
Chitosan–Selenium Nanoparticle (Cs–Se NP) Foliar Spray Alleviates Salt Stress in Bitter Melon

 , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Preparation of Chitosan-Selenium Nanoparticles (Cs-Se NPs)
2.3. Plant Growth, Fruit Parameters, and Relative Water Content (RWC) in Leaves
2.4. Photosynthetic Pigments and Gas Exchange Capacity
2.5. Leaf Sodium and Potassium Content
2.6. Leaf Proline and Total Soluble Carbohydrates
2.7. Malonaldehyde (MDA) and Hydrogen Peroxide (H2O2) Content and Electrolyte Leakage (EL)
2.8. Antioxidant Enzymes
2.9. Fruit Quality and Biochemical Parameters
2.9.1. Total Phenols, Flavonoids, Ascorbic Acid, Anthocyanin, and Radical Scavenging Activity (DPPH)
2.9.2. Essential Oils and Associated Compounds
2.10. Statistical Analysis
3. Results and Discussion
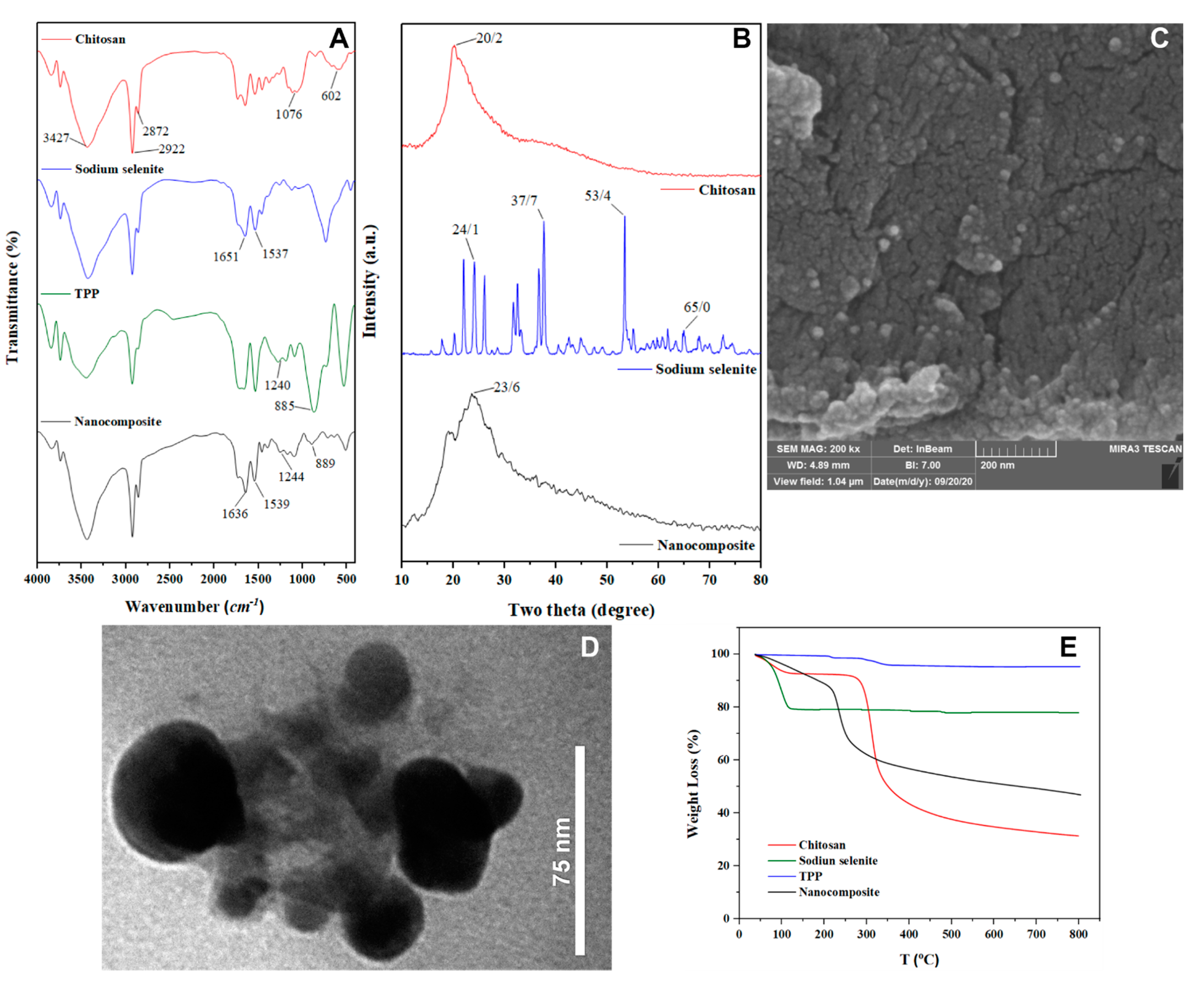
3.1. Synthesis and Characterization of Cs–Se NPs
3.2. Influence of Cs–Se NPs on Plant Growth, Fruiting, and RWC
3.3. Effects of Cs-Se NPs on Photosynthetic Pigments and Gas Exchange Capacity under Salinity Stress Conditions
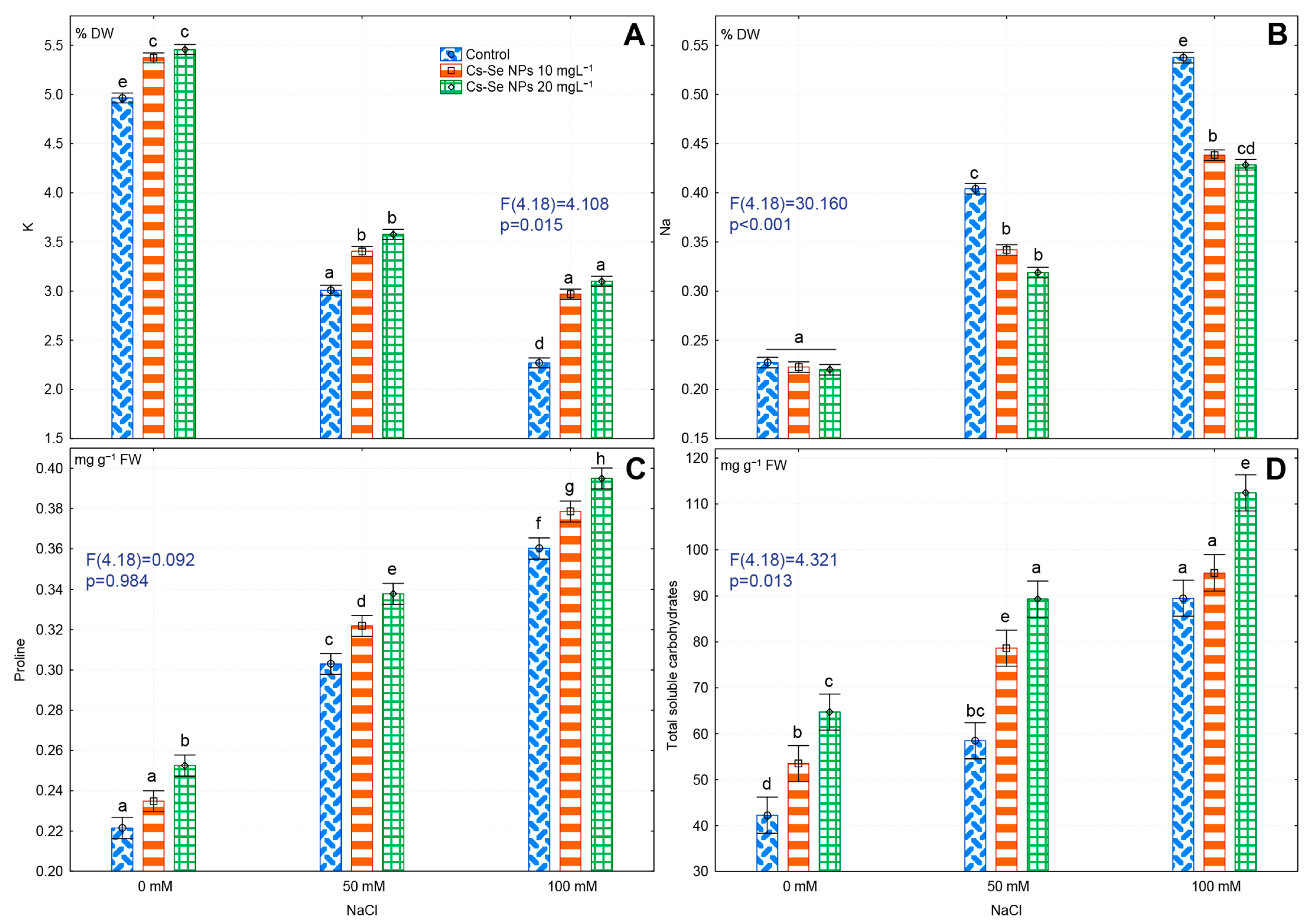
3.4. Effects of Cs-Se NPs on Leaf K and Na Concentration under Saline Conditions
3.5. Effects of Cs–Se NPs on Proline and Soluble Sugars in Leaves under Salinity Stress
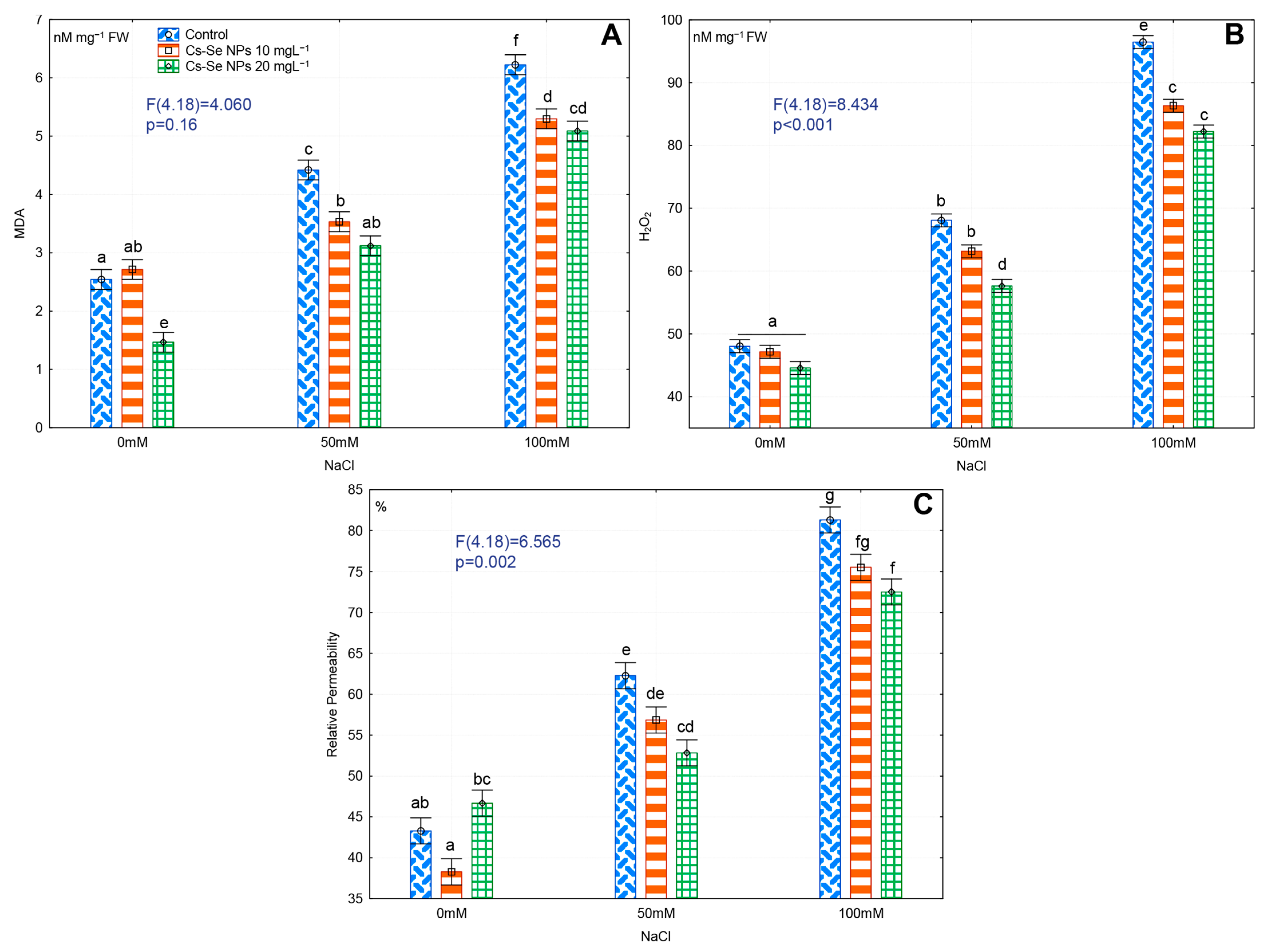
3.6. Effects of Cs–Se NPs on Leaf MDA and H2O2 Content and EL under Salinity Stress
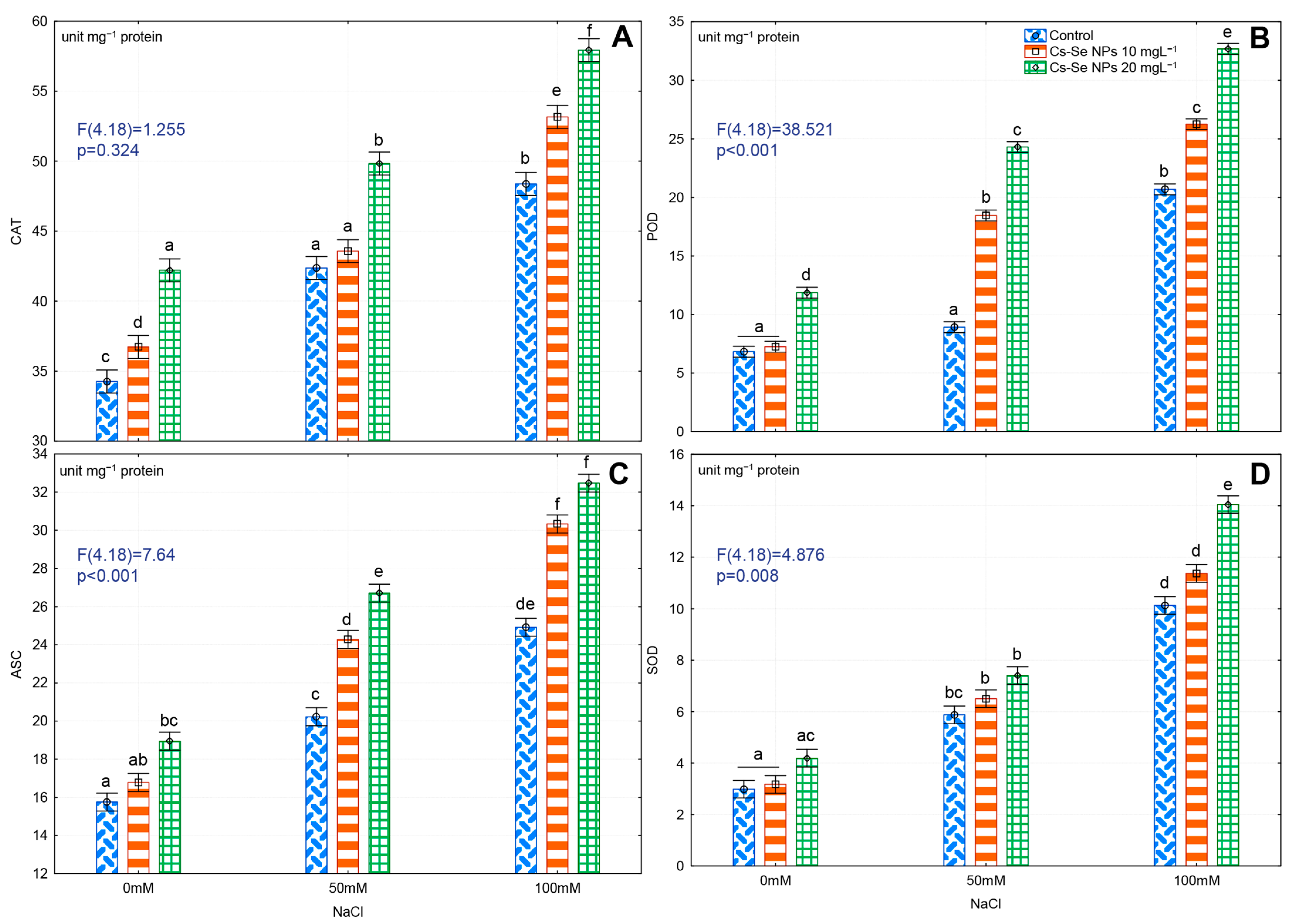
3.7. Effects of Cs–Se NPs on Antioxidant Enzyme Activity in Leaves under Salinity Stress
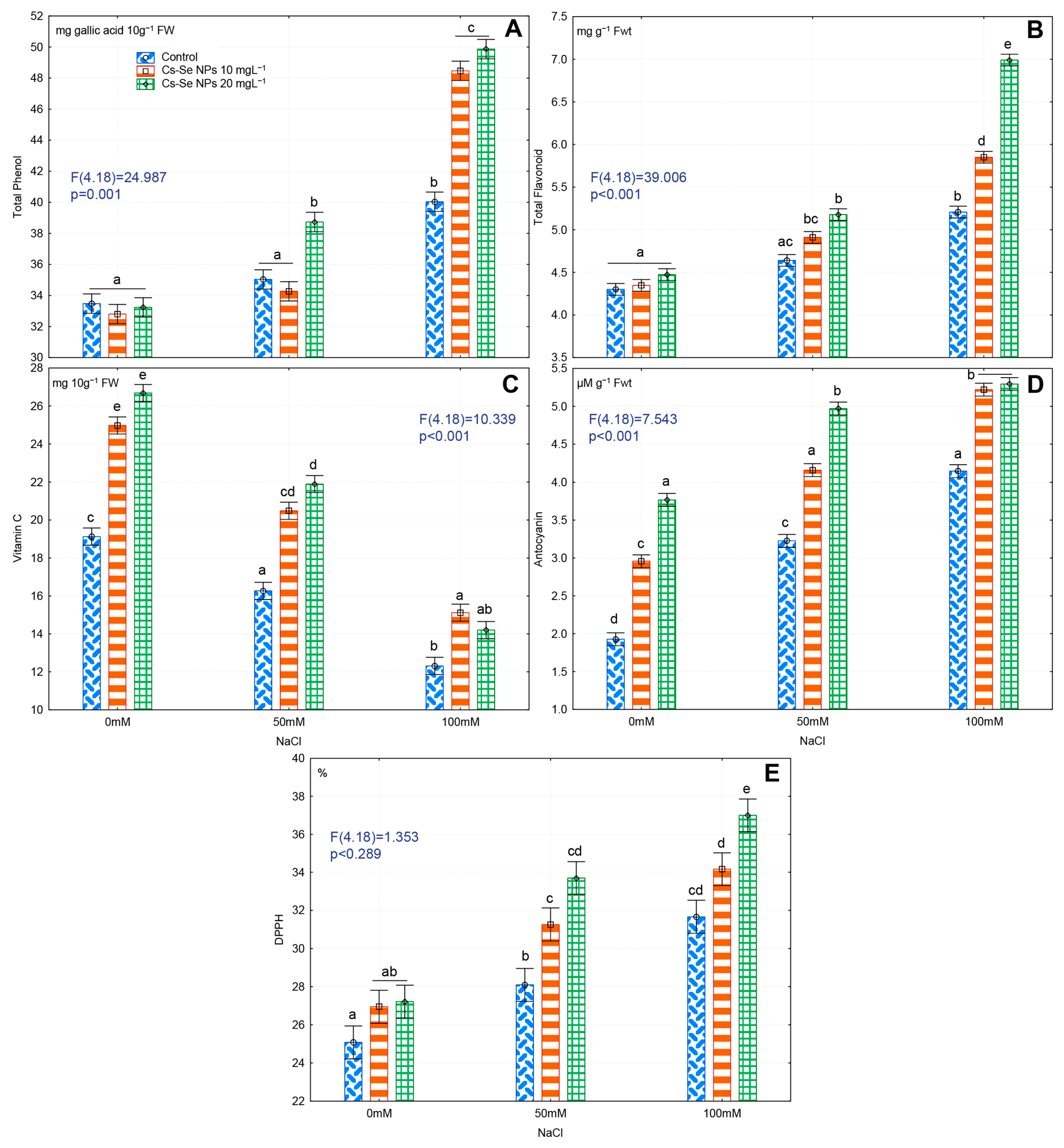
3.8. Effects of Cs-Se NPs on Non-Enzymatic Antioxidant Compounds in Fruit under Salinity Stress
3.9. Effects of Cs-Se NPs on Essential Oil Yield and Content in Fruit from Plants under Salt Stress
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Abdelrahman, M.; Jogaiah, S.; Burritt, D.J.; Tran, L.-S.P. Legume genetic resources and transcriptome dynamics under abiotic stress conditions. Plant Cell Environ. 2018, 41, 1972–1983. [Google Scholar] [CrossRef]
- Babitha, K.; Vemanna, R.S.; Nataraja, K.N.; Udayakumar, M. Overexpression of EcbHLH57 transcription factor from Eleusine coracana L. in tobacco confers tolerance to salt, oxidative and drought stress. PLoS ONE 2015, 10, e0137098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P. Application of nanomaterials toward development of nanobiosensors and their utility in agriculture. In Nanotechnology; Prasad, R., Kumar, M., Kumar, V., Eds.; Springer Nature: Singapore, 2017; pp. 293–303. [Google Scholar] [CrossRef]
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Iqbal, M.; Hussain, I.; Liaqat, H.; Ashraf, M.A.; Rasheed, R.; Rehman, A.U. Exogenously applied selenium reduces oxidative stress and induces heat tolerance in spring wheat. Plant Physiol. Biochem. 2015, 94, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Moore, L.; Mahmoudkhani, A. Methods for removing selenium from aqueous systems. In Proceedings of the Tailings and Mine Waste, Vancouver, BC, Canada, 6–9 November 2011; pp. 6–9. [Google Scholar]
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 1957, 79, 3292–3293. [Google Scholar] [CrossRef]
- Fordyce, F.M. Selenium deficiency and toxicity in the environment. In Essentials of Medical Geology; Springer: Dordrecht, The Netherlands, 2013; pp. 375–416. [Google Scholar]
