
Effects of Grass Silage Additive Type and Barley Grain Preservation Method on Rumen Fermentation, Microbial Community and Milk Production of Dairy Cows

 ,
,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Experimental Feeds
2.2. Animal Care, Experimental Design, and Treatments
2.3. Sampling Procedures
2.4. Analytical Methods and Calculations
2.5. Statistical Analyses
3. Results
4. Discussion
4.1. Feed Characteristics
4.2. Rumen Fermentation
4.3. Rumen Bacterial Community
4.4. Feed Intake and Production Responses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilkinson, J.M.; Rinne, M. Highlights of progress in silage conservation and future perspectives. Grass Forage Sci. 2018, 73, 40–52. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991; p. 340. [Google Scholar]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Heikkilä, T.; Toivonen, V.; Huhtanen, P. Effects of and interactions between the extent of silage fermentation and protein supplementation in lactating dairy cows. Agric. Food Sci. Finl. 1998, 7, 329–343. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Jaakkola, S.; Huhtanen, P. Effect of forage conservation method, concentrate level and propylene glycol on diet digestibility, rumen fermentation, blood metabolite concentrations and nutrient utilisation of dairy cows. Anim. Feed Sci. Technol. 2002, 97, 1–21. [Google Scholar] [CrossRef]
- Jaakkola, S.; Kaunisto, V.; Huhtanen, P. Volatile fatty acid proportions and microbial protein synthesis in the rumen of cattle receiving grass silage ensiled with different rates of formic acid. Grass Forage Sci. 2006, 61, 282–292. [Google Scholar] [CrossRef]
- Huhtanen, P.; Rinne, M.; Nousiainen, J. Evaluation of the factors affecting silage intake of dairy cows: A revision of the relative silage dry-matter intake index. Animal 2007, 1, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Jokiniemi, T.; Jaakkola, S.; Turunen, M.; Ahokas, J. Energy consumption in different grain preservation methods. Agron. Res. 2014, 12, 81–94. Available online: http://agronomy.emu.ee/vol121/2014_1_9_b5.pdf (accessed on 29 November 2021).
- Franco, M.; Stefański, T.; Jalava, T.; Kuoppala, K.; Huuskonen, A.; Rinne, M. Fermentation quality and aerobic stability of low moisture-crimped wheat grains manipulated by organic acid-based additives. J. Agric. Sci. 2019, 157, 245–253. [Google Scholar] [CrossRef]
- Ferland, M.-C.; Guesthier, M.-A.; Cue, R.I.; Lacroix, R.; Burgos, S.A.; Lefebvre, D.; Wade, K.M. Effect of feeding system and grain source on lactation characteristics and milk components in dairy cattle. J. Dairy Sci. 2018, 101, 8572–8585. [Google Scholar] [CrossRef]
- Ferraretto, L.F.; Crump, P.M.; Shaver, R.D. Effect of cereal grain type and corn grain harvesting and processing methods on intake, digestion, and milk production by dairy cows through a meta-analysis. J. Dairy Sci. 2013, 96, 533–550. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.S.; Ying, Y. Effects of corn grain endosperm type and conservation method on feed intake, feeding behavior, and productive performance of lactating dairy cows. J. Dairy Sci. 2021, 104, 7604–7616. [Google Scholar] [CrossRef] [PubMed]
- Torres, R.N.S.; Ghedini, C.P.; Coelho, L.M.; Ezequiel, J.M.B.; Almeida Júnior, G.A.; Almeida, M.T.C. Meta-analysis of the effects of silage additives on high-moisture grain silage quality and performance of dairy cows. Livest. Sci. 2021, 251, 104618. [Google Scholar] [CrossRef]
- Jaakkola, S.; Saarisalo, E.; Kangasniemi, R. Ensiled high moisture barley or dry barley in the grass silage-based diet of dairy cows. In Silage Production and Utilisation: A Satellite Workshop of the XXth International Grassland Congress; Park, R.S., Stronge, M.D., Eds.; Wageningen Academic Publishers: Belfast, Northern Ireland, 2005; p. 184. [Google Scholar]
