Magnetic Immobilization of Pichia pastoris Cells for the Production of Recombinant Human Serum Albumin


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Synthesis of Nanoparticles
2.3. Nanoparticles Characterization
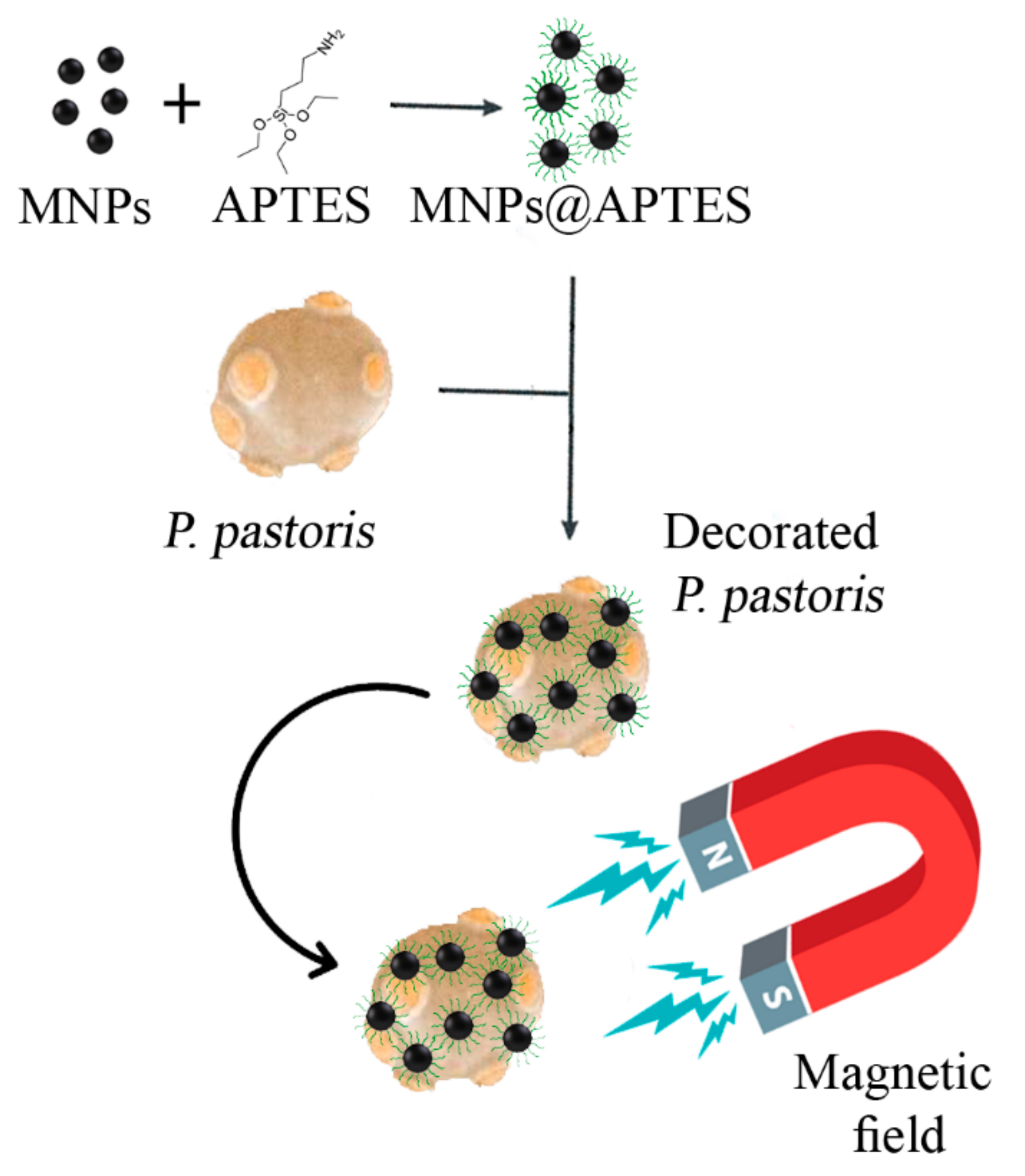
2.4. Cell Immobilization
2.5. Recombinant Protein Production
2.6. Nanoparticles Interaction with the Cells
3. Results
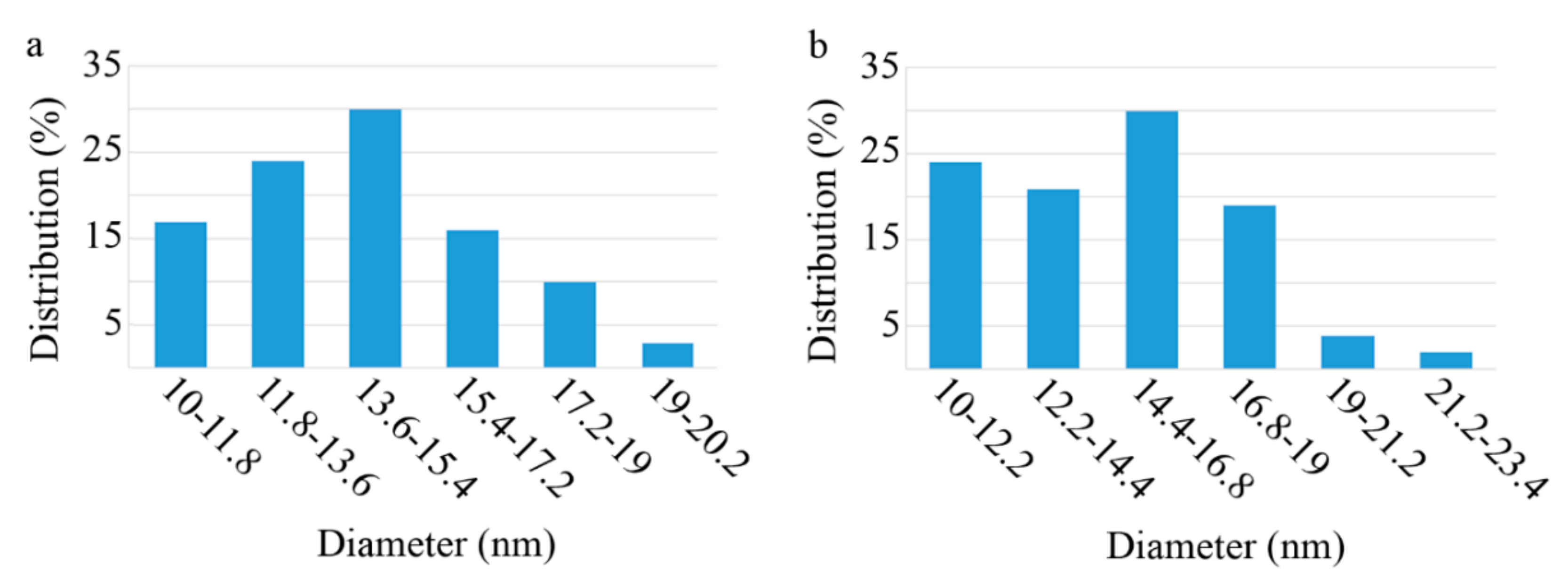
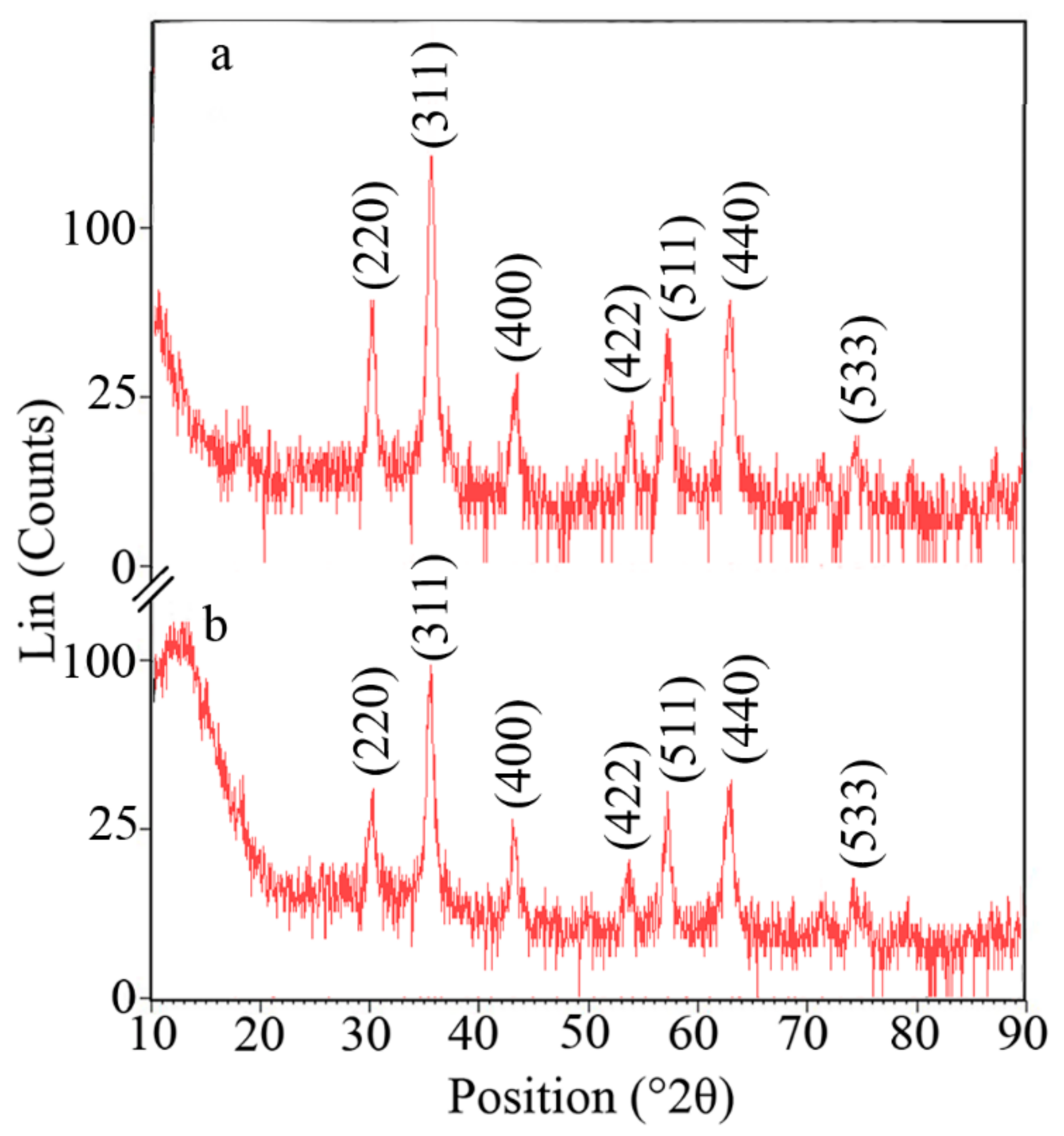
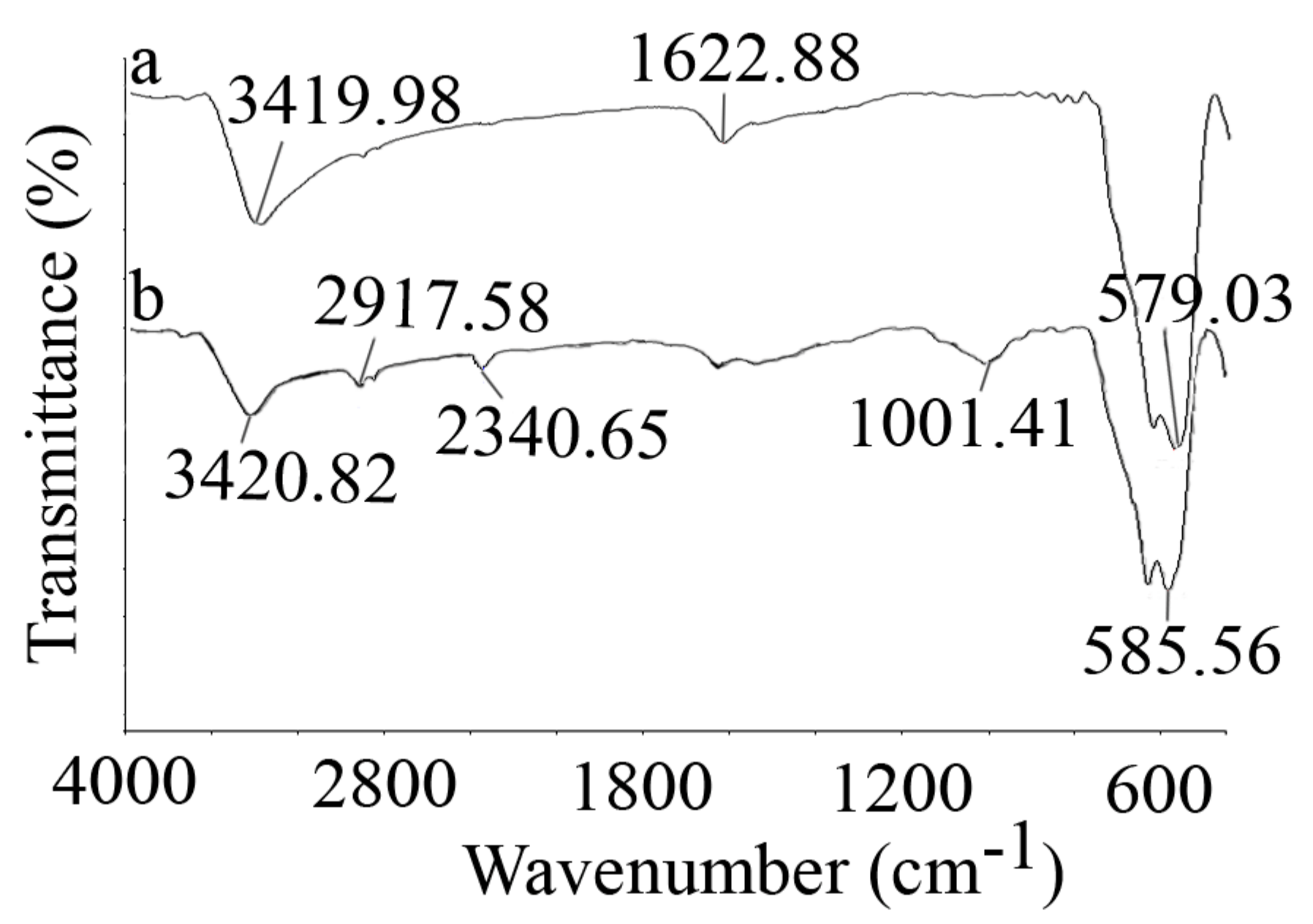
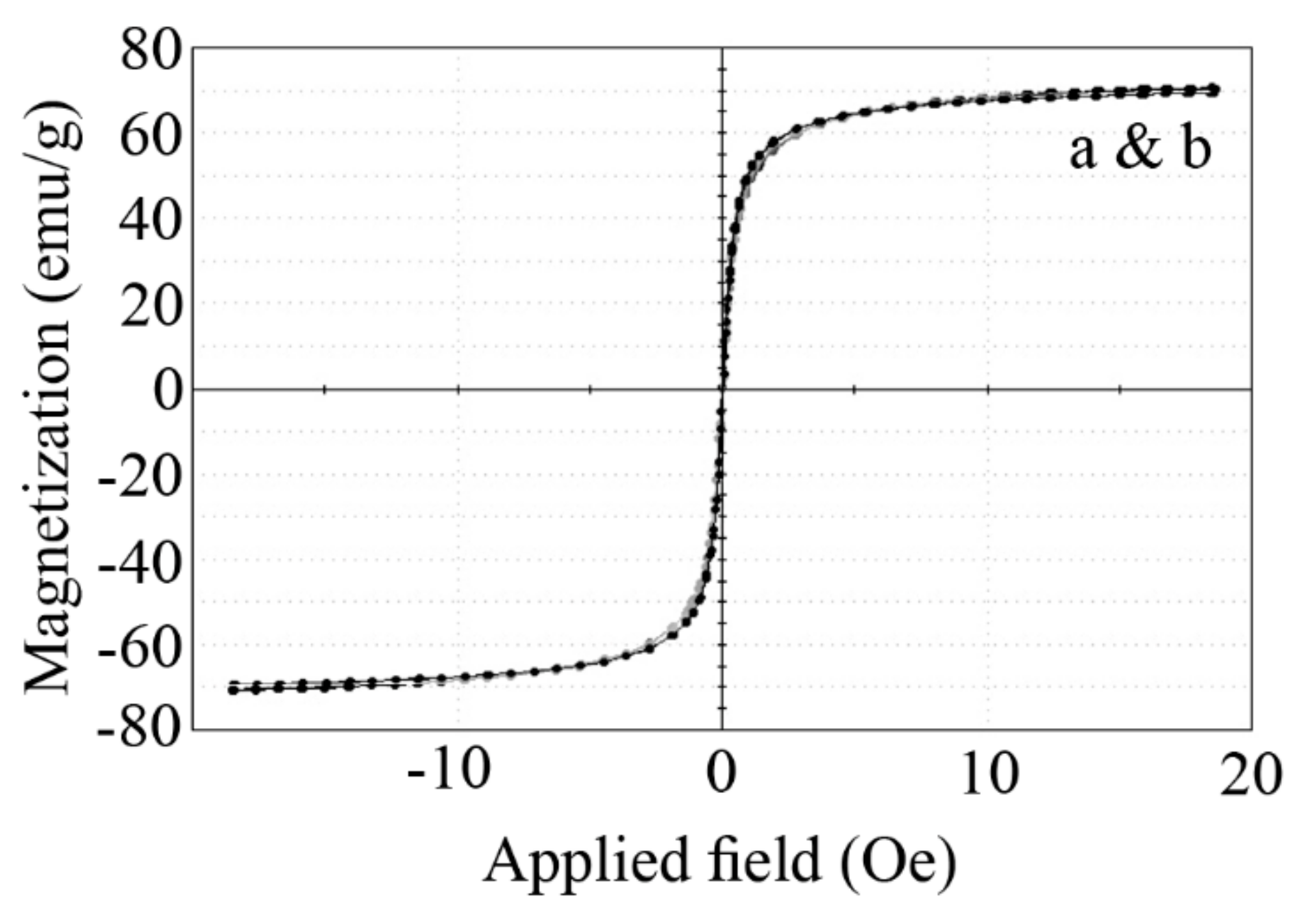
3.1. Nanoparticle Characterization
