Multiplexed smFRET Nucleic Acid Sensing Using DNA Nanotweezers


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. DNA Construct Assembly
2.3. Surface-Functionalization of Flow Cells
2.4. Sample Preparation and Imaging
2.5. Data Acquisition and Analysis
3. Results and Discussion
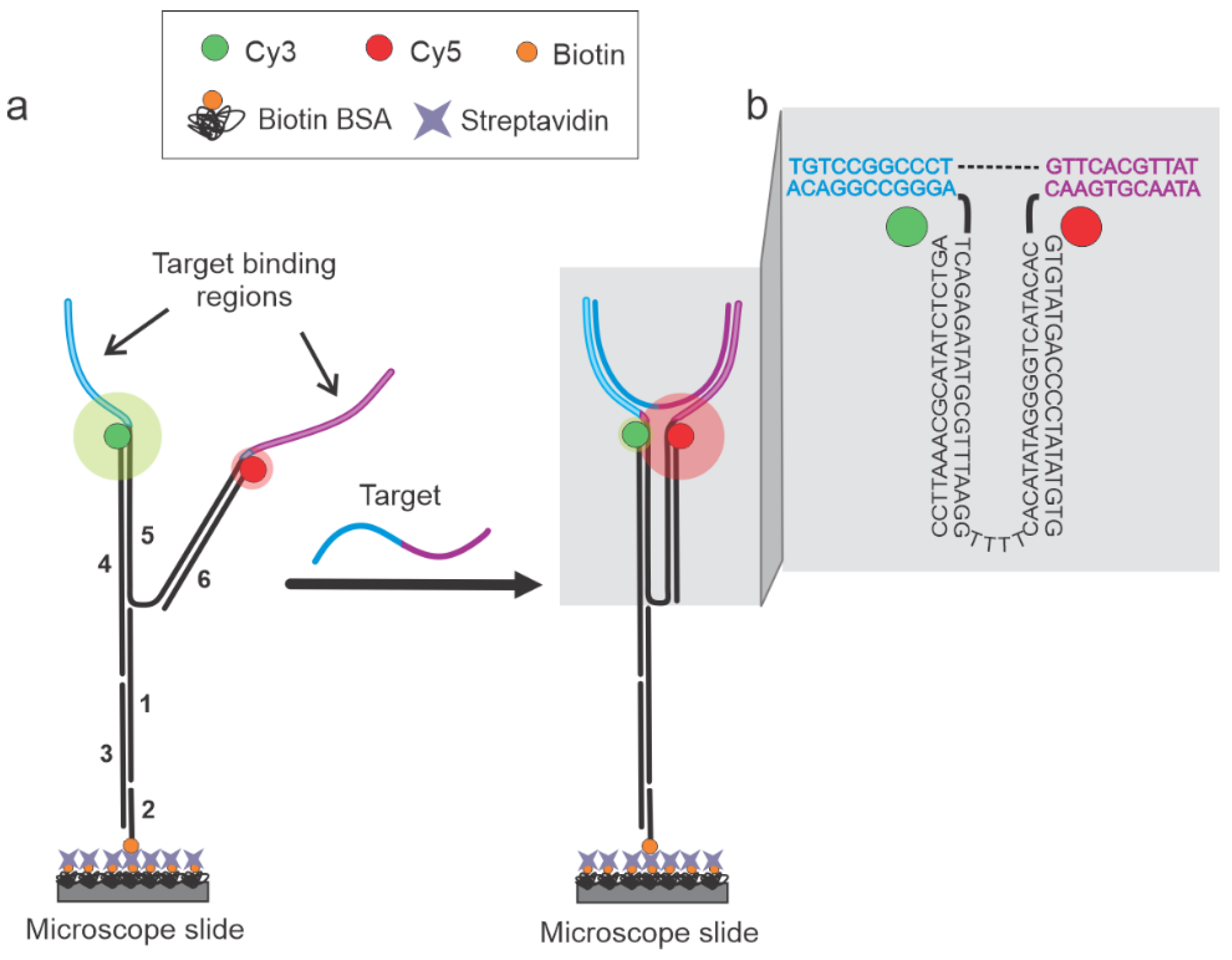
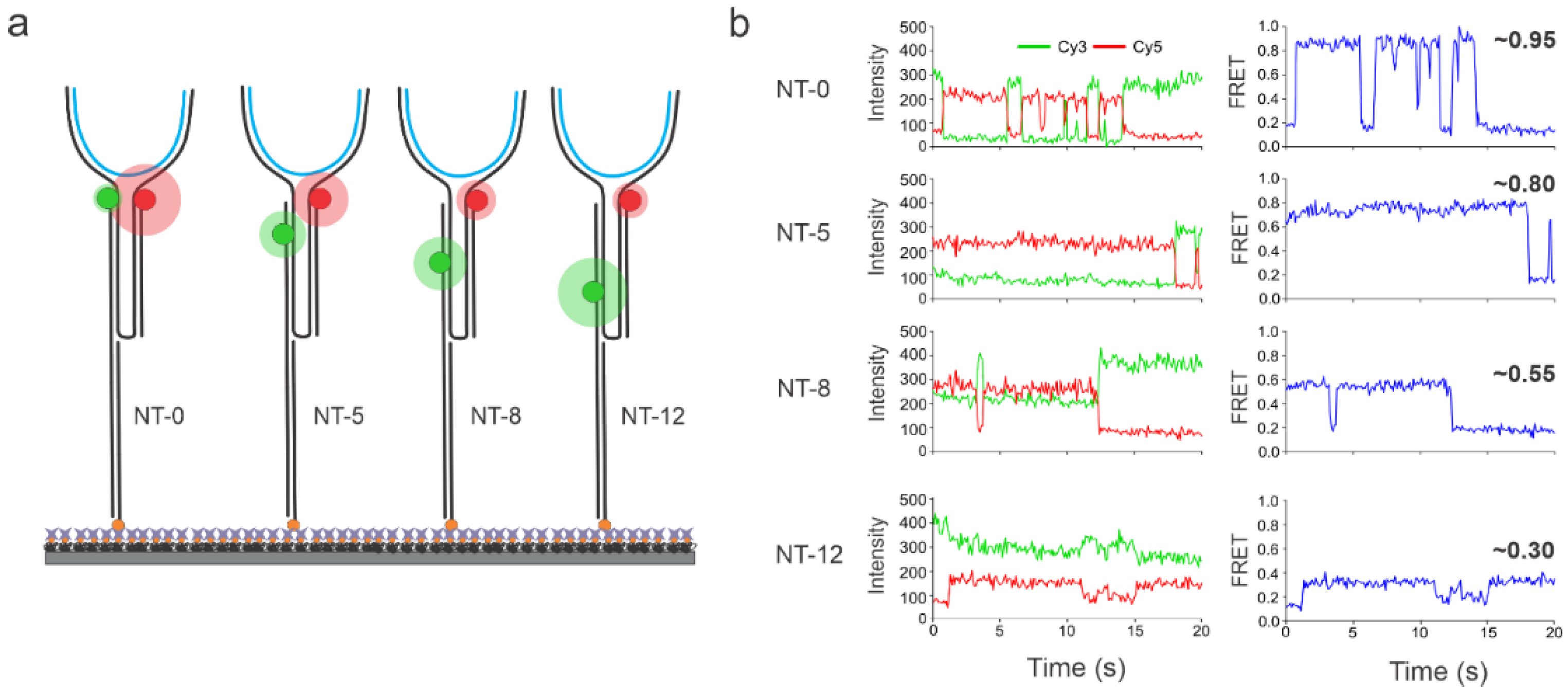
3.1. Sensor Design and Working Principles
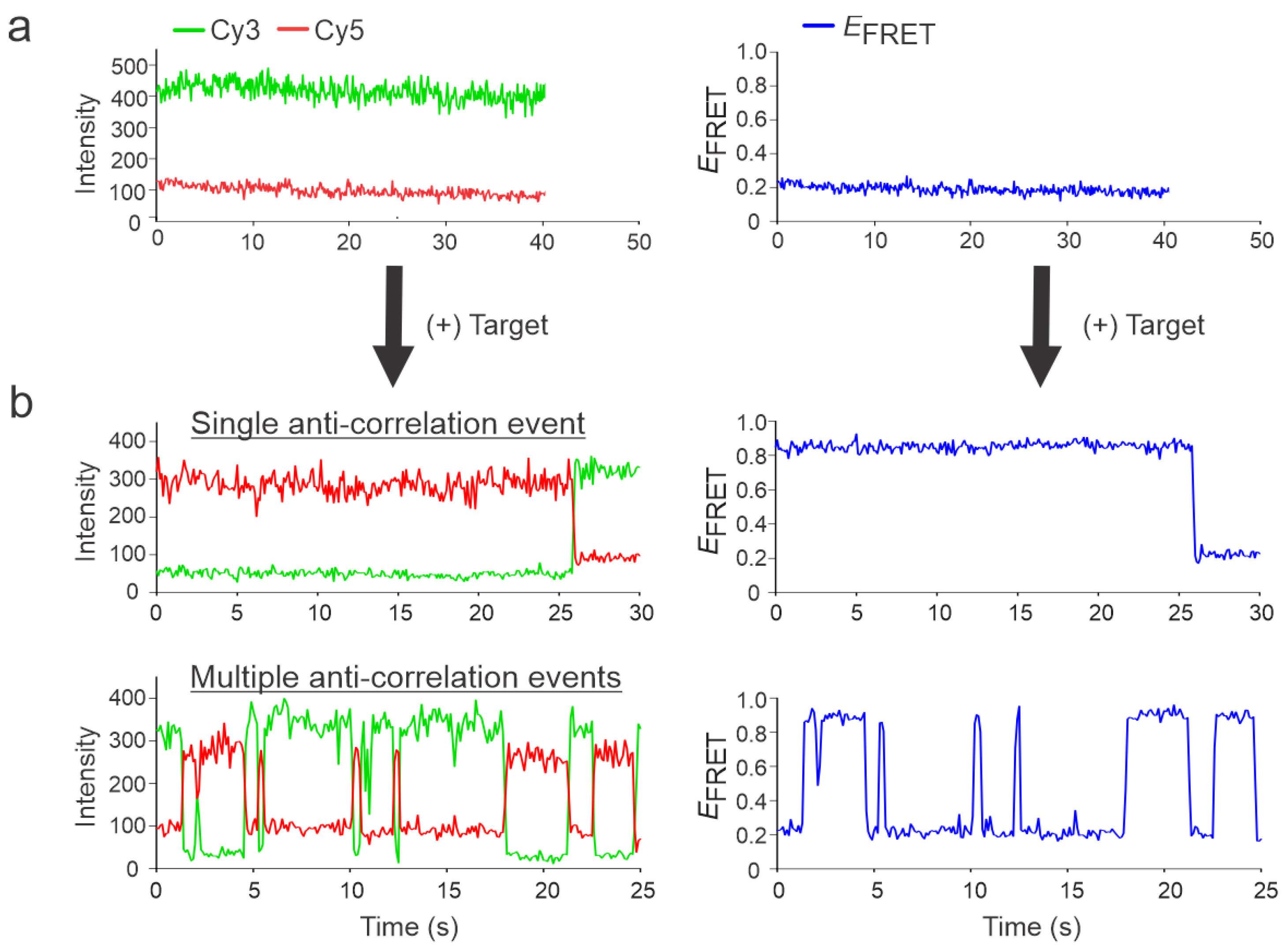
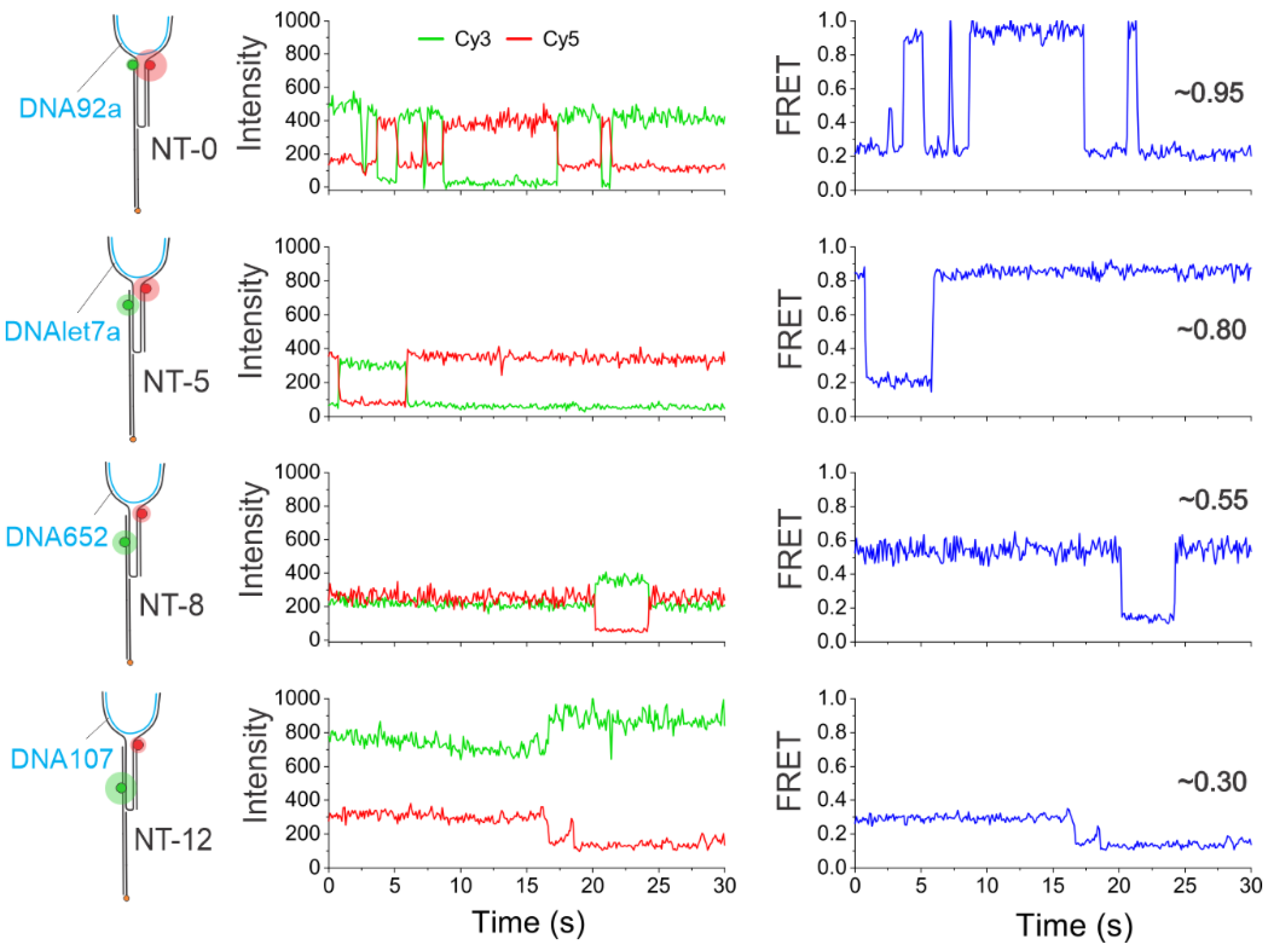
3.2. Single-Molecule Characterization of Sensors
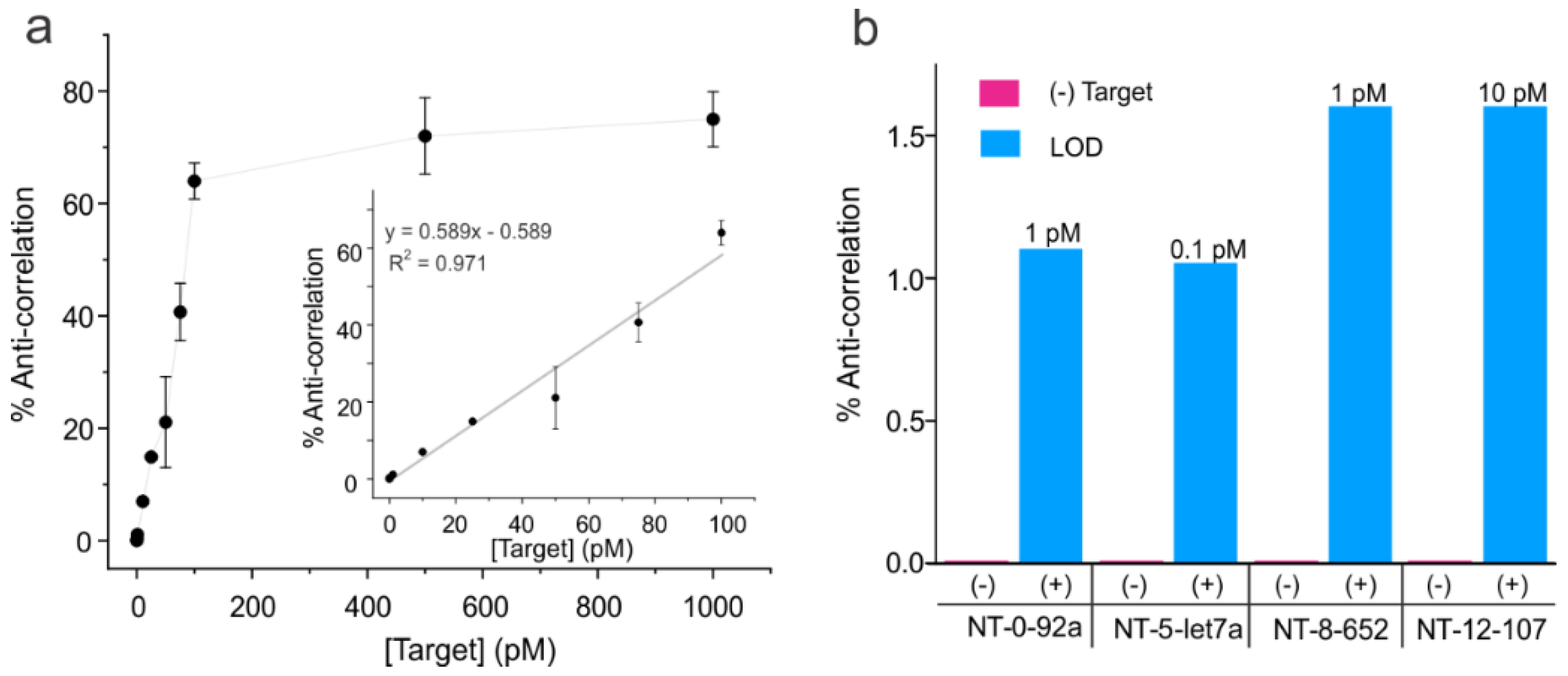
3.3. Determination of Analytical Sensitivity and Specificity of NT Sensors
3.4. Validation of the Multiplexed Sensing Method
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Timilsina, S.S.; Jolly, P.; Durr, N.; Yafia, M.; Ingber, D.E. Enabling Multiplexed Electrochemical Detection of Biomarkers with High Sensitivity in Complex Biological Samples. Acc. Chem. Res. 2021, 54, 3529–3539. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, H.; Yang, X.; Wang, K.; Liu, F.; Zhao, Q.; Liu, P.; Liu, R. Multiplex Detection of Nucleic Acids Using a Low Cost Microfluidic Chip and a Personal Glucose Meter at the Point-of-Care. Chem. Commun. 2014, 50, 3824–3826. [Google Scholar] [CrossRef]
- He, X.-P.; Hu, X.-L.; James, T.D.; Yoon, J.; Tian, H. Multiplexed Photoluminescent Sensors: Towards Improved Disease Diagnostics. Chem. Soc. Rev. 2017, 46, 6687–6696. [Google Scholar] [CrossRef] [Green Version]
- Mo, M.-H.; Chen, L.; Fu, Y.; Wang, W.; Fu, S.W. Cell-Free Circulating MiRNA Biomarkers in Cancer. J. Cancer 2012, 3, 432–448. [Google Scholar] [CrossRef]