- Adler, S.A.; Randby, Å.T. The effect of preservation method of barley, maturity of gras silage, and type of protein supplement on sensory milk quality in organic farming. J. Anim. Feed Sci. 2007, 16 (Suppl. 1), 75–78. [Google Scholar] [CrossRef] [Green Version]
- Pettersson, T.; Bernes, G.; Martinsson, K. Ensiled rolled or dried barley drain and different levels of grass silage and hay to dairy cows. Swed. J. Agric. Res. 1998, 28, 99–109. [Google Scholar]
- Savonen, O.; Franco, M.; Stefanski, T.; Mäntysaari, P.; Kuoppala, K.; Rinne, M. Grass silage pulp as a dietary component for high yielding dairy cows. Animal 2020, 14, 1472–1480. [Google Scholar] [CrossRef]
- Salo, M.-L.; Salmi, M. Determination of starch by the amyloglucosidase method. J. Sci. Agric. Soc. 1968, 40, 38–45. [Google Scholar] [CrossRef]
- Weissbach, F.; Schmidt, L.; Hein, E. Method of anticipation of the run of fermentation in silage making based on the chemical composition of green fodder. In Proceedings of the 12th International Grassland Congress, Moscow, Russia, 11–30 June 1974; Russian Academy of Agricultural Sciences: Moscow, Russia, 1974; pp. 663–673. [Google Scholar]
- Rinne, M.; Franco, M.; Jalava, T.; Järvenpää, E.; Kahala, M.; Blasco, L.; Siljander-Rasi, H.; Kuoppala, K. Carrot by-product fermentation quality and aerobic spoilage could be modified with silage additives. Agric. Food Sci. 2019, 28, 59–69. [Google Scholar] [CrossRef]
- Abrahamse, P.A.; Vlaeminck, B.; Tamminga, S.; Dijkstra, J. The effect of silage and concentrate type on intake behavior, rumen function, and milk production in dairy cows in early and late lactation. J. Dairy Sci. 2008, 91, 4778–4792. [Google Scholar] [CrossRef]
- Rius, A.G.; Kittelmann, S.; Macdonald, K.A.; Waghorn, G.C.; Janssen, P.H.; Sikkema, E. Nitrogen metabolism and rumen microbial enumeration in lactating cows with divergent residual feed intake fed high-digestibility pasture. J. Dairy Sci. 2012, 95, 5024–5034. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucl. Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Seedorf, H.; Kittelmann, S.; Henderson, G.; Janssen, P.H. RIM-DB: A taxonomic framework for community structure analysis of methanogenic archaea from the rumen and other intestinal environments. PeerJ 2014, 2, e494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luke. Feed Tables and Nutrient Requirements. Natural Resources Institute Finland (Luke), Helsinki, Finland. 2022. Available online: https://www.luke.fi/feedtables (accessed on 4 January 2022).
- Sjaunja, L.O.; Bævre, L.; Junkkarinen, L.; Pedersen, J.; Setälä, J. A Nordic proposal for an energy corrected milk (ECM) formula. In Performance Recording of Animals: 27th Biennial Session of the International Committee for Animal Recording; EAAP Publication No. 50. Pudoc: Wageningen, The Netherlands, 1990; pp. 156–192. [Google Scholar]
- Xu, S.; Yu, G. MicrobiotaProcess: An R Package for Analysis, Visualization and Biomarker Discovery of Microbiome. R Package Version 1.6.1, 2021. Available online: https://github.com/YuLab-SMU/MicrobiotaProcess/ (accessed on 9 December 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6, 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 9 December 2021).
- Nadeau, E.M.G.; Russell, J.R.; Buxton, D.R. Intake, digestibility, and composition of orchardgrass and alfalfa silages treated with cellulase, inoculant, and formic acid fed to lambs. J. Anim. Sci. 2000, 78, 2980–2989. [Google Scholar] [CrossRef]
- Franco, M.; Pirttiniemi, J.; Tapio, I.; Rinne, M. Modulation of bacterial communities of grass silage by additives, compaction and soil contamination. In Proceedings of the 18th International Symposium Forage Conservation, Brno, Czech Republic, 13–16 August 2019; pp. 104–105, ISBN 978-80-7509-670-8. Available online: http://www.isfc.eu/18ISFC_2019.pdf (accessed on 30 November 2021).
