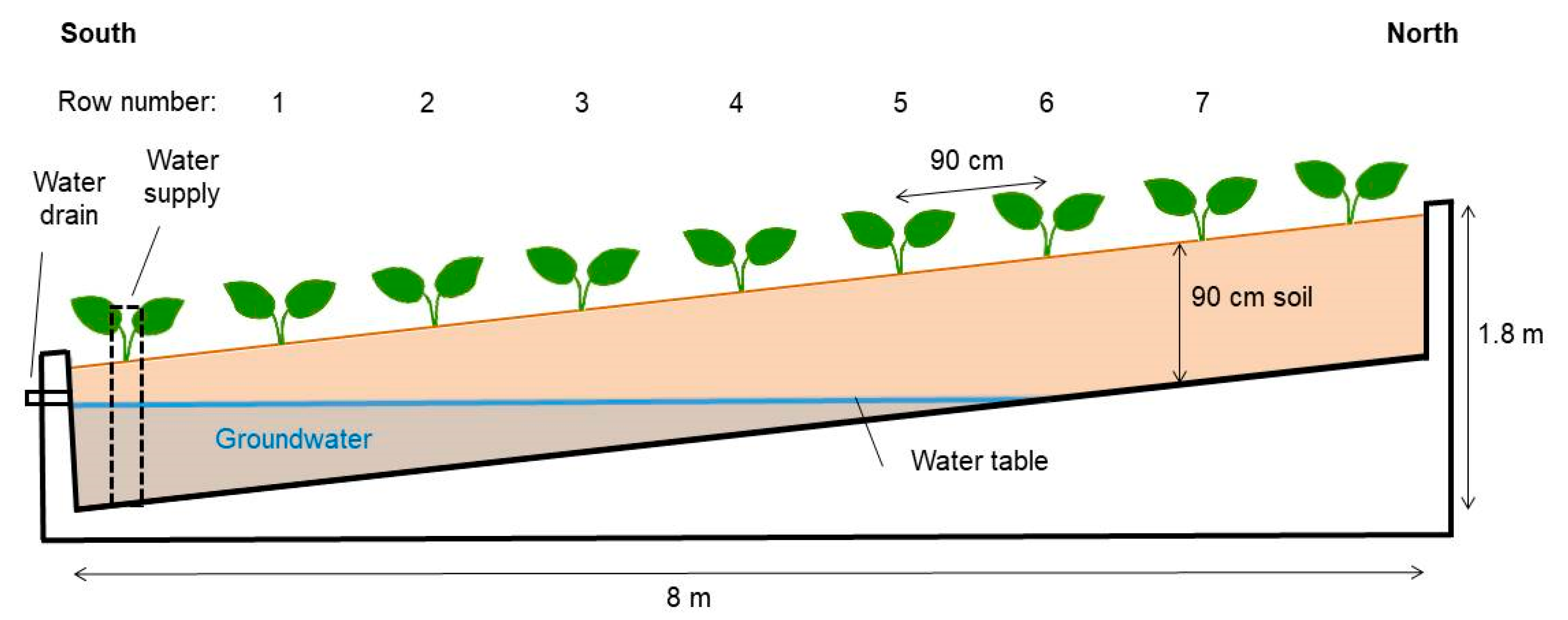
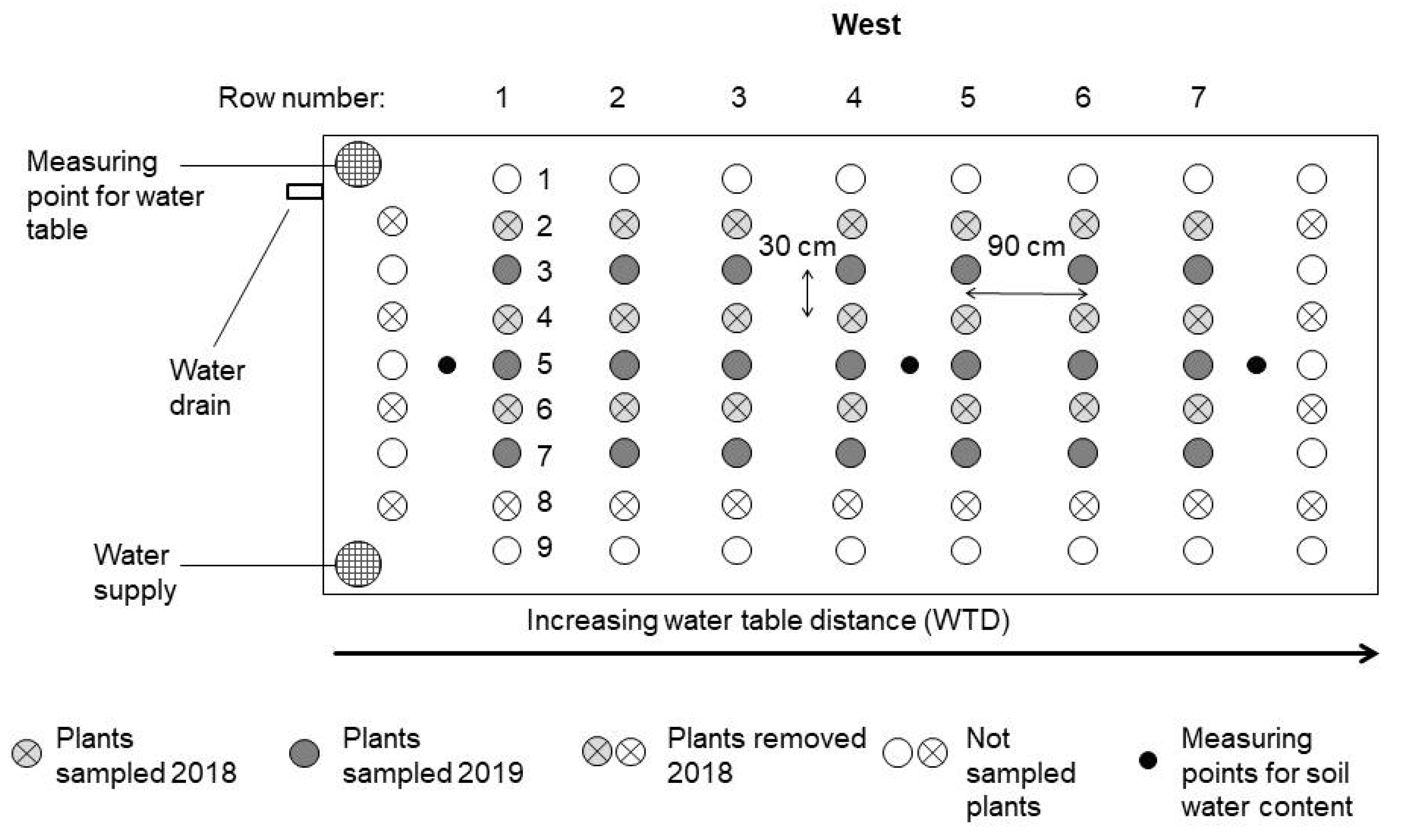
2.1. Experimental Setup
2.2. Data Collection
2.3. Statistics
3. Results
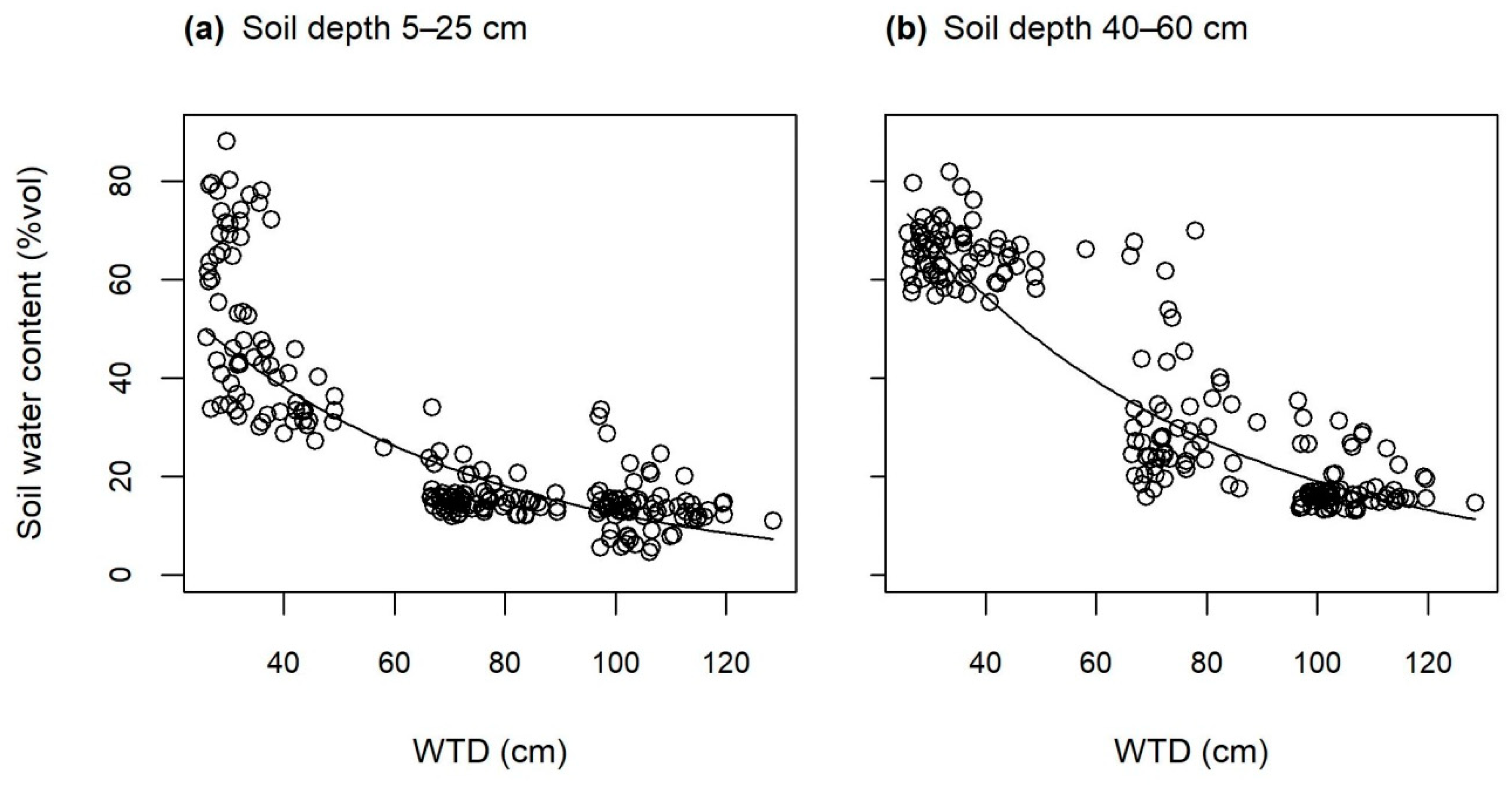
3.1. Soil Water Conditions
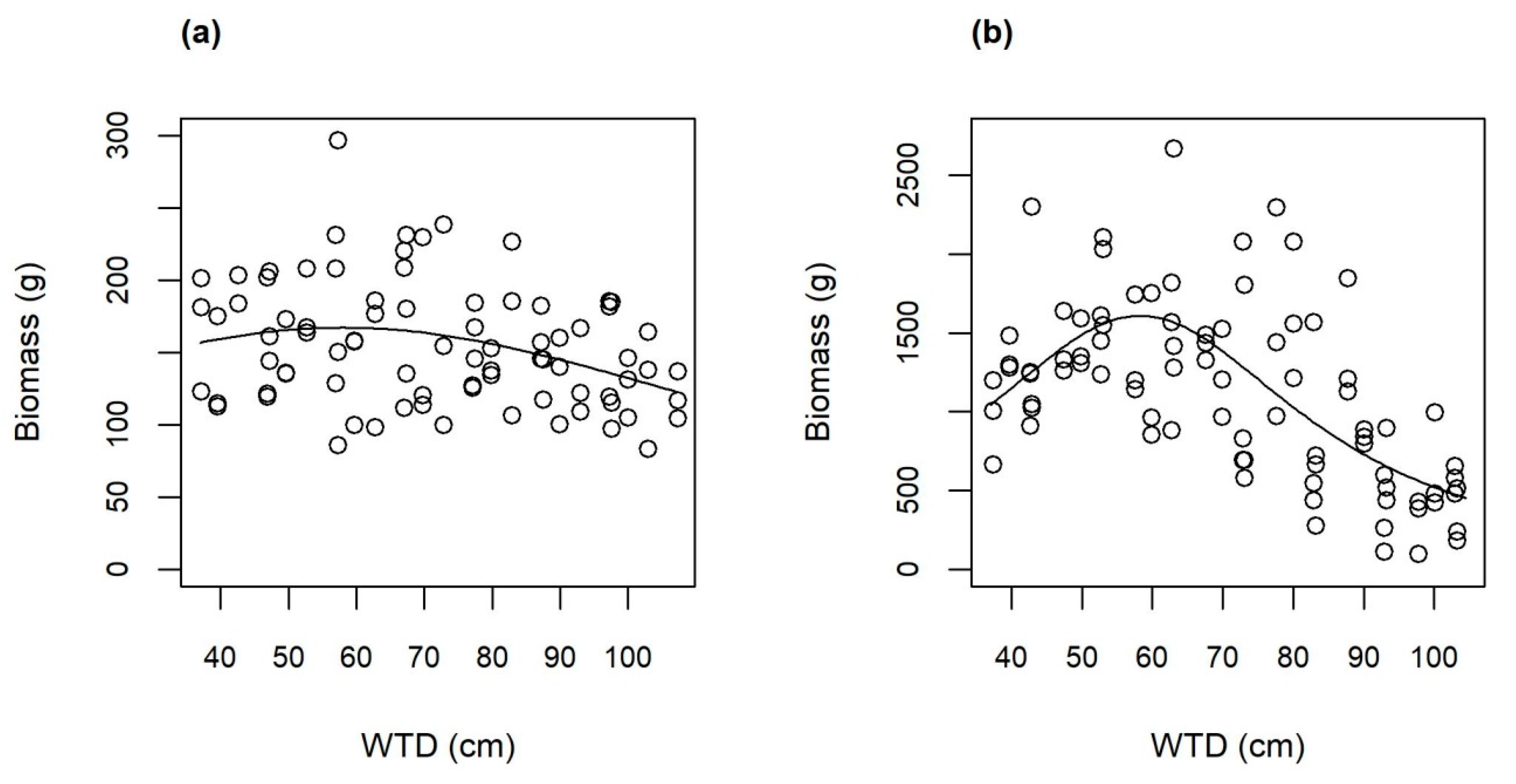
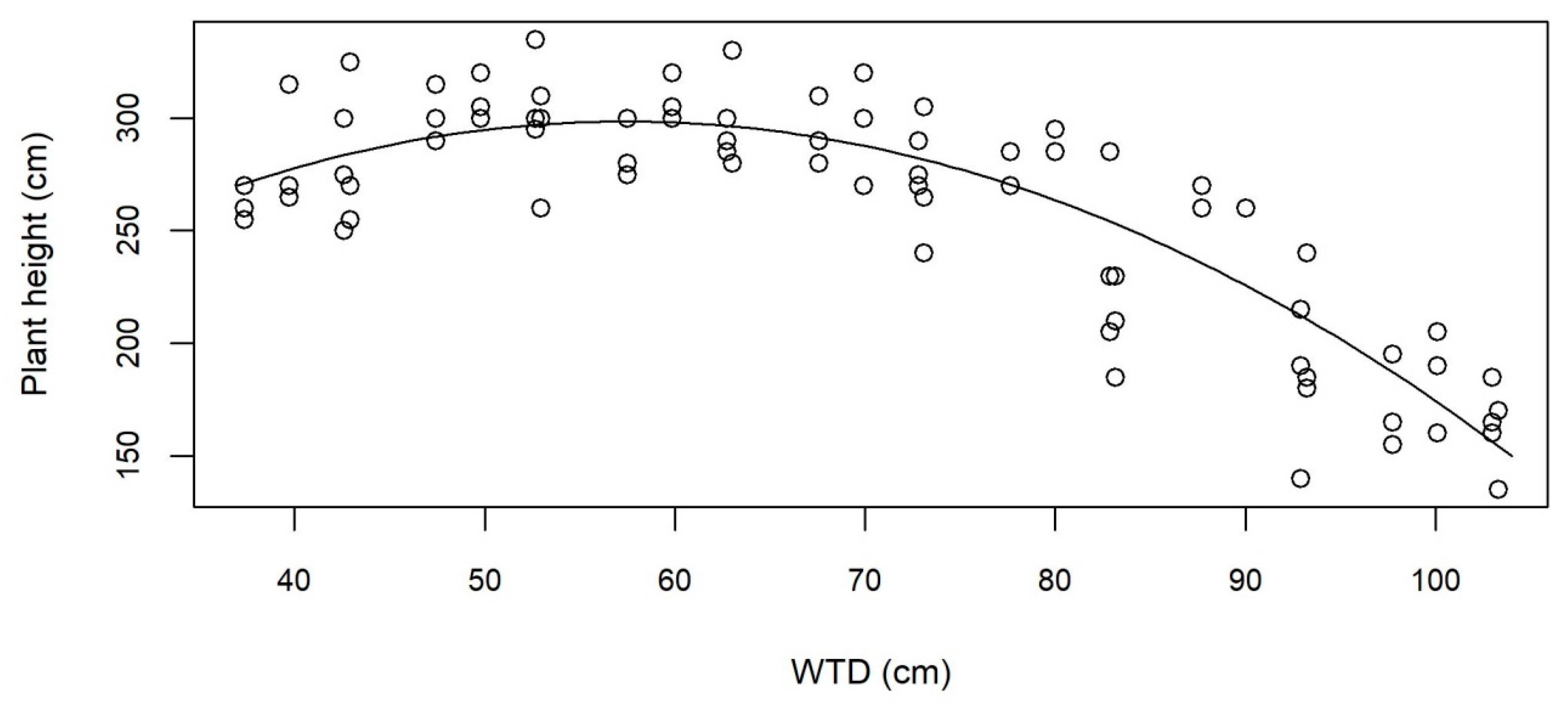
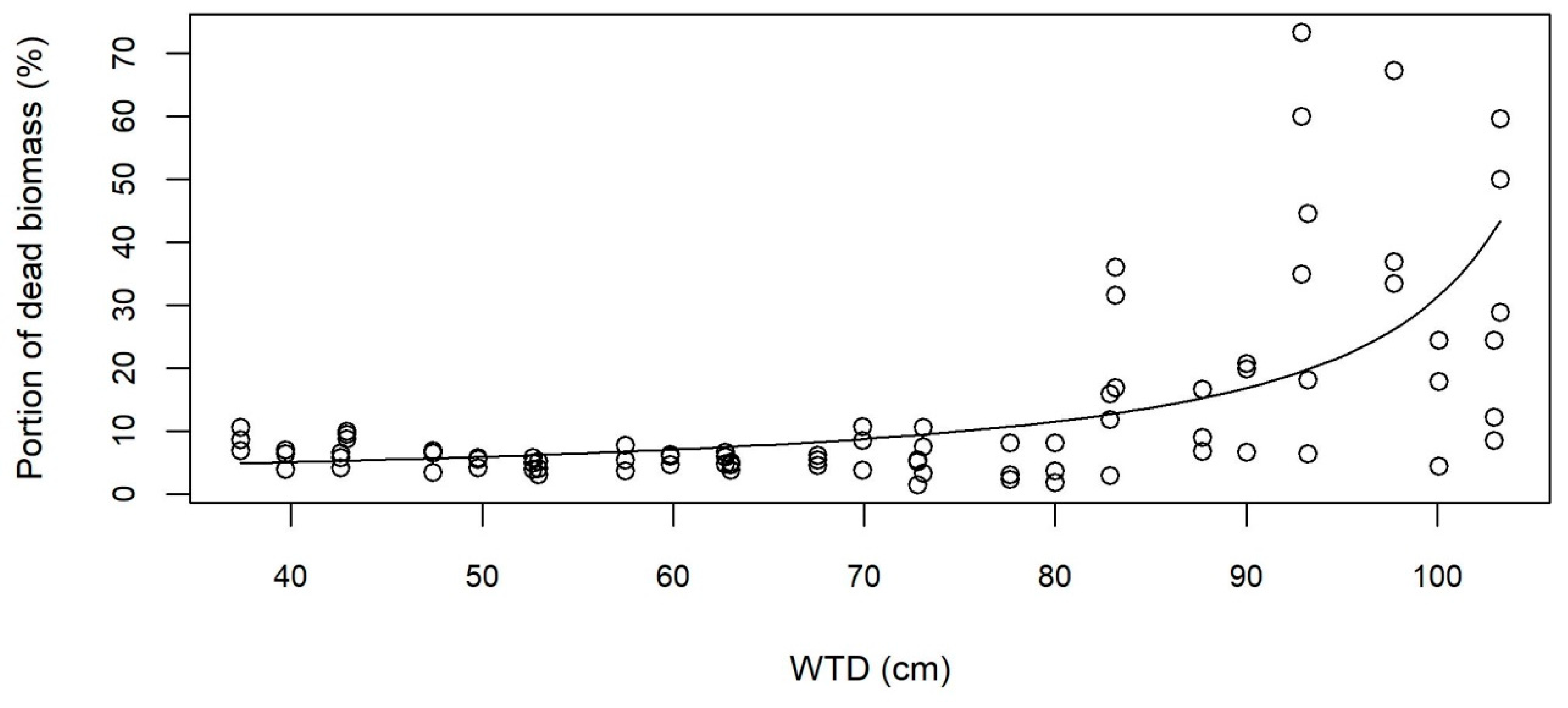
3.2. Growth and Aboveground Biomass