- Wei, F.; Patel, P.; Liao, W.; Chaudhry, K.; Zhang, L.; Arellano-Garcia, M.; Hu, S.; Elashoff, D.; Zhou, H.; Shukla, S.; et al. Electrochemical Sensor for Multiplex Biomarkers Detection. Clin. Cancer Res. 2009, 15, 4446–4452. [Google Scholar] [CrossRef] [Green Version]
- Kaur, A.; Sapkota, K.; Dhakal, S. Multiplexed Nucleic Acid Sensing with Single-Molecule FRET. ACS Sens. 2019, 4, 623–633. [Google Scholar] [CrossRef]
- Taketa, K. α-Fetoprotein: Reevaluation in Hepatology. Hepatology 1990, 12, 1420–1432. [Google Scholar] [CrossRef]
- Kodadek, T. Protein Microarrays: Prospects and Problems. Chem. Biol. 2001, 8, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Xu, M.; Tian, X.; Cai, S.; Zeng, S. Research Advances in the Detection of MiRNA. J. Pharm. Anal. 2019, 9, 217–226. [Google Scholar] [CrossRef]
- Cissell, K.A.; Shrestha, S.; Deo, S.K. MicroRNA Detection: Challenges for the Analytical Chemist. Anal. Chem. 2007, 79, 4754–4761. [Google Scholar] [CrossRef]
- Ouyang, T.; Liu, Z.; Han, Z.; Ge, Q. MicroRNA Detection Specificity: Recent Advances and Future Perspective. Anal. Chem. 2019, 91, 3179–3186. [Google Scholar] [CrossRef] [Green Version]
- Gunderson, K.L.; Steemers, F.J.; Lee, G.; Mendoza, L.G.; Chee, M.S. A Genome-Wide Scalable SNP Genotyping Assay Using Microarray Technology. Nat. Genet. 2005, 37, 549–554. [Google Scholar] [CrossRef]
- Li, D.; Xia, L.; Zhou, Q.; Wang, L.; Chen, D.; Gao, X.; Li, Y. Label-Free Detection of MiRNA Using Surface-Enhanced Raman Spectroscopy. Anal. Chem. 2020, 92, 12769–12773. [Google Scholar] [CrossRef]