- Sousa, D.O.; Hansen, H.H.; Nussio, L.G.; Nadeau, E. Effects of wilting and ensiling with or without additive on protein quality and fermentation of a lucerne-white clover mixture. Anim. Feed Sci. Technol. 2019, 258, 114301. [Google Scholar] [CrossRef]
- Saarisalo, E.; Skyttä, E.; Jaakkola, S. Effects of wilted grass silages varying in fermentation quality on rumen fermentation of dairy cows. J. Anim. Feed Sci. 2004, 13 (Suppl. 1), 199–202. [Google Scholar] [CrossRef]
- Halmemies-Beauchet-Filleau, A.; Kairenius, P.; Ahvenjärvi, S.; Crosley, L.K.; Muetzel, S.; Huhtanen, P.; Vanhatalo, A.; Toivonen, V.; Wallace, R.J.; Shingfield, K.J. Effects of forage conservation method on ruminal lipid metabolism and microbial ecology in lactating cows fed diets containing a 60:40 forage to concentrate ratio. J. Dairy Sci. 2013, 96, 2428–2447. [Google Scholar] [CrossRef]
- Jaakkola, S.; Rinne, M.; Heikkilä, T.; Toivonen, V.; Huhtanen, P. Effects of restriction of silage fermentation with formic acid on milk production. Agric. Food Sci. 2006, 15, 200–218. [Google Scholar] [CrossRef]
- Seppälä, A.; Heikkilä, T.; Mäki, M.; Rinne, M. Effects of additives on the fermentation and aerobic stability of grass silages and total mixed rations. Grass Forage Sci. 2016, 71, 458–471. [Google Scholar] [CrossRef]
- Rinne, M.; Franco, M.; Kuoppala, K.; Seppälä, A.; Jalava, T. Response to total mixed ration stabilizers depends on feed quality. In Proceedings of the XVIII International Silage Conference, Bonn, Germany, 24–26 July 2018; 2018; pp. 538–539. Available online: https://www.isc2018.de (accessed on 4 January 2022).