- Kong, L.; Wang, M.; Bi, D. Selenium modulates the activities of antioxidant enzymes, osmotic homeostasis and promotes the growth of sorrel seedlings under salt stress. Plant Growth Regul. 2005, 45, 155–163. [Google Scholar] [CrossRef]
- Diao, M.; Ma, L.; Wang, J.; Cui, J.; Fu, A.; Liu, H.-Y. Selenium promotes the growth and photosynthesis of tomato seedlings under salt stress by enhancing chloroplast antioxidant defense system. J. Plant Growth Regul. 2014, 33, 671–682. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Beneficial effects of exogenous selenium in cucumber seedlings subjected to salt stress. Biol. Trace Elem. Res. 2009, 132, 259–269. [Google Scholar] [CrossRef]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Allah, E.F.A.; Alqarawi, A.A.; Ahmad, P. Selenium modulates dynamics of antioxidative defence expression, photosynthetic attributes and secondary metabolites to mitigate chromium toxicity in Brassica juncea L. plants. Environ. Exp. Bot. 2019, 161, 180–192. [Google Scholar] [CrossRef]
- Duhan, J.S.; Kumar, R.; Kumar, N.; Kaur, P.; Nehra, K.; Duhan, S. Nanotechnology: The new perspective in precision agriculture. Biotechnol. Rep. 2017, 15, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Gohari, G.; Mohammadi, A.; Akbari, A.; Panahirad, S.; Dadpour, M.R.; Fotopoulos, V.; Kimura, S. Titanium dioxide nanoparticles (TiO2 NPs) promote growth and ameliorate salinity stress effects on essential oil profile and biochemical attributes of Dracocephalum moldavica. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Hussein, H.-A.A.; Darwesh, O.M.; Mekki, B. Environmentally friendly nano-selenium to improve antioxidant system and growth of groundnut cultivars under sandy soil conditions. Biocatal. Agric. Biotechnol. 2019, 18, 101080. [Google Scholar] [CrossRef]
- Morales-Espinoza, M.C.; Cadenas-Pliego, G.; Pérez-Alvarez, M.; Hernández-Fuentes, A.D.; de la Fuente, M.C.; Benavides-Mendoza, A.; Valdés-Reyna, J.; Juárez-Maldonado, A. Se nanoparticles induce changes in the growth, antioxidant responses, and fruit quality of tomato developed under NaCl Stress. Molecules 2019, 24, 3030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djanaguiraman, M.; Belliraj, N.; Bossmann, S.H.; Prasad, P.V. High-temperature stress alleviation by selenium nanoparticle treatment in grain sorghum. ACS Omega 2018, 3, 2479–2491. [Google Scholar] [CrossRef] [Green Version]
- Domokos-Szabolcsy, E.; Marton, L.; Sztrik, A.; Babka, B.; Prokisch, J.; Fari, M. Accumulation of red elemental selenium nanoparticles and their biological effects in Nicotinia tabacum. Plant Growth Regul. 2012, 68, 525–531. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Recent applications of chitin and chitosan-based polymers in plants. Polymers 2019, 11, 839. [Google Scholar] [CrossRef] [Green Version]
- Mujtaba, M.; Khawar, K.M.; Camara, M.C.; Carvalho, L.B.; Fraceto, L.F.; Morsi, R.E.; Elsabee, M.Z.; Kaya, M.; Labidi, J.; Ullah, H.; et al. Chitosan-based delivery systems for plants: A brief overview of recent advances and future directions. Int. J. Biol. Macromol. 2020, 154, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Basch, E.; Gabardi, S.; Ulbricht, C. Bitter melon (Momordica charantia): A review of efficacy and safety. Am. J. Health-Syst. Pharm. 2003, 60, 356–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soomro, A.; Soomro, K.B.; Akhtar, J.; Soomro, S.; Ali, A. Impact of salinity on growth, yield and water use efficiency of Momordica charantia L. under raisedbed irrigation. Int. J. Biol. Biotechnol. 2015, 12, 485–491. [Google Scholar]