- Mehta, S.; Zhang, Y.; Roth, R.H.; Zhang, J.; Mo, A.; Tenner, B.; Huganir, R.L.; Zhang, J. Single-Fluorophore Biosensors for Sensitive and Multiplexed Detection of Signalling Activities. Nat. Cell. Biol. 2018, 20, 1215–1225. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Chen, S.X.; Yin, P. Optimizing the Specificity of Nucleic Acid Hybridization. Nat. Chem. 2012, 4, 208–214. [Google Scholar] [CrossRef]
- Geiss, G.K.; Bumgarner, R.E.; Birditt, B.; Dahl, T.; Dowidar, N.; Dunaway, D.L.; Fell, H.P.; Ferree, S.; George, R.D.; Grogan, T.; et al. Direct Multiplexed Measurement of Gene Expression with Color-Coded Probe Pairs. Nat. Biotechnol. 2008, 26, 317–325. [Google Scholar] [CrossRef]
- Kang, D.; White, R.J.; Xia, F.; Zuo, X.; Vallée-Bélisle, A.; Plaxco, K.W. DNA Biomolecular-Electronic Encoder and Decoder Devices Constructed by Multiplex Biosensors. NPG Asia Mater. 2012, 4, e1. [Google Scholar] [CrossRef]
- Bunt, G.; Wouters, F.S. FRET from Single to Multiplexed Signaling Events. Biophys. Rev. 2017, 9, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Tsai, H.-Y.; Algar, W.R. A Dendrimer-Based Time-Gated Concentric FRET Configuration for Multiplexed Sensing. ACS Nano. 2022, 16, 8150–8160. [Google Scholar] [CrossRef]
- Algar, W.R.; Malanoski, A.P.; Susumu, K.; Stewart, M.H.; Hildebrandt, N.; Medintz, I.L. Multiplexed Tracking of Protease Activity Using a Single Color of Quantum Dot Vector and a Time-Gated Förster Resonance Energy Transfer Relay. Anal. Chem. 2012, 84, 10136–10146. [Google Scholar] [CrossRef]