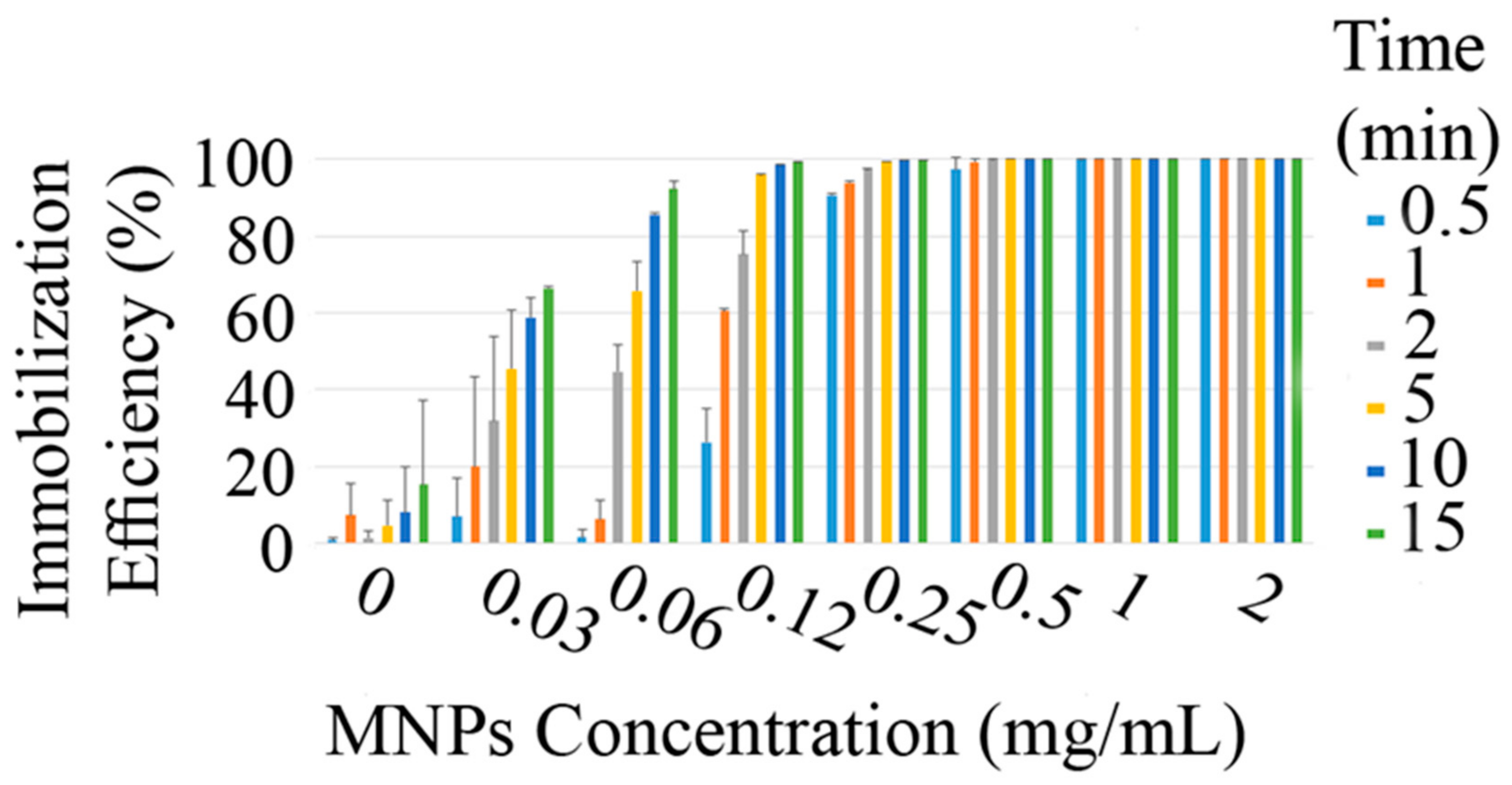
3.2. Cell Immobilization
3.3. Recombinant HSA Production
3.4. Nanoparticles’ Interaction with Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Daly, R.; Hearn, M.T. Expression of heterologous proteins in Pichia pastoris: A useful experimental tool in protein engineering and production. J. Mol. Recognit. Interdiscip. J. 2005, 18, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Naseem, M.U.; Ahmed, N.; Khan, M.A.; Tahir, S.; Zafar, A.U. Production of potent long-lasting consensus interferon using