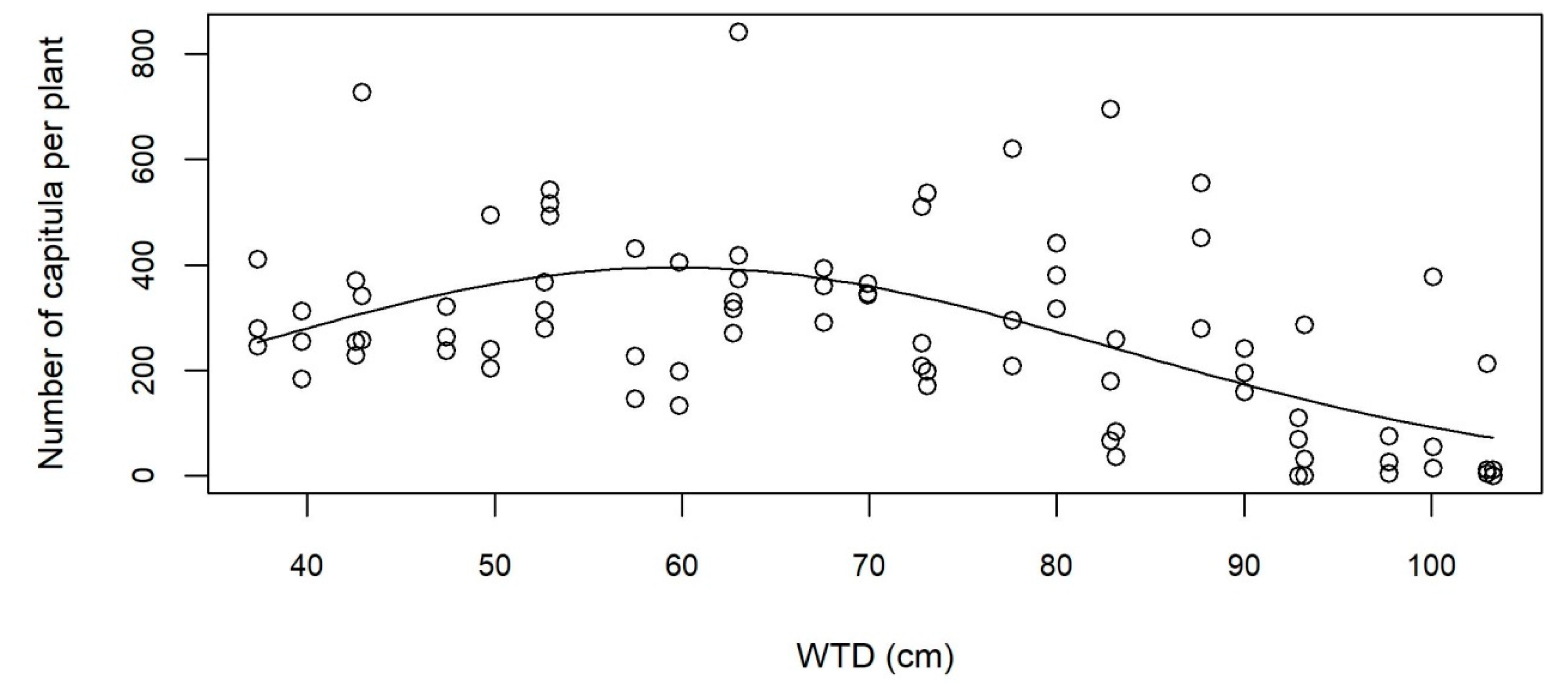
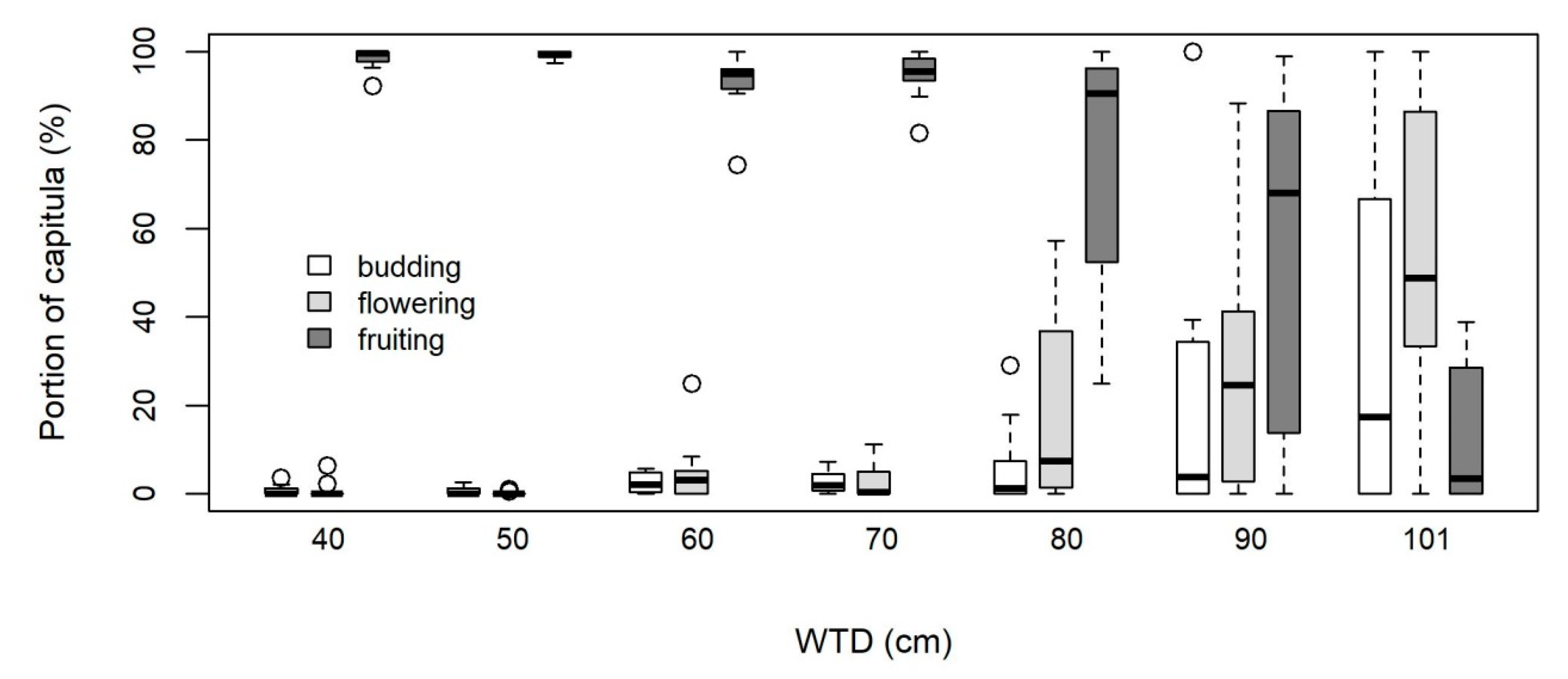
3.3. Reproductive Potential
4. Discussion
4.1. Highest Yield on Moist Soil
4.2. High Reproduction and Rapid Development on Moist Soil
4.3. Consequences and Recommendations for Nature Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IRENA. Renewable Energy Statistics 2019; The International