- Guo, J.; Mingoes, C.; Qiu, X.; Hildebrandt, N. Simple, Amplified, and Multiplexed Detection of MicroRNAs Using Time-Gated FRET and Hybridization Chain Reaction. Anal. Chem. 2019, 91, 3101–3109. [Google Scholar] [CrossRef] [Green Version]
- Stein, I.H.; Steinhauer, C.; Tinnefeld, P. Single-Molecule Four-Color FRET Visualizes Energy-Transfer Paths on DNA Origami. J. Am. Chem. Soc. 2011, 133, 4193–4195. [Google Scholar] [CrossRef]
- Yim, S.W.; Kim, T.; Laurence, T.A.; Partono, S.; Kim, D.; Kim, Y.; Weiss, S.; Reitmair, A. Four-Color Alternating-Laser Excitation Single-Molecule Fluorescence Npectroscopy for Next-Generation Biodetection Assays. Clin. Chem. 2012, 58, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Mo, G.C.H.; Posner, C.; Rodriguez, E.A.; Sun, T.; Zhang, J. A Rationally Enhanced Red Fluorescent Protein Expands the Utility of FRET Biosensors. Nat. Commun. 2020, 11, 1848. [Google Scholar] [CrossRef] [Green Version]
- Jungmann, R.; Avendaño, M.S.; Woehrstein, J.B.; Dai, M.; Shih, W.M.; Yin, P. Multiplexed 3D Cellular Super-Resolution Imaging with DNA-PAINT and Exchange-PAINT. Nat. Methods 2014, 11, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yi, X.; Tang, H.; Han, H.; Wu, M.; Zhou, F. Direct Quantification of MicroRNA at Low Picomolar Level in Sera of Glioma Patients Using a Competitive Hybridization Followed by Amplified Voltammetric Detection. Anal. Chem. 2012, 84, 6400–6406. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Shi, X.-M.; Guo, H.-Q.; Zhao, X.