- Wang, X.; Ou-yang, C.; Fan, Z.R.; Gao, S.; Chen, F.; Tang, L. Effects of exogenous silicon on seed germination and antioxidant enzyme activities of Momordica charantia under salt stress. J. Anim. Plant Sci. 2010, 6, 700–708. [Google Scholar]
- Alisofi, S.; Einali, A.; Sangtarash, M.H. Jasmonic acid-induced metabolic responses in bitter melon (Momordica charantia) seedlings under salt stress. J. Hortic. Sci. Biotechnol. 2020, 95, 247–259. [Google Scholar] [CrossRef]
- Jafari, H.; Hassanpour, M.; Akbari, A.; Rezaie, J.; Gohari, G.; Mahdavinia, G.R.; Jabbari, E. Characterization of pH-sensitive chitosan/hydroxypropyl methylcellulose composite nanoparticles for delivery of melatonin in cancer therapy. Mater. Lett. 2020, 282, 128818. [Google Scholar] [CrossRef]
- Sairam, R.; Srivastava, G. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci. 2002, 162, 897–904. [Google Scholar] [CrossRef]
- Arnon, A. Method of extraction of chlorophyll in the plants. Agron. J. 1967, 23, 112–121. [Google Scholar]
- Silva, E.N.; Ferreira-Silva, S.L.; de Vasconcelos Fontenele, A.; Ribeiro, R.V.; Viégas, R.A.; Silveira, J.A.G. Photosynthetic changes and protective mechanisms against oxidative damage subjected to isolated and combined drought and heat stresses in Jatropha curcas plants. J. Plant Physiol. 2010, 167, 1157–1164. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Yemm, E.; Willis, A. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Allen, N. Quantitative determination of small amounts of hydrogen peroxide and of ozone. Ind. Eng. Chem. Anal. Ed. 1930, 2, 55–56. [Google Scholar] [CrossRef]
- Rao, G.N. New diet (NTP-2000) for rats in the National Toxicology Program toxicity and carcinogenicity studies. Fundam. Appl. Toxicol. 1996, 32, 102–108. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. Methods Enzym. Anal. 1983, 2, 673–686. [Google Scholar]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Chun, O.K.; Kim, D.-O.; Lee, C.Y. Superoxide radical scavenging activity of the major polyphenols in fresh plums. J. Agric. Food Chem. 2003, 51, 8067–8072. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Suntornsuk, L.; Gritsanapun, W.; Nilkamhank, S.; Paochom, A. Quantitation of vitamin C content in herbal juice using direct titration. J. Pharm. Biomed. Anal. 2002, 28, 849–855. [Google Scholar] [CrossRef]
- Sakamoto, M.; Suzuki, T. Synergistic effects of a night temperature shift and methyl jasmonate on the production of anthocyanin in red leaf lettuce. Am. J. Plant Sci. 2017, 8, 1534. [Google Scholar] [CrossRef] [Green Version]
- Suja, K.; Jayalekshmy, A.; Arumughan, C. Antioxidant activity of sesame cake extract. Food Chem. 2005, 91, 213–219. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, A.; Giri, D.D.; Bhuvaneshwari, K.; Pandey, K.D. Gas chromatography-mass spectrometry analysis and phytochemical screening of methanolic fruit extract of Momordica charantia. J. Recent Adv. Agric. 2012, 1, 122–127. [Google Scholar]