- Yang, H.E.; Zotti, C.A.; McKinnon, J.J.; McAllister, T.A. Lactobacilli are prominent members of the microbiota involved in the ruminal digestion of barley and corn. Front. Microbiol. 2018, 9, 718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huuskonen, A.; Rinne, M.; Manni, K. Effects of different barley grain preservation techniques on intake, growth and carcass traits of finishing dairy bulls fed grass silage-based rations. J. Agric. Sci. 2021, 158, 748–755. [Google Scholar] [CrossRef]
- Allen, M.S.; Ying, Y. Effects of corn grain endosperm type and conservation method on site of digestion, ruminal digestion kinetics, and flow of nitrogen fractions to the duodenum in lactating dairy cows. J. Dairy Sci. 2021, 104, 7617–7629. [Google Scholar] [CrossRef] [PubMed]
- Huhtanen, P.; Rinne, M. Effects of increasing the milk yield of dairy cows on milk composition. J. Anim. Feed Sci. 2007, 16 (Suppl. 1), 42–58. [Google Scholar] [CrossRef]






{kind=link}
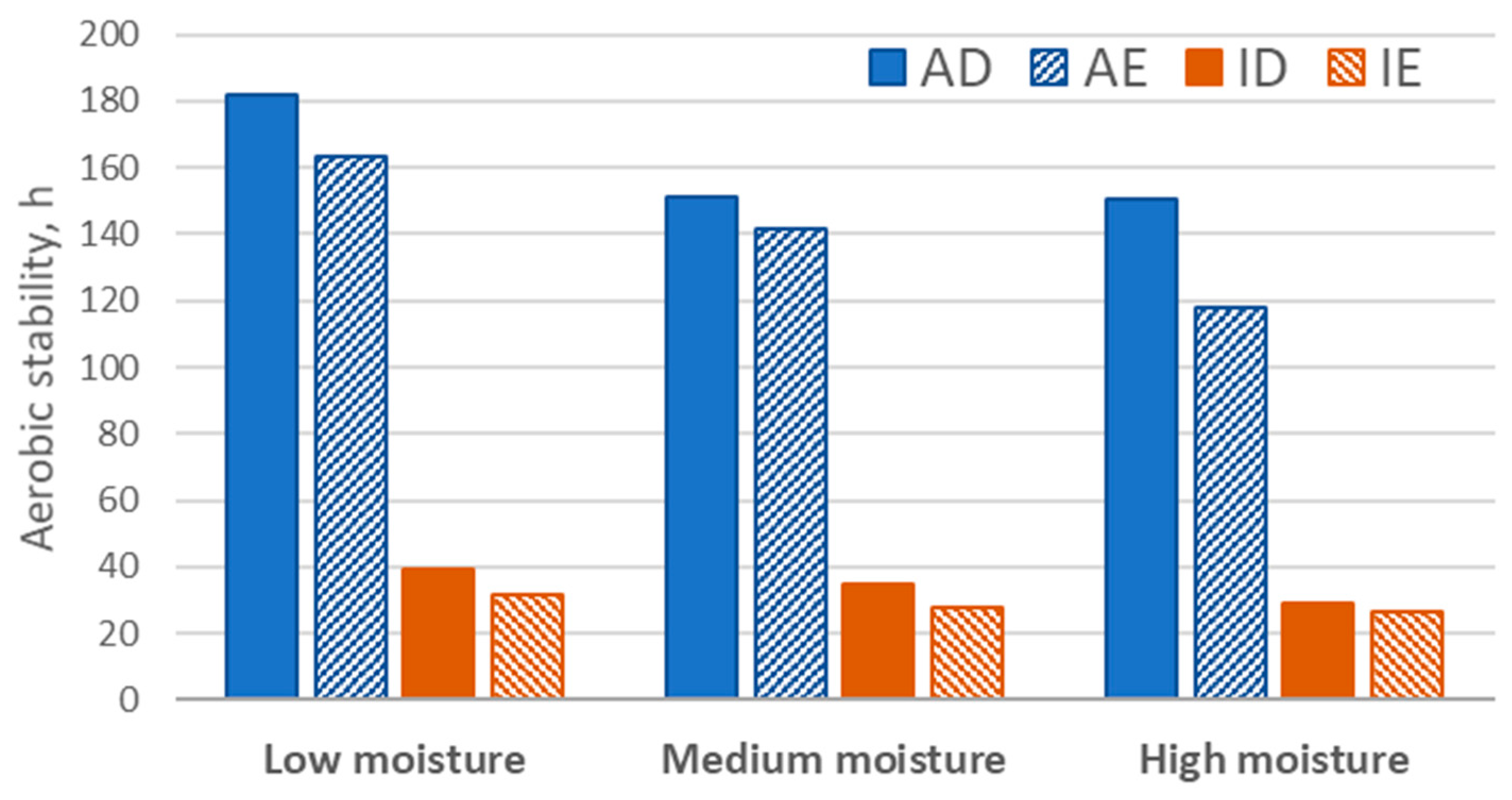{kind=link}
{kind=link}
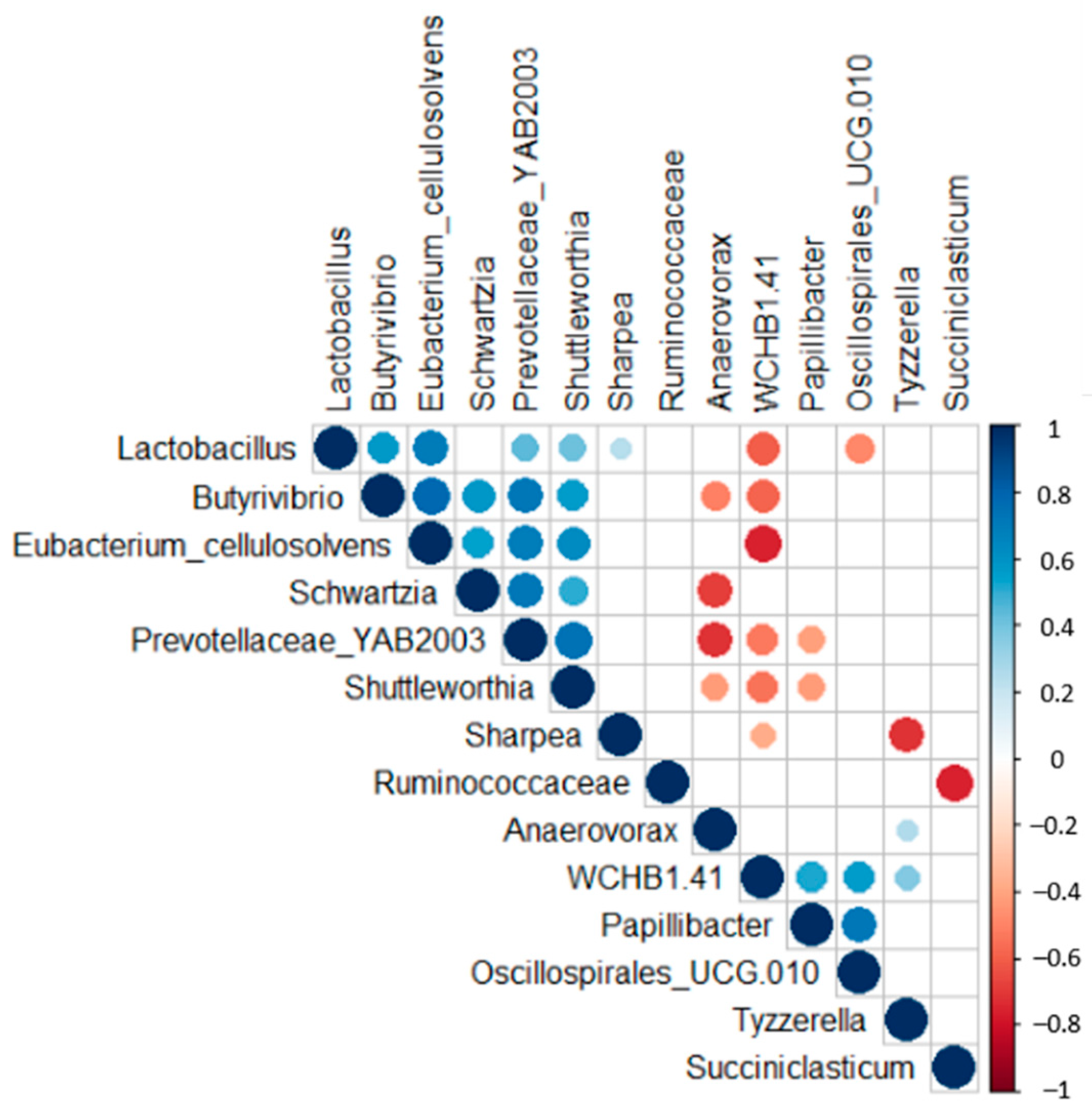{kind=link}
{kind=link}
| Fresh Forage | Grass Silage | Concentrate Feeds | Experimental Total Mixed Rations 3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Acid | Inoculant | Acid | Inoculant | MixC 1 | Dry Barley | Ensiled Barley | MPC 2 | AD | AE | ID | IE | |
| Number of samples | 7 | 5 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 | 4 |
| Dry matter (DM), g/kg | 259 ± 12.7 | 258 ± 13.9 | 271 ± 6.1 | 260 ± 11.4 | 871 ± 3.5 | 877 ± 4.4 | 730 ± 4.9 | 877 ± 0.6 | 401 ± 6.5 | 397 ± 6.2 | 396 ± 11.4 | 389 ± 11.3 |
| Buffering capacity, g lactic acid/100 g DM | 6.2 ± 0.30 | 7.8 + 0.33 | ||||||||||
| In DM, g/kg | ||||||||||||
| Ash | 76 ± 2.1 | 76 ± 2.1 | 78 ± 1.0 | 81 ± 3.4 | 108 ± 3.2 | 30 ± 1.2 | 32 ± 0.3 | 81 ± 15.3 | 76 ± 0.6 | 76 ± 0.8 | 77 ± 1.7 | 77 ± 1.8 |