Renewable Energy Agency: Abu Dhabi, UAE, 2019; ISBN 978-92-9260-137-9. [Google Scholar]
- Frölich, W.; Brodmann, R.; Metzler, T. The cup plant (Silphium perfoliatum)—A story of success from agricultural practice. J. Cultiv. Plants 2016, 68, 351–355. [Google Scholar] [CrossRef]
- Gansberger, M.; Montgomery, L.F.R.; Liebhard, P. Botanical characteristics, crop management and potential of Silphium perfoliatum L. as a renewable resource for biogas production: A review. Ind. Crops Prod. 2015, 63, 362–372. [Google Scholar] [CrossRef]
- Emmerling, C. Soil quality through the cultivation of perennial bioenergy crops by example of Silphium perfoliatum—An innovative agro-ecosystem in future. J. Cultiv. Plants 2016, 68, 399–406. [Google Scholar] [CrossRef]
- Ruf, T.; Audu, V.; Holzhauser, K.; Emmerling, C. Bioenergy from Periodically Waterlogged Cropland in Europe: A First Assessment of the Potential of Five Perennial Energy Crops to Provide Biomass and Their Interactions with Soil. Agronomy 2019, 9, 374. [Google Scholar] [CrossRef] [Green Version]
- Stanford, G. Silphium perfoliatum (cup-plant) as a new forage. In Proceedings of the Twelfth North American Prairie Conference, Cedar Falls, IA, USA, 5–9 August 1990; pp. 33–37. [Google Scholar]
- Südwestrundfunk. Anbaufläche von Silphie ist größer geworden. SWR Aktuell. 5 September 2019. Available online: https://www.swr.de/swraktuell/baden-wuerttemberg/friedrichshafen/groessere-anbauflaeche-silphie-100.html (accessed on 15 May 2020).