- Wang, Y.H.; Avula, B.; Liu, Y.; Khan, I.A. Determination and quantitation of five cucurbitane triterpenoids in Momordica charantia by reversed-phase high-performance liquid chromatography with evaporative light scattering detection. J. Chromatogr. Sci. 2008, 46, 133–136. [Google Scholar] [CrossRef]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge university press: Cambridge, UK, 2014. [Google Scholar]
- Sarimov, R.M.; Lednev, V.N.; Sibirev, A.V.; Gudkov, S.V. The Use of Fluorescence Spectra for the Detection of Scab and Rot in Fruit and Vegetable Crops. Front. Phys. 2021, 8, 640887. [Google Scholar] [CrossRef]
- Luo, Y.; Zhang, B.; Cheng, W.-H.; Wang, Q. Preparation, characterization and evaluation of selenite-loaded chitosan/TPP nanoparticles with or without zein coating. Carbohydr. Polym. 2010, 82, 942–951. [Google Scholar] [CrossRef]
- Bai, K.; Hong, B.; He, J.; Hong, Z.; Tan, R. Preparation and antioxidant properties of selenium nanoparticles-loaded chitosan microspheres. Int. J. Nanomed. 2017, 12, 4527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef] [Green Version]
- Mozafariyan, M.; Kamelmanesh, M.M.; Hawrylak-Nowak, B. Ameliorative effect of selenium on tomato plants grown under salinity stress. Arch. Agron. Soil Sci. 2016, 62, 1368–1380. [Google Scholar] [CrossRef]
- Hernández-Hernández, H.; Juárez-Maldonado, A.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Sánchez-Aspeytia, D.; González-Morales, S. Chitosan-PVA and copper nanoparticles improve growth and overexpress the SOD and JA genes in tomato plants under salt stress. Agronomy 2018, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.S.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef]
- Abul-Soud, M.; Abd-Elrahman, S.H. Foliar selenium application to improve the tolerance of eggplant grown under salt stress conditions. Int. J. Plant Soil Sci. 2016, 9, 1–10. [Google Scholar] [CrossRef]
- Astaneh, R.K.; Bolandnazar, S.; Nahandi, F.Z.; Oustan, S. The effects of selenium on some physiological traits and K, Na concentration of garlic (Allium sativum L.) under NaCl stress. Inf. Process. Agric. 2018, 5, 156–161. [Google Scholar] [CrossRef]
- Saha, J.; Brauer, E.K.; Sengupta, A.; Popescu, S.C.; Gupta, K.; Gupta, B. Polyamines as redox homeostasis regulators during salt stress in plants. Front. Environ. Sci. 2015, 3, 21. [Google Scholar] [CrossRef] [Green Version]
- Cannella, D.; Möllers, K.B.; Frigaard, N.-U.; Jensen, P.E.; Bjerrum, M.J.; Johansen, K.S.; Felby, C. Light-driven oxidation of polysaccharides by photosynthetic pigments and a metalloenzyme. Nat. Commun. 2016, 7, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N. NaCl-inhibited chlorophyll synthesis and associated changes in ethylene evolution and antioxidative enzyme activities in wheat. Biol. Plant 2003, 47, 437–440. [Google Scholar] [CrossRef]
- Sultana, N.; Ikeda, T.; Itoh, R. Effect of NaCl salinity on photosynthesis and dry matter accumulation in developing rice grains. Environ. Exp. Bot. 1999, 42, 211–220. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Sankar, B.; Sridharan, R.; Panneerselvam, R. Soil salinity alters growth, chlorophyll content, and secondary metabolite accumulation in Cathar roseus. Turk. J. Biol. 2008, 32, 79–83. [Google Scholar]