-Z.; Zhao, W.-W.; Xu, J.-J.; Chen, H.-Y. Gold Nanoparticle Couples with Entropy-Driven Toehold-Mediated DNA Strand Displacement Reaction on Magnetic Beads: Toward Ultrasensitive Energy-Transfer-Based Photoelectrochemical Detection of MiRNA-141 in Real Blood Sample. Anal. Chem. 2018, 90, 11892–11898. [Google Scholar] [CrossRef]
- Kleivi Sahlberg, K.; Bottai, G.; Naume, B.; Burwinkel, B.; Calin, G.A.; Børresen-Dale, A.-L.; Santarpia, L. A Serum MicroRNA Signature Predicts Tumor Relapse and Survival in Triple-Negative Breast Cancer Patients. Clin. Cancer Res. 2015, 21, 1207–1214. [Google Scholar] [CrossRef] [Green Version]
- Shin, V.Y.; Siu, J.M.; Cheuk, I.; Ng, E.K.O.; Kwong, A. Circulating Cell-Free MiRNAs as Biomarker for Triple-Negative Breast Cancer. Br. J. Cancer 2015, 112, 1751–1759. [Google Scholar] [CrossRef]
- Thakur, S.; Grover, R.K.; Gupta, S.; Yadav, A.K.; Das, B.C. Identification of Specific MiRNA Signature in Paired Sera and Tissue Samples of Indian Women with Triple Negative Breast Cancer. PLoS ONE 2016, 11, e0158946. [Google Scholar] [CrossRef]
- Si, H.; Sun, X.; Chen, Y.; Cao, Y.; Chen, S.; Wang, H.; Hu, C. Circulating MicroRNA-92a and MicroRNA-21 as Novel Minimally Invasive Biomarkers for Primary Breast Cancer. J. Cancer Res. Clin. Oncol. 2013, 139, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, D.R.; Dhakal, S. Single-Molecule Imaging Reveals Conformational Manipulation of Holliday Junction DNA by the Junction Processing Protein RuvA. Biochemistry 2018, 57, 3616–3624. [Google Scholar] [CrossRef]