| Crude protein | 137 ± 8.7 | 135 ± 8.6 | 149 ± 2.2 | 150 ± 5.8 | 240 ± 3.7 | 136 ± 2.4 | 132 ± 2.1 | 184 ± 8.0 | 170 ± 1.6 | 169 ± 1.6 | 171 ± 3.4 | 170 ± 3.4 |
| Ether extract | 36.6 ± 4.6 | 30.6 ± 1.0 | 31.3 ± 0.7 | 28.4 ± 3.9 | ||||||||
| Water soluble carbohydrates | 141 ± 14.2 | 128 ± 15.4 | 26.0 ± 4.0 | 40.5 ± 10.1 | ||||||||
| Neutral detergent fibre | 538 ± 16.5 | 565 ± 17.6 | 522 ± 17.6 | 520 ± 17.5 | 251 ± 8.3 | 191 ± 9.1 | 193 ± 8.6 | 204 ± 4.4 | 379 ± 9.0 | 379 ± 12.2 | 375 ± 9.7 | 375 ± 12.8 |
| Starch | 156 ± 7.6 | 605 ± 45.9 | 585 ± 18.3 | 363 ± 10.5 | 173 ± 8.7 | 170 ± 3.1 | 177 ± 8.1 | 173 ± 3.2 | ||||
| Fermentation profile, g/kg DM | ||||||||||||
| pH | 3.99 ± 0.307 | 3.97 ± 0.029 | 4.42 ± 0.11 | |||||||||
| Ammonia N, g/kg N | 35.8 ± 2.06 | 42.8 ± 4.90 | 19.9 ± 3.1 | |||||||||
| Ethanol | 6.6 ± 0.75 | 6.6 ± 0.84 | 4.6 ± 0.7 | |||||||||
| Lactic acid | 88.8 ± 4.69 | 99.8 ± 7.76 | 14.5 ± 1.4 | |||||||||
| Acetic acid | 16.9 ± 1.64 | 12.5 ± 6.18 | 8.45 ± 1.7 | |||||||||
| Propionic acid | 2.36 ± 0.102 4 | 0.63 ± 0.307 | 0.05 ± 0.01 | |||||||||
| Butyric acid | 0.36 ± 0.069 | 0.83 ± 0.815 | 0.003 ± 0.01 | |||||||||
| In vitro organic matter digestibility, g/kg | 793 ± 10.0 | 780 ± 10.7 | 793 ± 2.5 | 786 ± 5.8 | ||||||||
| Feed values | ||||||||||||
| Metabolisable energy, MJ/kg DM | 11.7 ± 0.04 | 11.6 ± 0.12 | 11.3 ± 0.03 | 12.9 ± 0.02 | 12.9 ± 0.003 | 12.1 ± 0.21 | 11.9 ± 0.02 | 11.9 ± 0.02 | 11.8 ± 0.06 | 11.8 ± 0.06 | ||
| Metabolisable protein, g/kg DM | 87 ± 0.5 | 86 ± 0.9 | 126 ± 1.2 | 110 ± 0.6 | 109 ± 0.5 | 117 ± 2.1 | 102 ± 0.4 | 102 ± 0.4 | 102 ± 0.5 | 102 ± 0.6 | ||
| Protein balance in the rumen, g/kg DM | 19 ± 1.7 | 21 ± 5.2 | 65 ± 0.3 | −23 ± 0.7 | −26 ± 0.5 | 18 ± 0.5 | 22 ± 1.2 | 21 ± 1.2 | 23 ± 2.9 | 22 ± 2.9 | ||
