PtNPs/PEDOT:PSS-Modified Microelectrode Arrays for Detection of the Discharge of Head Direction Cells in the Retrosplenial Cortex of Rats under Dissociation between Visual and Vestibular Inputs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
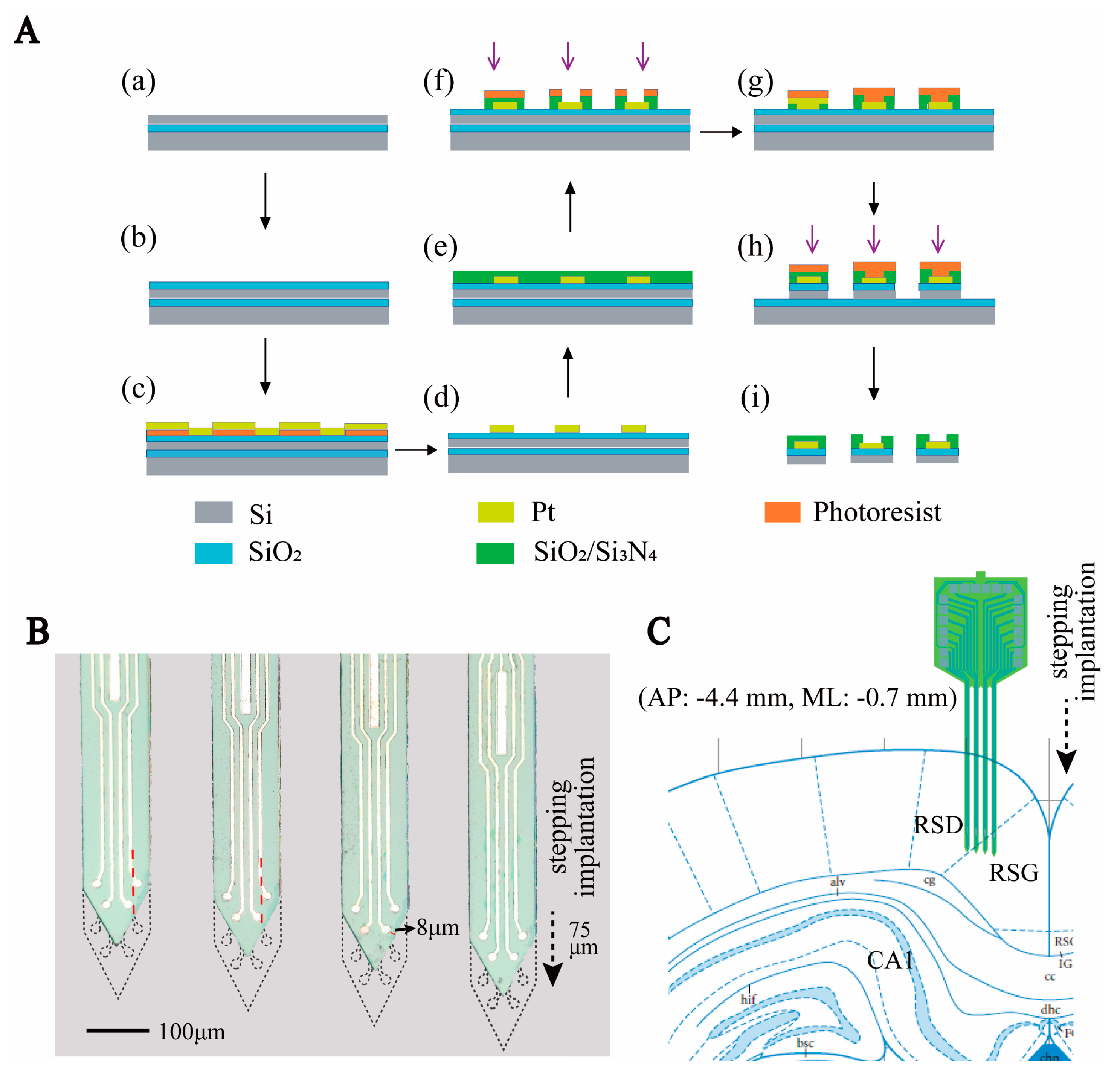
2.1. Design and Fabrication of the MEA
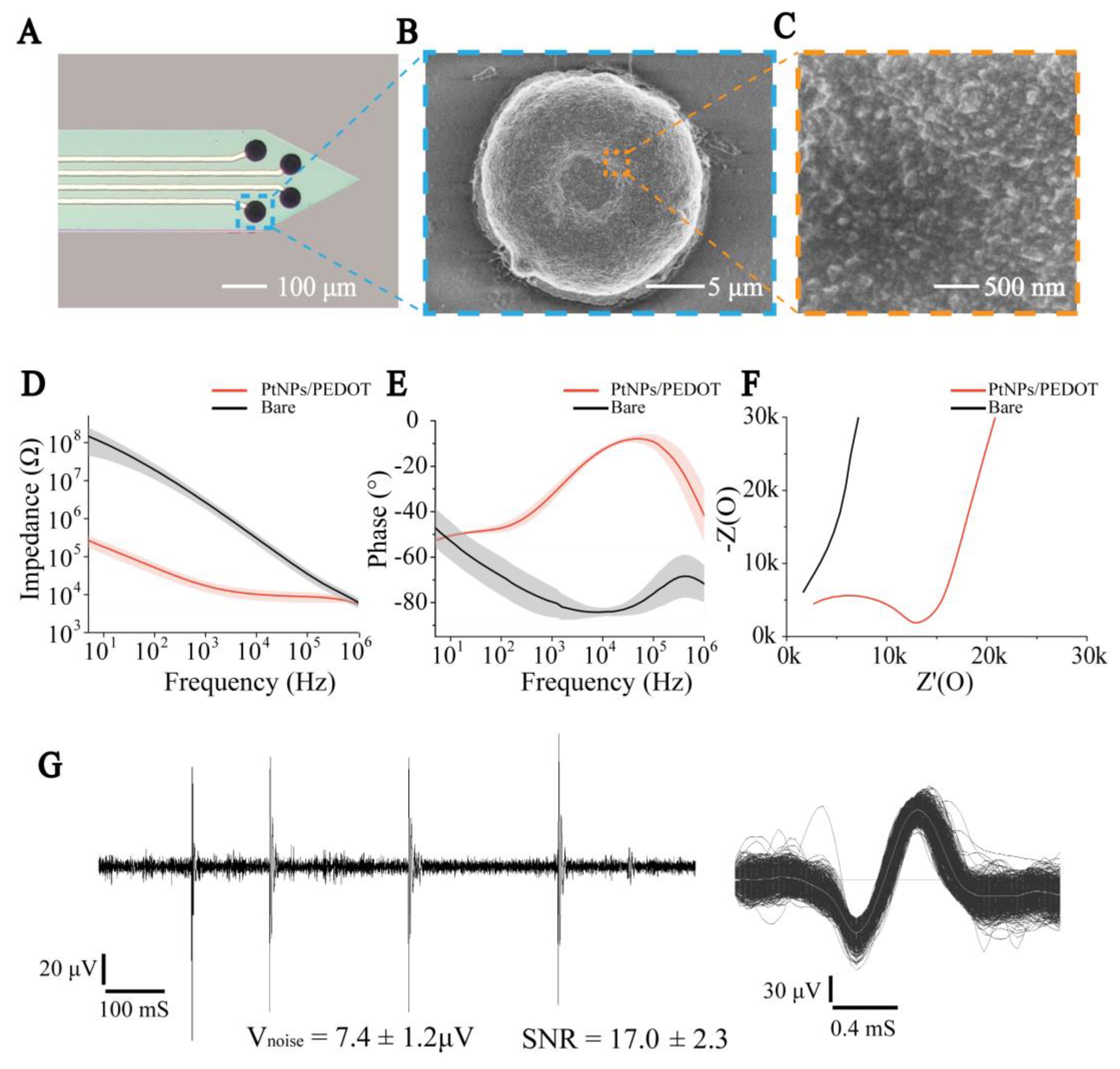
2.2. PtNPs/PEDOT:PSS Modification Method
2.3. Subjects and Surgery
2.4. Apparatus
2.5. Recording Setup
2.6. Behavioral Training and Testing Protocol
2.7. Identification and Analysis of HD Cells
3. Results
3.1. Characteristics of PtNPs/PEDOT:PSS Nanocomposites