albumin fusion technology in Pichia pastoris expression system. Protein Expr. Purif. 2019, 166. [Google Scholar] [CrossRef] [PubMed]
- Bobik, T.; Popov, R.Y.; Aliev, T.; Mokrushina, Y.A.; Shamborant, O.; Khurs, E.; Knorre, V.; Smirnov, I.; Gabibov, A. Production of recombinant human transferrin in eukaryotic Pichia pastoris expression system. Bull. Exp. Biol. Med. 2019, 167, 335–338. [Google Scholar] [CrossRef] [PubMed]
- Pichia Produced Products on the Market. Available online: https://pichia.com/science-center/commercialized-products/ (accessed on 26 October 2019).
- Iglesias-Figueroa, B.; Valdiviezo-Godina, N.; Siqueiros-Cendón, T.; Sinagawa-García, S.; Arévalo-Gallegos, S.; Rascón-Cruz, Q. High-level expression of recombinant bovine lactoferrin in Pichia pastoris with antimicrobial activity. Int. J. Mol. Sci. 2016, 17, 902. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, A.; Hosseini, S.N.; Karimi, A.; Aghdasinia, H.; Mianroodi, R.A. Enhancing the efficiency of recombinant hepatitis B surface antigen production in Pichia pastoris by employing continuous fermentation. Biochem. Eng. J. 2019, 141, 112–119. [Google Scholar] [CrossRef]
- Azadi, S.; Sadjady, S.K.; Mortazavi, S.A.; Naghdi, N.; Mahboubi, A.; Solaimanian, R. Bioprocess and downstream optimization of recombinant human growth hormone in Pichia pastoris. Res. Pharm. Sci. 2018, 13, 222. [Google Scholar]
- Yang, P.; Jiang, S.; Wu, Y.; Hou, Z.; Zheng, Z.; Cao, L.; Du, M.; Jiang, S. Recombinant expression of Serratia marcescens outer membrane phospholipase A (A1) in Pichia pastoris and immobilization with graphene oxide-based Fe3O4 nanoparticles for rapeseed oil degumming. Front. Microbiol. 2019, 10, 334. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ling, C.; Chen, Y.; Jiang, X.; Chen, G.-Q. Microbial engineering for easy downstream processing. Biotechnol. Adv. 2019, 37. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A. Downstream processing for biopharmaceuticals recovery. In Pharmaceuticals from Microbes; Springer: Berlin, Germany, 2019; pp. 163–190. [Google Scholar] [CrossRef]
- Thipayarat, A. Production of Human Serum Albumin by Immobilized Yeast. Available online: https://surface.syr.edu/bce_etd/25 (accessed on 8 December 2019).