| Silage DM intake index 5 | 108 ± 0.8 | 104 ± 2.5 | ||||||||||
| Microbial counts 6, CFU/g | ||||||||||||
| Yeasts | 1.1 × 105 | 2.0 × 102 | 1.2 × 107 | 3.6 × 102 | 3.3 × 102 | 1.6 × 105 | ||||||
| Moulds | 3.8 × 104 | 1.8 × 102 | 1.0 × 102 | 3.4 × 102 | 3.3 × 102 | 3.7 × 105 | ||||||
| Aerobic bacteria | 3.0 × 107 | 2.1 × 105 | 1.5 × 107 | 1.1 × 105 | 8.1 × 104 | 5.8 × 107 | ||||||
| Lactic acid bacteria | Not analysed | 4.1 × 106 | 7.4 × 107 | 2.9 × 104 | 6.3 × 103 | 2.2 × 108 | ||||||
| Silage Additive (S): | Acid | Inoculant | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Barley Preservation (B): | Dry | Ensiled | Dry | Ensiled | S | B | S × B | |
| Total dry matter | 27.8 | 27.7 | 27.3 | 27.0 | 0.72 | 0.005 | 0.34 | 0.61 |
| Organic matter | 25.7 | 25.6 | 25.2 | 24.9 | 0.67 | 0.003 | 0.32 | 0.62 |
| Crude protein | 4.74 | 4.70 | 4.67 | 4.60 | 0.140 | 0.018 | 0.11 | 0.65 |
| Neutral detergent fibre | 10.5 | 10.4 | 10.2 | 10.0 | 0.26 | <0.001 | 0.34 | 0.59 |
| Starch | 4.90 | 4.80 | 4.92 | 4.78 | 0.148 | 0.91 | 0.007 | 0.61 |
| Metabolisable energy 1, MJ | 302 | 301 | 296 | 292 | 7.5 | 0.001 | 0.34 | 0.62 |
| Metabolisable protein | 2.85 | 2.84 | 2.80 | 2.76 | 0.077 | 0.004 | 0.27 | 0.63 |
| Protein balance in the rumen | 0.612 | 0.590 | 0.623 | 0.600 | 0.0366 | 0.17 | 0.004 | 0.88 |
| Silage Additive (S): | Acid | Inoculant | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| Barley Preservation (B): | Dry | Ensiled | Dry | Ensiled | Time | S | B | S × B | |
| Ammonia N, mg/dL | 7.4 | 11.1 | 9.3 | 10.4 | 1.34 | <0.001 | 0.55 | 0.012 | 0.17 |
| pH | 6.05 | 6.06 | 6.10 | 6.11 | 0.142 | 0.010 | 0.54 | 0.83 | 0.99 |
| VFA 1, mmol/L | 122 | 123 | 124 | 120 | 3.8 | <0.001 | 0.90 | 0.63 | 0.33 |
| In total VFA, mmol/mol | |||||||||