- Gibbs, D.; Kaur, A.; Megalathan, A.; Sapkota, K.; Dhakal, S. Build Your Own Microscope: Step-by-Step Guide for Building a Prism-Based TIRF Microscope. Methods Protoc. 2018, 1, 40. [Google Scholar] [CrossRef] [Green Version]
- Sapkota, K.; Kaur, A.; Megalathan, A.; Donkoh-Moore, C.; Dhakal, S. Single-Step FRET-Based Detection of Femtomoles DNA. Sensors 2019, 19, 3495. [Google Scholar] [CrossRef] [Green Version]
- Ha, T. Single-Molecule Fluorescence Resonance Energy Transfer. Methods 2001, 25, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Roy, R.; Hohng, S.; Ha, T. A Practical Guide to Single-Molecule FRET. Nat. Methods 2008, 5, 507–516. [Google Scholar] [CrossRef]
- Yang, W.; Shen, Y.; Zhang, D.; Li, C.; Yuan, R.; Xu, W. Programmed Dual-Functional DNA Tweezer for Simultaneous and Recognizable Fluorescence Detection of MicroRNA and Protein. Anal. Chem. 2019, 91, 7782–7789. [Google Scholar] [CrossRef]
- Lu, C.-H.; Willner, B.; Willner, I. DNA Nanotechnology: From Sensing and DNA Machines to Drug-Delivery Systems. ACS Nano 2013, 7, 8320–8332. [Google Scholar] [CrossRef]
- Zeng, S.; Liu, D.; Li, C.; Yu, F.; Fan, L.; Lei, C.; Huang, Y.; Nie, Z.; Yao, S. Cell-Surface-Anchored Ratiometric DNA Tweezer for Real-Time Monitoring of Extracellular and Apoplastic pH. Anal. Chem. 2018, 90, 13459–13466. [Google Scholar] [CrossRef]
- Chen, X.; Fu, X.; Wu, Y.; Jin, Y.; Li, W. A DNA Tweezers-Actuated Regenerated DNAzyme Nanoreactor: A Fluorescence Sensor for MiRNA and Adenosine Detection. Anal. Methods 2020, 12, 1579–1586. [Google Scholar] [CrossRef]