- Germ, M.; Kreft, I.; Osvald, J. Influence of UV-B exclusion and selenium treatment on photochemical efficiency of photosystem II, yield and respiratory potential in pumpkins (Cucurbita pepo L.). Plant Physiol. Biochem. 2005, 43, 445–448. [Google Scholar] [CrossRef]
- Padmaja, K.; Prasad, D.; Prasad, A. Selenium as a novel regulator of porphyrin biosynthesis in germinating seedlings of mung bean (Phaseolus vulgaris). Biochem. Int. 1990, 22, 441–446. [Google Scholar]
- Saqib, M.; Akhtar, J.; Qureshi, R.H. Pot study on wheat growth in saline and waterlogged compacted soil: I. Grain yield and yield components. Soil Tillage Res. 2004, 77, 169–177. [Google Scholar] [CrossRef]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants, 2nd ed; London Academic Press: London, UK, 2011. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.; Randall, D.; Flowers, T. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell Environ. 2006, 29, 1970–1979. [Google Scholar] [CrossRef]
- Liang, Y.; Chen, Q.; Liu, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef]
- Shekari, F.; Abbasi, A.; Mustafavi, S.H. Effect of silicon and selenium on enzymatic changes and productivity of dill in saline condition. J. Saudi Soc. Agric. Sci. 2017, 16, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Zheng, Q.; Liu, Z.; Xu, W.; Liu, L.; Zhao, G.; Long, X. Overexpression of Arabidopsis thaliana Na+/H+ antiporter gene enhanced salt resistance in transgenic poplar (Populus × euramericana ‘Neva’). Trees 2012, 26, 685–694. [Google Scholar] [CrossRef]
- Subramanyam, K.; Du Laing, G.; Van Damme, E.J. Sodium selenate treatment using a combination of seed priming and foliar spray alleviates salinity stress in rice. Front. Plant Sci. 2019, 10, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, F.; Ashraf, M.; Ahmad, R.; Waraich, E.; Shabbir, R.; Bukhari, M. Supplemental selenium improves wheat grain yield and quality through alterations in biochemical processes under normal and water deficit conditions. Food Chem. 2015, 175, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.N.; Mostofa, M.G.; Akter, M.M.; Srivastava, A.K.; Sayed, M.A.; Hasan, M.S.; Tran, L.-S.P. Impact of salt-induced toxicity on growth and yield-potential of local wheat cultivars: Oxidative stress and ion toxicity are among the major determinants of salt-tolerant capacity. Chemosphere 2017, 187, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, W.R. Salinity and nitrogen nutrition. In Salinity: Environment–Plants–Molecules; Läuchli, A., Lüttge, U., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 229–248. [Google Scholar]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savouré, A. Diversity, distribution and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Fujita, M. Selenium and Plants’ Health: The Physiological Role of Selenium. In Selenium: Sources, Functions and Health Effects; Aomori, C., Hokkaido, M., Eds.; Nova Science Publishers: New York, NY, USA, 2012; pp. 101–122. [Google Scholar]
- Moulick, D.; Santra, S.C.; Ghosh, D. Seed priming with Se mitigates As-induced phytotoxicity in rice seedlings by enhancing essential micronutrient uptake and translocation and reducing as translocation. Environ. Sci. Pollut. Res. 2018, 25, 26978–26991. [Google Scholar] [CrossRef] [PubMed]