| Acetic acid | 618 | 620 | 624 | 630 | 10.6 | <0.001 | 0.041 | 0.41 | 0.51 |
| Propionic acid | 204 | 207 | 210 | 198 | 5.3 | <0.001 | 0.55 | 0.55 | 0.022 |
| Butyric acid | 142 | 136 | 130 | 134 | 6.5 | <0.001 | 0.066 | 0.47 | 0.001 |
| Isobutyric acid | 6.1 | 6.7 | 6.6 | 6.8 | 0.26 | 0.31 | 0.20 | 0.002 | 0.16 |
| Valeric acid | 15.6 | 15.3 | 15.4 | 14.7 | 0.57 | <0.001 | 0.19 | 0.14 | 0.48 |
| Isovaleric acid | 7.2 | 9.0 | 8.7 | 9.1 | 0.66 | <0.001 | 0.017 | 0.16 | 0.034 |
| Caproic acid | 6.2 | 6.7 | 6.1 | 6.7 | 0.38 | <0.001 | 0.66 | 0.15 | 0.76 |
| NGR 2 | 3.97 | 3.86 | 3.80 | 4.05 | 0.108 | 0.002 | 0.86 | 0.64 | 0.010 |
| Silage Additive (S): | Acid | Inoculant | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| Barley Preservation (B): | Dry | Ensiled | Dry | Ensiled | S | B | S × B | |
| Production per day | ||||||||
| Milk, kg | 39.6 | 39.0 | 39.5 | 38.9 | 2.30 | 0.89 | 0.086 | 0.93 |
| Energy corrected milk, kg | 43.3 | 43.1 | 42.7 | 42.6 | 1.45 | 0.13 | 0.76 | 0.88 |
| Fat, g | 1803 | 1817 | 1759 | 1783 | 53.1 | 0.079 | 0.38 | 0.82 |
| Protein, g | 1505 | 1478 | 1493 | 1469 | 52.7 | 0.43 | 0.061 | 0.91 |
| Lactose, g | 1786 | 1763 | 1791 | 1762 | 105.0 | 0.88 | 0.12 | 0.84 |
| Milk composition, g/kg | ||||||||
| Fat | 45.8 | 46.9 | 45.0 | 46.1 | 1.78 | 0.17 | 0.060 | 0.99 |
| Protein | 38.2 | 38.1 | 38.1 | 37.9 | 1.17 | 0.30 | 0.22 | 0.77 |
| Lactose | 45.1 | 45.2 | 45.3 | 45.3 | 0.29 | 0.20 | 0.85 | 0.37 |
| Total solids | 139 | 140 | 138 | 139 | 3.0 | 0.12 | 0.080 | 0.84 |
| Urea, mg/100 mL | 23.3 | 25.5 | 24.7 | 24.5 | 0.99 | 0.74 | 0.057 | 0.021 |
| Somatic cells, 1000/mL | 95 | 67 | 89 | 75 | 23.9 | 0.97 | 0.24 | 0.70 |
| Efficiency of milk production | ||||||||
| NUE 1 | 0.312 | 0.308 | 0.313 | 0.313 | 0.0082 | 0.22 | 0.48 | 0.45 |