- Hartmann, A.; Lunenberg, T. Yield potential of cup plant under Bavarian cultivation conditions. J. Cultiv. Plants 2016, 68, 385–388. [Google Scholar] [CrossRef]
- Burmeister, J.; Walter, R. Studies on the ecological effect of Silphium perfoliatum in Bavaria. J. Cultiv. Plants 2016, 68, 407–411. [Google Scholar] [CrossRef]
- Dauber, J.; Müller, A.L.; Schittenhelm, S.; Schoo, B.; Schorpp, Q.; Schrader, S.; Schroetter, S. Schlussbericht zum Vorhaben: Agrarökologische Bewertung der Durchwachsenen Silphie (Silphium perfoliatum L.) als eine Biomassepflanze der Zukunft. 2016. Available online: https://literatur.thuenen.de/digbib_extern/dn056633.pdf (accessed on 13 March 2019).
- Mueller, A.L.; Dauber, J. Hoverflies (Diptera: Syrphidae) benefit from a cultivation of the bioenergy crop Silphium perfoliatum L. (Asteraceae) depending on larval feeding type, landscape composition and crop management. Agric. For. Entomol. 2016, 18, 419–431. [Google Scholar] [CrossRef]
- Ende, L.M.; Lauerer, M. Spontaneous occurrences of the cup plant in the Bayreuth region: Does this new bioenergy crop have invasive potential? Nat. Landsch. 2020, 95, 310–315. [Google Scholar] [CrossRef]
- Roskov, Y.; Ower, G.; Orrell, T.; Nicolson, D.; Bailly, N.; Kirk, P.M.; Bourgoin, T.; DeWalt, R.E.; Decock, W.; van Nieukerken, E.; et al. Species 2000 & ITIS Catalogue of Life, 2019 Annual Checklist. Available online: http://www.catalogueoflife.org/annual-checklist/2019/details/species/id/d1c5f933225a3fa3a7974259458811ac (accessed on 7 February 2020).
- Brennenstuhl, G. Beobachtungen zur Einbürgerung von Gartenflüchtlingen im Raum Salzwedel (Altmark). Mitt. Florist. Kart. Sachs. Anhalt. 2010, 15, 121–134. [Google Scholar]
- Wölfel, U. Zur Flora von Bitterfeld und Umgebung (11. Beitrag). Mitt. Florist. Kart. Sachs. Anhalt 2013, 18, 47–53. [Google Scholar]
- Verbreitung der Farn- und Blütenpflanzen in Deutschland; aggregiert im Raster der Topographischen Karte 1:25000—Silphium perfoliatum. Available online: http://www.floraweb.de/webkarten/karte.html?taxnr=5635 (accessed on 12 March 2019).
- Matthews, J.; Beringen, R.; Huijbregts, M.A.J.; van der Mheen, H.J.; Odé, B.; Trindade, L.; van Valkenburg, J.L.C.H.; van der Velde, G.; Leuven, R.S.E.W. Horizon Scanning and Environmental Risk Analyses of Non-Native Biomass Crops in the Netherlands; Department of Environmental Science, Institute for Water and Wetland Research, Radboud University Nijmegen: Nijmegen, The Netherlands, 2015. [Google Scholar]
- Vinogradova, Y.K.; Mayorov, S.R.; Bochkin, V.D. Changes in the spontaneous flora of the Main Botanic Garden, Moscow, over 65 years. Skvortsovia 2015, 2, 45–95. [Google Scholar]
- Bufe, C.; Korevaar, H. Evaluation of Additional Crops for Dutch List of Ecological Focus Area; Wageningen Research Foundation (WR) business unit Agrosystems Research: Wageningen, The Netherlands, 2018. [Google Scholar] [CrossRef] [Green Version]
- Schoo, B.; Wittich, K.P.; Böttcher, U.; Kage, H.; Schittenhelm, S. Drought Tolerance and Water-Use Efficiency of Biogas Crops: A Comparison of Cup Plant, Maize and Lucerne-Grass. J. Agron. Crop Sci. 2017, 203, 117–130. [Google Scholar] [CrossRef]
- Ellenberg, H. Physiologisches und ökologisches Verhalten derselben Pflanzenarten. Ber. Deutsch. Bot. Ges. 1952, 65, 350–361. [Google Scholar]
- Ellenberg, H. Über einige Fortschritte der kausalen Vegetationskunde. Plant Ecol. 1954, 5, 199–211. [Google Scholar] [CrossRef]
- Hector, A.; von Felten, S.; Hautier, Y.; Weilenmann, M.; Bruelheide, H. Effects of dominance and diversity on productivity along Ellenberg’s experimental water table gradients. PLoS ONE 2012, 7, e43358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H. The Split-Apply-Combine Strategy for Data Analysis. J. Stat. Softw. 2011, 40, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Holzheu, S.; Archner, O. BayEOS: BayEOS Server Access; R package version 1.4.6.: Bayreuth, Germany, 2017. [Google Scholar]
- Giraudoux, P. Pgirmess: Spatial Analyses and Data Mining for Field Ecologists; R Package version 1.6.9.: Besançon, France, 2018. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Soft. 2015, 67. [Google Scholar] [CrossRef]