- Xiong, Z.; Wang, Q.; Zhang, J.; Yun, W.; Wang, X.; Ha, X.; Yang, L. A Simple and Programmed DNA Tweezer Probes for One-Step and Amplified Detection of UO22+. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 229, 118017. [Google Scholar] [CrossRef]
- Yurke, B.; Turberfield, A.J.; Mills, A.P.; Simmel, F.C.; Neumann, J.L. A DNA-Fuelled Molecular Machine Made of DNA. Nature 2000, 406, 605–608. [Google Scholar] [CrossRef]
- Li, W.; Wu, Y.; Jin, Y.; Wang, J.; Gao, H. A Regenerated Nanotweezer-Managed Catalytic DNA Circuit for Cancer-Related Sensing. Sens. Actuators B Chem. 2020, 305, 127474. [Google Scholar] [CrossRef]
- Shigeto, H.; Nakatsuka, K.; Ikeda, T.; Hirota, R.; Kuroda, A.; Funabashi, H. Continuous Monitoring of Specific MRNA Expression Responses with a Fluorescence Resonance Energy Transfer-Based DNA Nano-Tweezer Technique That Does Not Require Gene Recombination. Anal. Chem. 2016, 88, 7894–7898. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Lu, D.-Q.; Liang, H.; Xie, S.; Luo, C.; Hu, M.; Xu, L.; Zhang, X.; Tan, W. Fluorescence Resonance Energy Transfer-Based DNA Tetrahedron Nanotweezer for Highly Reliable Detection of Tumor-Related MRNA in Living Cells. ACS Nano 2017, 11, 4060–4066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Li, Z.; Wang, H.; Zhuo, Y.; Yuan, R.; Chai, Y. DNA Nanomachine-Based Regenerated Sensing Platform: A Novel Electrochemiluminescence Resonance Energy Transfer Strategy for Ultra-High Sensitive Detection of MicroRNA from Cancer Cells. Nanoscale 2017, 9, 2310–2316. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Jie, G. An “on-off” Electrochemiluminescence Biosensor Based on DNA Nanotweezer Probe Coupled with Tripod Capture DNA for High Sensitive Detection of Pb2+. Sens. Actuators B Chem. 2021, 326, 128985. [Google Scholar] [CrossRef]