- Li, Y.-M.; Gao, J.-Q.; Pei, X.-Z.; Du, C.; Fan, C.; Yuan, W.-J.; Bai, F.-W. Production of L-alanyl-L-glutamine by immobilized Pichia pastoris GS115 expressing α-amino acid ester acyltransferase. Microb. Cell Factories 2019, 18, 27. [Google Scholar] [CrossRef] [PubMed]
- Duff, S.J.; Murray, W.D. Comparison of free and immobilized Pichia pastoris cells for conversion of ethanol to acetaldehyde. Biotechnol. Bioeng. 1988, 31, 790–795. [Google Scholar] [CrossRef] [PubMed]
- Martínez, D.; Menéndez, C.; Echemendia, F.M.; Pérez, E.R.; Trujillo, L.E.; Sobrino, A.; Ramírez, R.; Quintero, Y.; Hernández, L. Complete sucrose hydrolysis by heat-killed recombinant Pichia pastoris cells entrapped in calcium alginate. Microb. Cell Factories 2014, 13, 87. [Google Scholar] [CrossRef] [Green Version]
- Maleknia, S.; Ahmadi, H.; Norouzian, D. Immobilization of Pichia pastoris cells containing alcohol oxidase activity. Iran. J. Microbiol. 2011, 3, 210. [Google Scholar]
- Ho, C.L.; Hwang, I.Y.; Loh, K.; Chang, M.W. Matrix-immobilized yeast for large-scale production of recombinant human lactoferrin. Med. Chem. Commun. 2015, 6, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Guan, C.; Wang, G.; Ji, J.; Wang, J.; Wang, H.; Tan, M. Bioencapsulation of living yeast (Pichia pastoris) with silica after transformation with lysozyme gene. J. Sol-Gel Sci. Technol. 2008, 48, 369–377. [Google Scholar] [CrossRef]
- Borovikova, D.; Rozenfelde, L.; Pavlovska, I.; Rapoport, A. Immobilisation increases yeast cells’ resistance to dehydration–rehydration treatment. J. Biotechnol. 2014, 184, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.Y.; Cantú, M.E.; Garcia-Reyes, B.; Garza-Gonzalez, M.T.; Meza-Escalante, E.R.; Serrano, D.; Alvarez, L.H. Biotransformation of 4-nitrophenol by co-immobilized Geobacter sulfurreducens and anthraquinone-2-sulfonate in barium alginate beads. Chemosphere 2019, 221, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Güereca, D.A.; del Pilar Sánchez-Saavedra, M. Growth and phosphorus removal by Synechococcus elongatus co-immobilized in alginate beads with Azospirillum brasilense. J. Appl. Phycol. 2016, 28, 1501–1507. [Google Scholar] [CrossRef]
- Ebrahiminezhad, A.; Varma, V.; Yang, S.; Berenjian, A. Magnetic immobilization of Bacillus subtilis natto cells for menaquinone-7 fermentation. Appl. Microbiol. Biotechnol. 2016, 100, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Ebrahiminezhad, A.; Varma, V.; Yang, S.; Ghasemi, Y.; Berenjian, A. Synthesis and application of amine functionalized iron oxide nanoparticles on menaquinone-7 fermentation: A step towards process intensification. Nanomaterials 2015, 6, 1. [Google Scholar] [CrossRef]
- Ranmadugala, D.; Ebrahiminezhad, A.; Manley-Harris, M.; Ghasemi, Y.; Berenjian, A. Magnetic immobilization of bacteria using iron oxide nanoparticles. Biotechnol. Lett. 2017, 40, 237–248. [Google Scholar] [CrossRef]
- Ansari, F.; Grigoriev, P.; Libor, S.; Tothill, I.E.; Ramsden, J.J. DBT degradation enhancement by decorating Rhodococcus erythropolis IGST8 with magnetic Fe3O4 nanoparticles. Biotechnol. Bioeng. 2009, 102, 1505–1512. [Google Scholar] [CrossRef] [Green Version]
- Raee, M.J.; Ebrahiminezhad, A.; Gholami, A.; Ghoshoon, M.B.; Ghasemi, Y. Magnetic immobilization of recombinant E. coli producing extracellular asparaginase: An effective way to intensify downstream process. Sep. Sci. Technol. 2018, 53, 1397–1404. [Google Scholar] [CrossRef]
- Ranmadugala, D.; Ebrahiminezhad, A.; Manley-Harris, M.; Ghasemi, Y.; Berenjian, A. The effect of iron oxide nanoparticles on Bacillus subtilis biofilm, growth and viability. Process Biochem. 2017, 62, 231–240. [Google Scholar] [CrossRef]
- Taghizadeh, S.-M.; Jafari, S.; Ahmad-Kiadaliri, T.; Mobasher, M.A.; Lal, N.; Raee, M.J.; Berenjian, A.; Ghasemi, Y.; Ebrahiminezhad, A. Magnetic immobilisation as a promising approach against bacteriophage infection. Mater. Res. Express 2019. [Google Scholar] [CrossRef]