| kg ECM 2/kg DM 3 intake | 1.56 | 1.56 | 1.56 | 1.58 | 0.031 | 0.24 | 0.48 | 0.38 |
| kg ECM/MJ ME 4 intake | 0.143 | 0.143 | 0.144 | 0.146 | 0.0030 | 0.10 | 0.51 | 0.41 |
| Energy balance 5 | 9.56 | 9.47 | 6.30 | 3.41 | 4.896 | 0.013 | 0.41 | 0.44 |
| Body weight | ||||||||
| Mean, kg | 695 | 694 | 694 | 694 | 16.4 | 0.69 | 0.89 | 0.61 |
| Change, kg/week | 2.81 | 0.77 | 1.12 | 1.26 | 1.163 | 0.47 | 0.26 | 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinne, M.; Franco, M.; Tapio, I.; Stefański, T.; Bayat, A.-R.; Mäntysaari, P. Effects of Grass Silage Additive Type and Barley Grain Preservation Method on Rumen Fermentation, Microbial Community and Milk Production of Dairy Cows. Agriculture 2022, 12, 266. https://doi.org/10.3390/agriculture12020266
Rinne M, Franco M, Tapio I, Stefański T, Bayat A-R, Mäntysaari P. Effects of Grass Silage Additive Type and Barley Grain Preservation Method on Rumen Fermentation, Microbial Community and Milk Production of Dairy Cows. Agriculture. 2022; 12(2):266. https://doi.org/10.3390/agriculture12020266
Chicago/Turabian StyleRinne, Marketta, Marcia Franco, Ilma Tapio, Tomasz Stefański, Ali-Reza Bayat, and Päivi Mäntysaari. 2022. "Effects of Grass Silage Additive Type and Barley Grain Preservation Method on Rumen Fermentation, Microbial Community and Milk Production of Dairy Cows" Agriculture 12, no. 2: 266. https://doi.org/10.3390/agriculture12020266
APA StyleRinne, M., Franco, M., Tapio, I., Stefański, T., Bayat, A.-R., & Mäntysaari, P. (2022). Effects of Grass Silage Additive Type and Barley Grain Preservation Method on Rumen Fermentation, Microbial Community and Milk Production of Dairy Cows. Agriculture, 12(2), 266. https://doi.org/10.3390/agriculture12020266