- Aitken, C.E.; Marshall, R.A.; Puglisi, J.D. An Oxygen Scavenging System for Improvement of Dye Stability in Single-Molecule Fluorescence Experiments. Biophys. J. 2008, 94, 1826–1835. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.; Uhm, H.; Tauzin, L.J.; Poddar, N.; Landes, C.F. Photobleaching Lifetimes of Cyanine Fluorophores Used for Single-Molecule Förster Resonance Energy Transfer in the Presence of Various Photoprotection Systems. ChemBioChem 2013, 14, 1075–1080. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, A.; Mahmoud, R.; Megalathan, A.; Pettit, S.; Dhakal, S. Multiplexed smFRET Nucleic Acid Sensing Using DNA Nanotweezers. Biosensors 2023, 13, 119. https://doi.org/10.3390/bios13010119
Kaur A, Mahmoud R, Megalathan A, Pettit S, Dhakal S. Multiplexed smFRET Nucleic Acid Sensing Using DNA Nanotweezers. Biosensors. 2023; 13(1):119. https://doi.org/10.3390/bios13010119
Chicago/Turabian StyleKaur, Anisa, Roaa Mahmoud, Anoja Megalathan, Sydney Pettit, and Soma Dhakal. 2023. "Multiplexed smFRET Nucleic Acid Sensing Using DNA Nanotweezers" Biosensors 13, no. 1: 119. https://doi.org/10.3390/bios13010119
APA StyleKaur, A., Mahmoud, R., Megalathan, A., Pettit, S., & Dhakal, S. (2023). Multiplexed smFRET Nucleic Acid Sensing Using DNA Nanotweezers. Biosensors, 13(1), 119. https://doi.org/10.3390/bios13010119