- Safarik, I.; Maderova, Z.; Pospiskova, K.; Baldikova, E.; Horska, K.; Safarikova, M. Magnetically responsive yeast cells: Methods of preparation and applications. Yeast 2015, 32, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Berovic, M.; Berlot, M.; Kralj, S.; Makovec, D. A new method for the rapid separation of magnetized yeast in sparkling wine. Biochem. Eng. J. 2014, 88, 77–84. [Google Scholar] [CrossRef]
- Safarik, I.; Pospiskova, K.; Maderova, Z.; Baldikova, E.; Horska, K.; Safarikova, M. Microwave-synthesized magnetic chitosan microparticles for the immobilization of yeast cells. Yeast 2015, 32, 239–243. [Google Scholar] [CrossRef]
- Ebrahiminezhad, A.; Ghasemi, Y.; Rasoul-Amini, S.; Barar, J.; Davaran, S. Preparation of novel magnetic fluorescent nanoparticles using amino acids. Colloids Surf. B 2013, 102, 534–539. [Google Scholar] [CrossRef]
- Li, Y.G.; Gao, H.S.; Li, W.L.; Xing, J.M.; Liu, H.Z. In situ magnetic separation and immobilization of dibenzothiophene-desulfurizing bacteria. Bioresour. Technol. 2009, 100, 5092–5096. [Google Scholar] [CrossRef]
- Feng, B.; Hong, R.; Wang, L.; Guo, L.; Li, H.; Ding, J.; Zheng, Y.; Wei, D. Synthesis of Fe3O4/APTES/PEG diacid functionalized magnetic nanoparticles for MR imaging. Colloids Surf. A Physicochem. Eng. Asp. 2008, 328, 52–59. [Google Scholar] [CrossRef]
- Mohapatra, S.; Pramanik, N.; Mukherjee, S.; Ghosh, S.K.; Pramanik, P. A simple synthesis of amine-derivatised superparamagnetic iron oxide nanoparticles for bioapplications. J. Mater. Sci. 2007, 42, 7566–7574. [Google Scholar] [CrossRef]
- Tang, T.; Fan, H.; Ai, S.; Han, R.; Qiu, Y. Hemoglobin (Hb) immobilized on amino-modified magnetic nanoparticles for the catalytic removal of bisphenol A. Chemosphere 2011, 83, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.C.; Fang, X.Z.; Zhou, Y.H.; Liang, H. Synthesis and characterization of 3-aminopropyltriethoxysilane-modified superparamagnetic magnetite nanoparticles. Chem. Lett. 2004, 33, 1468–1469. [Google Scholar] [CrossRef]
- Yamaura, M.; Camilo, R.; Sampaio, L.; Mac do, M.; Nakamura, M.; Toma, H. Preparation and characterization of (3-aminopropyl) triethoxysilane-coated magnetite nanoparticles. J. Magn. Magn. Mater. 2004, 279, 210–217. [Google Scholar] [CrossRef]
- Ozmen, M.; Can, K.; Arslan, G.; Tor, A.; Cengeloglu, Y.; Ersoz, M. Adsorption of Cu(II) from aqueous solution by using modified Fe3O4 magnetic nanoparticles. Desalination 2010, 254, 162–169. [Google Scholar] [CrossRef]
- Lin, L.; Wu, W.; Huang, J.; Sun, D.; Zhou, Y.; Wang, H.; Li, Q. Catalytic gold nanoparticles immobilized on yeast: From biosorption to bioreduction. Chem. Eng. J. 2013, 225, 857–864. [Google Scholar] [CrossRef]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taghizadeh, S.-M.; Ebrahiminezhad, A.; Ghoshoon, M.B.; Dehshahri, A.; Berenjian, A.; Ghasemi, Y. Magnetic Immobilization of Pichia pastoris Cells for the Production of Recombinant Human Serum Albumin. Nanomaterials 2020, 10, 111. https://doi.org/10.3390/nano10010111
Taghizadeh S-M, Ebrahiminezhad A, Ghoshoon MB, Dehshahri A, Berenjian A, Ghasemi Y. Magnetic Immobilization of Pichia pastoris Cells for the Production of Recombinant Human Serum Albumin. Nanomaterials. 2020; 10(1):111. https://doi.org/10.3390/nano10010111
Chicago/Turabian StyleTaghizadeh, Seyedeh-Masoumeh, Alireza Ebrahiminezhad, Mohammad Bagher Ghoshoon, Ali Dehshahri, Aydin Berenjian, and Younes Ghasemi. 2020. "Magnetic Immobilization of Pichia pastoris Cells for the Production of Recombinant Human Serum Albumin" Nanomaterials 10, no. 1: 111. https://doi.org/10.3390/nano10010111
APA StyleTaghizadeh, S. -M., Ebrahiminezhad, A., Ghoshoon, M. B., Dehshahri, A., Berenjian, A., & Ghasemi, Y. (2020). Magnetic Immobilization of Pichia pastoris Cells for the Production of Recombinant Human Serum Albumin. Nanomaterials, 10(1), 111. https://doi.org/10.3390/nano10010111