- Nouet, C.; Motte, P.; Hanikenne, M. Chloroplastic and mitochondrial metal homeostasis. Trends Plant Sci. 2011, 16, 395–404. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Racchi, M.L. Antioxidant defenses in plants with attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef] [PubMed]
- Ruotolo, R.; Maestri, E.; Pagano, L.; Marmiroli, M.; White, J.C.; Marmiroli, N. Plant response to metal-containing engineered nanomaterials: An omics-based perspective. Environ. Sci. Technol. 2018, 52, 2451–2467. [Google Scholar] [CrossRef]
- Gohari, G.; Safai, F.; Panahirad, S.; Akbari, A.; Rasouli, F.; Dadpour, M.R.; Fotopoulos, V. Modified multiwall carbon nanotubes display either phytotoxic or growth promoting and stress protecting activity in Ocimum basilicum L. in a concentration-dependent manner. Chemosphere 2020, 249, 126171. [Google Scholar] [CrossRef]
- Oloumi, H.; Soltaninejad, R.; Baghizadeh, A. The comparative effects of nano and bulk size particles of CuO and ZnO on glycyrrhizin and phenolic compounds contents in Glycyrrhiza glabra L. seedlings. Indian J. Plant Physiol. 2015, 20, 157–161. [Google Scholar] [CrossRef]
- López-Vargas, E.R.; Ortega-Ortíz, H.; Cadenas-Pliego, G.; de Alba Romenus, K.; de La Fuente, M.C.; Benavides-Mendoza, A.; Juárez-Maldonado, A. Foliar application of copper nanoparticles increases the fruit quality and the content of bioactive compounds in tomatoes. Appl. Sci. 2018, 8, 1020. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xu, H.; Chen, Z.-S.; Chen, G. Biosynthesis of nanoparticles by microorganisms and their applications. J. Nanomater. 2011, 2011, 270974. [Google Scholar] [CrossRef] [Green Version]
- Hochella, M.F.; Lower, S.K.; Maurice, P.A.; Penn, R.L.; Sahai, N.; Sparks, D.L.; Twining, B.S. Nanominerals, mineral nanoparticles, and earth systems. Science 2008, 319, 1631–1635. [Google Scholar] [CrossRef] [Green Version]
- Sosan, A.; Svistunenko, D.; Straltsova, D.; Tsiurkina, K.; Smolich, I.; Lawson, T.; Subramaniam, S.; Golovko, V.; Anderson, D.; Sokolik, A. Engineered silver nanoparticles are sensed at the plasma membrane and dramatically modify the physiology of Arabidopsis thaliana plants. Plant J. 2016, 85, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Mirzajani, F.; Askari, H.; Hamzelou, S.; Schober, Y.; Römpp, A.; Ghassempour, A.; Spengler, B. Proteomics study of silver nanoparticles toxicity on Oryza sativa L. Ecotoxicol. Environ. Saf. 2014, 108, 335–339. [Google Scholar] [CrossRef]
- Kang, J.K.; Kim, O.-H.; Hur, J.; Yu, S.H.; Lamichhane, S.; Lee, J.W.; Ojha, U.; Hong, J.H.; Lee, C.S.; Cha, J.-Y. Increased intracellular Ca2+ concentrations prevent membrane localization of PH domains through the formation of Ca2+-phosphoinositides. PNAS 2017, 114, 11926–11931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY transcription factors: Molecular regulation and stress responses in plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kole, C.; Kole, P.; Randunu, K.M.; Choudhary, P.; Podila, R.; Ke, P.C.; Rao, A.M.; Marcus, R.K. Nanobiotechnology can boost crop production and quality: First evidence from increased plant biomass, fruit yield and phytomedicine content in bitter melon (Momordica charantia). BMC Biotechnol. 2013, 13, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl (mM) | Treatments | Plant Height (cm) | Shoot FW (g) | Shoot DW (g) | Root FW (g) | Root DW (g) | RWC (%) | No. Fruits/Plant | Fruit Weight (kg) | Yield (kg) |