- Blüthner, W.-D.; Krähmer, A.; Hänsch, K.-T. Breeding progress in cup plant-first steps. J. Cultiv. Plants 2016, 68, 392–398. [Google Scholar] [CrossRef]
- Wrobel, M.; Fraczek, J.; Francik, S.; Slipek, Z.; Krzysztof, M. Influence of degree of fragmentation on chosen quality parameters of briquette made from biomass of cup plant Silphium perfoliatum L. Eng. Rural Dev. 2013, 12, 653–657. [Google Scholar]
- Schittenhelm, S.; Schoo, B.; Schroetter, S. Yield physiology of biogas crops: Comparison of cup plant, maize, and lucerne-grass. J. Cultiv. Plants 2016, 68, 378–384. [Google Scholar] [CrossRef]
- Figas, A.; Siwik-Ziomek, A.; Rolbiecki, R. Effect of irrigation on some growth parameters of cup plant and dehydrogenase activity in soil. Ann. Warsaw Univ. Life Sci. SGGW Land Reclam. 2015, 47, 279–288. [Google Scholar] [CrossRef]
- Boe, A.; Albrecht, K.A.; Johnson, P.J.; Wu, J. Biomass Production of Cup Plant (Silphium perfoliatum L.) in Response to Variation in Plant Population Density in the North Central USA. Am. J. Plant Sci. 2019, 10, 904–910. [Google Scholar] [CrossRef] [Green Version]
- Bury, M.; Możdżer, E.; Kitczak, T.; Siwek, H.; Włodarczyk, M. Yields, Caloric Value and Chemical Properties of Cup Plant Silphium perfoliatum L. Biomass, Depending on the Method of Establishing the Plantation. Agronomy 2020, 10, 851. [Google Scholar] [CrossRef]
- Mueller, A.L.; Berger, C.A.; Schittenhelm, S.; Stever-Schoo, B.; Dauber, J. Water availability affects nectar sugar production and insect visitation of the cup plant Silphium perfoliatum L. (Asteraceae). J. Agron. Crop Sci. 2020. [Google Scholar] [CrossRef] [Green Version]
- Rothmaler—Exkursionsflora von Deutschland. Gefäßpflanzen: Grundband, 21st ed.; Jäger, E.J. (Ed.) Springer Spektrum: Berlin, Germany, 2017; ISBN 978-3-662-49707-4. [Google Scholar]
- Schmeil—Fitschen: Die Flora Deutschlands und Angrenzender Länder, 96th ed.; Parolly, G.; Rohwer, J.G. (Eds.) Quelle & Meyer Verlag QM: Wiebelsheim, Germany, 2016; ISBN 9783494017303. [Google Scholar]
- Finck, P.; Heinze, S.; Raths, U.; Riecken, U.; Ssymank, A. Rote Liste der gefährdeten Biotoptypen Deutschlands. Natursch. Biol. Vielf. 2017, 156, 637. [Google Scholar]
- Kowarik, I. Humann agency in biological invasions: Secondary releases foster naturalisation and population expansion of alien plant species. Biol. Invasions 2003, 5, 293–312. [Google Scholar] [CrossRef]
- Donau-Silphie. Available online: https://www.donau-silphie.de/ (accessed on 9 September 2020).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Row Number in Tank | First Year (2018) | Second Year (2019) | Classification | ||
|---|---|---|---|---|---|
| WTD (cm) | WTD (cm) | Soil Water Content (%vol) in Depths | |||
| 5–25 cm | 40–60 cm | ||||
| 1 | 41 ± 7 | 40 ± 11 | 38 | 57 | wet |
| 2 | 51 ± 7 | 50 ± 11 | 31 | 48 | very moist |
| 3 | 61 ± 7 | 60 ± 11 | 26 | 40 | slightly moist |
| 4 | 71 ± 7 | 70 ± 11 | 22 | 33 | fresh |
| 5 | 81 ± 7 | 80 ± 11 | 18 | 28 | slightly dry |
| 6 | 91 ± 7 | 90 ± 11 | 15 | 23 | medium dry |
| 7 | 101 ± 7 | 101 ± 11 | 12 | 19 | rather dry |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ende, L.M.; Knöllinger, K.; Keil, M.; Fiedler, A.J.; Lauerer, M. Possibly Invasive New Bioenergy Crop Silphium perfoliatum: Growth and Reproduction Are Promoted in Moist Soil. Agriculture 2021, 11, 24. https://doi.org/10.3390/agriculture11010024
Ende LM, Knöllinger K, Keil M, Fiedler AJ, Lauerer M. Possibly Invasive New Bioenergy Crop Silphium perfoliatum: Growth and Reproduction Are Promoted in Moist Soil. Agriculture. 2021; 11(1):24. https://doi.org/10.3390/agriculture11010024
Chicago/Turabian StyleEnde, L. Marie, Katja Knöllinger, Moritz Keil, Angelika J. Fiedler, and Marianne Lauerer. 2021. "Possibly Invasive New Bioenergy Crop Silphium perfoliatum: Growth and Reproduction Are Promoted in Moist Soil" Agriculture 11, no. 1: 24. https://doi.org/10.3390/agriculture11010024
APA StyleEnde, L. M., Knöllinger, K., Keil, M., Fiedler, A. J., & Lauerer, M. (2021). Possibly Invasive New Bioenergy Crop Silphium perfoliatum: Growth and Reproduction Are Promoted in Moist Soil. Agriculture, 11(1), 24. https://doi.org/10.3390/agriculture11010024