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | Control | 3.97 ± 0.09 a | 240.33 ± 16.31 a | 65.95 ± 4.17 a | 155.69 ± 1.00 a | 27.89 ± 0.17 b | 91.17 ± 2.98 b | 21 ± 0.81 b | 0.071 ± 0.003 a | 1.501 ± 0.016 b |
| Cs–Se NPs 10 mg L−1 | 3.99 ± 0.04 a | 242.73 ± 6.03 a | 67.25 ± 3.24 a | 156.54 ± 1.66 a | 28.06 ± 0.28 ab | 92.56 ± 1.71 ab | 23 ± 0.81 a | 0.06 ± 0.002 b | 1.470 ± 0.020 b | |
| Cs–Se NPs 20 mg L−1 | 4.07 ± 0.06 a | 246.84 ± 15.02 a | 69.60 ± 2.18 a | 158.32 ± 1.41 a | 28.81 ± 0.51 a | 95.98 ± 1.47 a | 22.66 ± 0.47 a | 0.075 ± 0.002 a | 1.702 ± 0.015 a | |
| 50 | Control | 2.51 ± 0.11 c | 143.77 ± 9.66 bc | 48.48 ± 2.71 bc | 107.26 ± 1.78 c | 19.25 ± 0.47 d | 72.26 ± 1.61 de | 14.33 ± 0.47 d | 0.052 ± 0.001 c | 0.747 ± 0.010 e |
| Cs–Se NPs 10 mg L−1 | 2.67 ± 0.04 b | 166.11 ± 6.70 b | 49.27 ± 0.73 bc | 123.48 ± 3.76 b | 22.16 ± 0.48 c | 76.56 ± 0.93 d | 15.66 ± 0.94 cd | 0.053 ± 0.006 c | 0.827 ± 0.053 d | |
| Cs–Se NPs 20 mg L−1 | 2.70 ± 0.05 b | 173.17 ± 11.26 b | 52.21 ± 0.87 b | 124.14 ± 1.15 b | 22.28 ± 0.34 c | 83.50 ± 3.24 c | 16.33 ± 0.47 c | 0.061 ± 0.001 b | 0.996 ± 0.004 c | |
| 100 | Control | 1.22 ± 0.05 e | 89.37 ± 5.43 e | 38.72 ± 0.65 e | 69.01 ± 2.51 e | 12.39 ± 0.42 f | 53.81 ± 2.17 f | 9 ± 0.81 f | 0.035 ± 0.002 e | 0.316 ± 0.026 g |
| Cs–Se NPs 10 mg L−1 | 1.34 ± 0.04 de | 108.23 ± 4.52 de | 40.71 ± 0.83 de | 73.92 ± 4.95 de | 13.27 ± 0.77 e | 70.70 ± 2.69 e | 11 ± 1.41 e | 0.032 ± 0.003 e | 0.352 ± 0.014 g | |
| Cs–Se NPs 20 mg L−1 | 1.41 ± 0.05 d | 117.94 ± 2.08 d | 44.69 ± 1.09 cd | 76.93 ± 0.71 d | 13.83 ± 0.11 e | 74.26 ± 0.84 de | 10 ± 1.41 ef | 0.043 ± 0.006 d | 0.422 ± 0.010 f |
| NaCl (mM) | Treatments | Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Total Chl (mg g−1 FW) | Carotenoids (mg g−1 FW) | Pn (µmol m−2 s−1) |
|---|---|---|---|---|---|---|
| 0 | Control | 11.40 ± 0.45 b | 4.39 ± 0.11 b | 15.80 ± 0.45 b | 0.780 ± 0.01 b | 9.60 ± 0.23 bc |
| Cs–Se NPs 10 mg L−1 | 11.98 ± 0.46 ab | 4.44 ± 0.08 b | 16.42 ± 0.43 ab | 0.820 ± 0.03 ab | 9.93 ± 0.14 b | |
| Cs–Se NPs 20 mg L−1 | 12.65 ± 0.70 a | 4.79 ± 0.14 a | 17.44 ± 0.57 a | 0.835 ± 0.03 a | 10.67 ± 0.25 a | |
| 50 | Control | 8.19 ± 0.82 de | 3.03 ± 0.05 d | 11.22 ± 0.87 e | 0.565 ± 0.00 d | 8.50 ± 0.16 ef |
| Cs–Se NPs 10 mg L−1 | 8.93 ± 0.65 d | 3.22 ± 0.15 d | 12.40 ± 0.58 d | 0.591 ± 0.01 d | 8.85 ± 0.08 de | |
| Cs–Se NPs 20 mg L−1 | 10.27 ± 0.59 c | 3.47 ± 0.09 c | 13.49 ± 0.44 c | 0.669 ± 0.02 c | 9.22 ± 0.12 cd | |
| 100 | Control | 5.27 ± 1.19 g | 1.89 ± 0.06 g | 7.17 ± 1.17 g | 0.430 ± 0.09 f | 7.26 ± 0.14 h |
| Cs–Se NPs 10 mg L−1 | 6.59 ± 0.66 f | 2.18 ± 0.12 f | 8.78 ± 0.72 f | 0.478 ± 0.00 e | 7.94 ± 0.20 g | |
| Cs–Se NPs 20 mg L−1 | 7.68 ± 0.54 e | 2.58 ± 0.21 e | 10.26 ± 0.70 e | 0.517 ± 0.00 e | 8.17 ± 0.14 g |
| NaCl (mM) | Treatments | Oil content (%) | Charantin | Momordin | Momordicoside | Momordicin | Karaviloside | Cucuritan | Oleic acid | Stigmasterol | Cis-9-hexadecenal | Cucurbitacin | pentadecyne | Gentisic acid |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | Control | 0.380 ± 0.02 f | 1.40 ± 0.07 c | 0.643 ± 0.19 e | 0.341 ± 0.00 d | 0.498 ± 0.01 c | 0.787 ± 0.08 bc | 0.495 ± 0.01 bcd | 0.700 ± 0.07 cd | 1.07 ± 0.08 f | 2.47 ± 0.06 c | 1.42 ± 0.32 bcd | 9.76 ± 1.47 a | 6.35 ± 0.34 c |
| Cs–Se NPs 10 mg L−1 | 0.480 ± 0.02 f | 1.84 ± 0.10 b | 0.623 ± 0.14 e | 0.348 ± 0.01 d | 0.844 ± 0.00 a | 0.667 ± 0.01 c | 0.484 ± 0.00 bcde | 1.316 ± 0.13 a | 1.41 ± 0.09 de | 2.36 ± 0.06 c | 2.30 ± 0.57 a | 8.51 ± 1.37 ab | 7.43 ± 0.20 b | |
| Cs–Se NPs 20 mg L−1 | 0.833 ± 0.01 e | 2.85 ± 0.10 a | 0.566 ± 0.19 e | 0.348 ± 0.01 d | 0.816 ± 0.00 a | 0.682 ± 0.01 c | 0.532 ± 0.00 bc | 1.323 ± 0.10 a | 1.20 ± 0.08 ef | 3.18 ± 0.07 a | 1.96 ± 0.10 ab | 5.66 ± 1.41 b | 9.34 ± 0.24 a | |
| 50 | Control | 1.02 ± 0.04 d | 1.12 ± 0.08 de | 1.10 ± 0.03 cd | 0.487 ± 0.00 b | 0.314 ± 0.01 d | 0.877 ± 0.07 b | 0.549 ± 0.08 b | 0.740 ± 0.09 bcd | 1.21 ± 0.12 ef | 1.91 ± 0.09 e | 0.91 ± 0.17 cd | 10.72 ± 1.50 a | 5.71 ± 0.26 de |
| Cs–Se NPs 10 mg L−1 | 1.24 ± 0.07 abc | 1.55 ± 0.09 c | 1.51 ± 0.08 b | 0.501 ± 0.01 b | 0.786 ± 0.00 a | 0.786 ± 0.01 bc | 0.474 ± 0.00 cde | 0.916 ± 0.05 b | 1.65 ± 0.21 d | 2.27 ± 0.11 cd | 1.56 ± 0.23 b | 9.84 ± 1.32 a | 6.45 ± 0.30 c | |
| Cs–Se NPs 20 mg L−1 | 1.17 ± 0.05 bc | 1.92 ± 0.06 b | 1.81 ± 0.02 a | 0.583 ± 0.02 a | 0.695 ± 0.00 b | 1.94 ± 0.05 a | 0.727 ± 0.00 a | 0.906 ± 0.08 b | 1.97 ± 0.08 c | 2.06 ± 0.07 de | 1.66 ± 0.14 b | 8.21 ± 1.50 ab | 7.24 ± 0.23 b | |
| 100 | Control | 1.15 ± 0.04 c | 0.966 ± 0.06 ef | 0.856 ± 0.09 de | 0.211 ± 0.02 e | 0.163 ± 0.01 e | 0.770 ± 0.05 bc | 0.392 ± 0.00 f | 0.663 ± 0.08 d | 2.29 ± 0.22 b | 2.39 ± 0.11 c | 0.862 ± 0.11 d | 11.74 ± 1.51 a | 5.21 ± 0.26 e |
| Cs–Se NPs 10 mg L−1 | 1.36 ± 0.04 a | 0.916 ± 0.04 f | 1.17 ± 0.04 c | 0.430 ± 0.01 c | 0.298 ± 0.01 d | 0.773 ± 0.06 bc | 0.417 ± 0.01 ef | 0.870 ± 0.07 bc | 2.68 ± 0.05 a | 1.89 ± 0.04 e | 1.50 ± 0.25 bc | 11.66 ± 1.33 a | 5.17 ± 0.31 e | |
| Cs–Se NPs 20 mg L−1 | 1.29 ± 0.08 ab | 1.20 ± 0.06 d | 1.35 ± 0.12 bc | 0.518 ± 0.01 b | 0.347 ± 0.11 d | 0.846 ± 0.08 b | 0.449 ± 0.03 def | 0.860 ± 0.04bc | 2.87 ± 0.08 a | 2.76 ± 0.19 b | 0.868 ± 0.06 d | 11.04 ± 1.53 a | 5.83 ± 0.21 cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheikhalipour, M.; Esmaielpour, B.; Behnamian, M.; Gohari, G.; Giglou, M.T.; Vachova, P.; Rastogi, A.; Brestic, M.; Skalicky, M. Chitosan–Selenium Nanoparticle (Cs–Se NP) Foliar Spray Alleviates Salt Stress in Bitter Melon. Nanomaterials 2021, 11, 684. https://doi.org/10.3390/nano11030684
Sheikhalipour M, Esmaielpour B, Behnamian M, Gohari G, Giglou MT, Vachova P, Rastogi A, Brestic M, Skalicky M. Chitosan–Selenium Nanoparticle (Cs–Se NP) Foliar Spray Alleviates Salt Stress in Bitter Melon. Nanomaterials. 2021; 11(3):684. https://doi.org/10.3390/nano11030684
Chicago/Turabian StyleSheikhalipour, Morteza, Behrooz Esmaielpour, Mahdi Behnamian, Gholamreza Gohari, Mousa Torabi Giglou, Pavla Vachova, Anshu Rastogi, Marian Brestic, and Milan Skalicky. 2021. "Chitosan–Selenium Nanoparticle (Cs–Se NP) Foliar Spray Alleviates Salt Stress in Bitter Melon" Nanomaterials 11, no. 3: 684. https://doi.org/10.3390/nano11030684
APA StyleSheikhalipour, M., Esmaielpour, B., Behnamian, M., Gohari, G., Giglou, M. T., Vachova, P., Rastogi, A., Brestic, M., & Skalicky, M. (2021). Chitosan–Selenium Nanoparticle (Cs–Se NP) Foliar Spray Alleviates Salt Stress in Bitter Melon. Nanomaterials, 11(3), 684. https://doi.org/10.3390/nano11030684