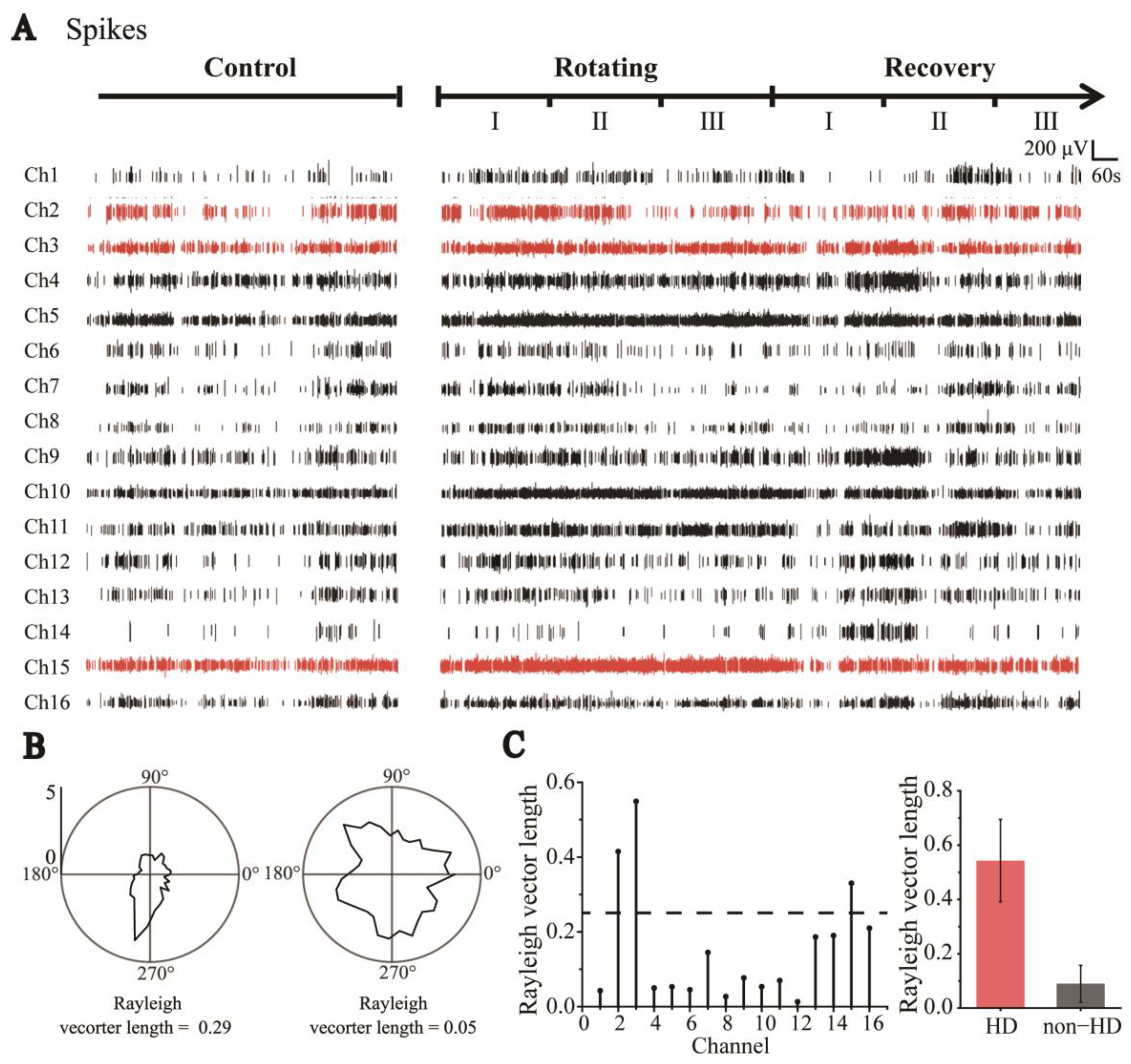
3.2. Identification of HD and Non-HD Cells
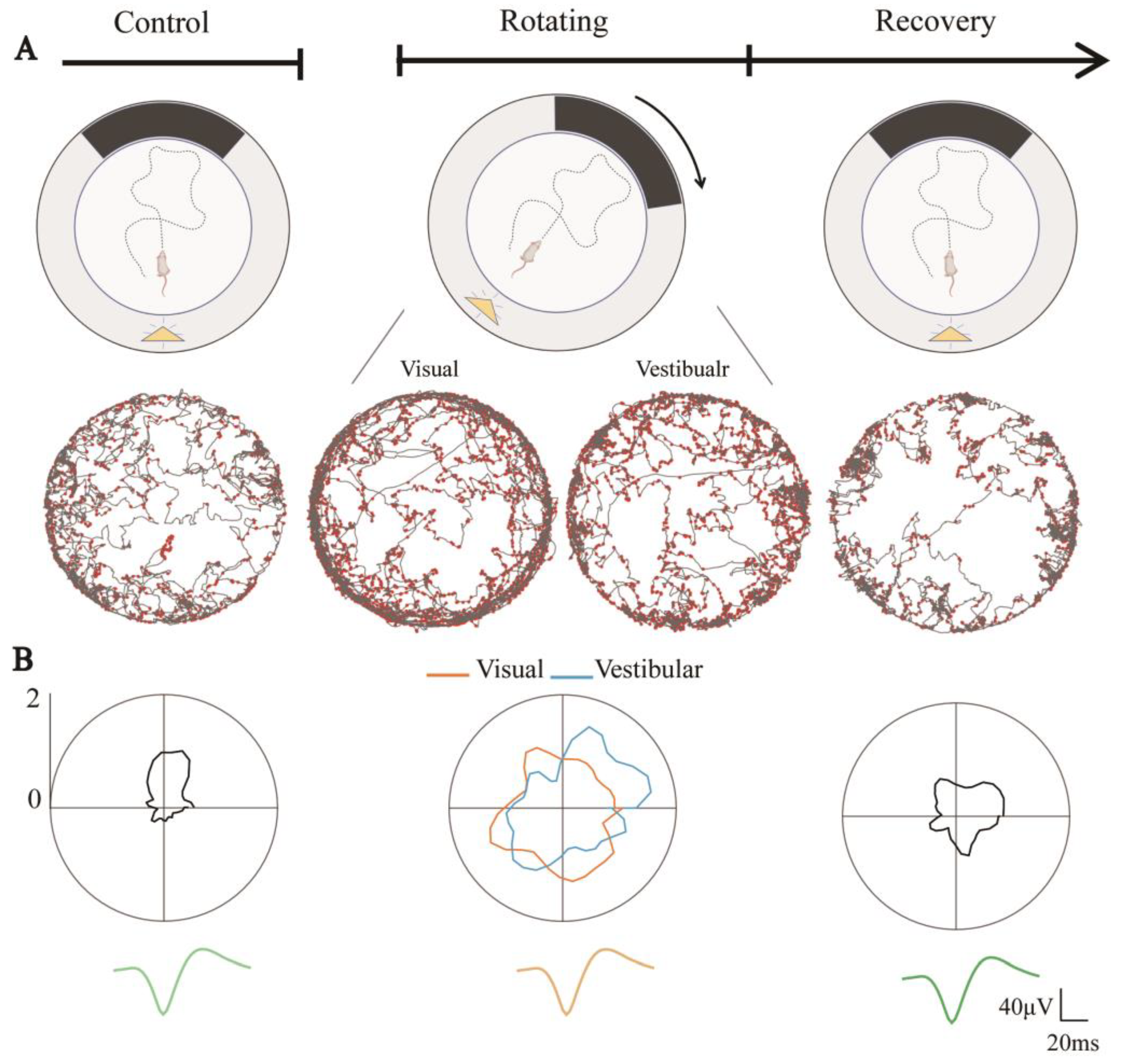
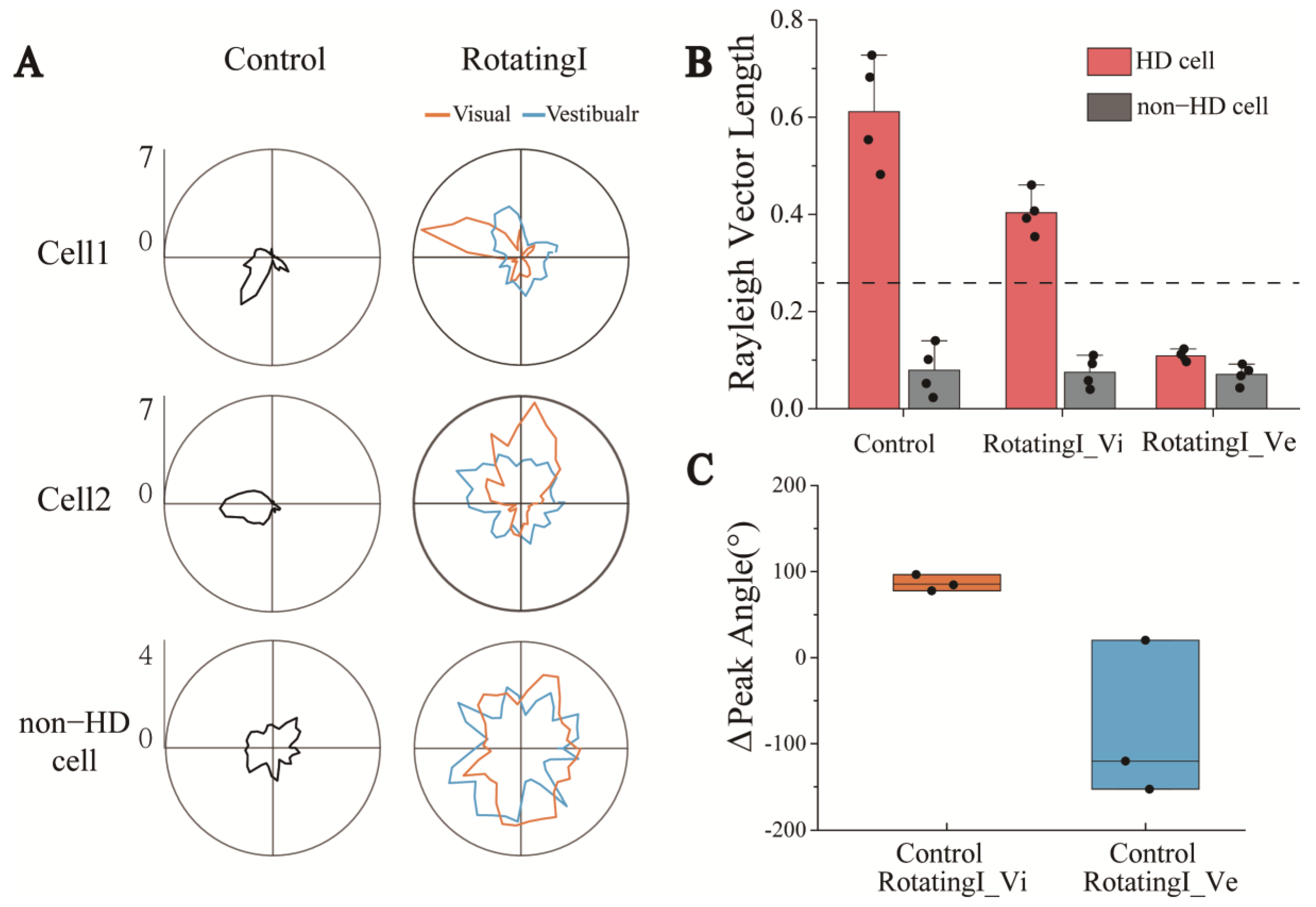
3.3. Degraded Directional Tuning of Head-Direction Cells Induced by Sensory Dissociation
3.4. Directional Tuning of HD Cells Is Dominated by Visual Rather than Vestibular Input during Short-Time Dissociated Navigation
3.5. Long-Term Dissociation between Visual and Vestibular Perception Degrades the Directional Tuning of HD Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aguirre, G.K.; D’Esposito, M. Topographical disorientation: A synthesis and taxonomy. Brain 1999, 122, 1613–1628. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Kawamura, M.; Shiota, J.; Kasahata, N.; Hirayama, K. Pure topographic disorientation due to right retrosplenial lesion. Neurology 1997, 49, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Taube, J.S.; Muller, R.U.; Ranck, J.B. Head-direction cells recorded from the postsubiculum in freely moving rats. 1. description and quantitative-analysis. J. Neurosci. 1990, 10, 420–435. [Google Scholar] [CrossRef] [PubMed]
- Angelaki, D.E.; Laurens, J. The head direction cell network: Attractor dynamics, integration within the navigation system, and three-dimensional properties. Curr. Opin. Neurobiol. 2020, 60, 136–144. [Google Scholar] [CrossRef]
- Stacho, M.; Manahan-Vaughan, D. Mechanistic flexibility of the retrosplenial cortex enables its contribution to spatial cognition. Trends Neurosci. 2022, 45, 284–296. [Google Scholar] [CrossRef]
- Wang, Q.X.; Sporns, O.; Burkhalter, A. Network Analysis of Corticocortical Connections Reveals Ventral and Dorsal Processing Streams in Mouse Visual Cortex. J. Neurosci. 2012, 32, 4386–4399. [Google Scholar] [CrossRef]
- Keshavarzi, S.; Bracey, E.F.; Faville, R.A.; Campagner, D.; Tyson, A.L.; Lenzi, S.C.; Branco, T.; Margrie, T.W. Multisensory coding of angular head velocity in the cortex. Neuron 2022, 110, 532–543.e9. [Google Scholar] [CrossRef]
- Kim, J.G.; Aminoff, E.M.; Kastner, S.; Behrmann, M. A Neural Basis for Developmental Topographic Disorientation. J. Neurosci. 2015, 35, 12954–12969. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, R.; Tanaka, Y.; Nakano, I. Heading Disorientation: A New Test and a Possible Underlying Mechanism. Eur. Neurol. 2010, 63, 87–93. [Google Scholar] [CrossRef]
- Goodridge, J.P.; Dudchenko, P.A.; Worboys, K.A.; Golob, E.J.; Taube, J.S. Cue control and head direction cells. Behav. Neurosci. 1998, 112, 749–761. [Google Scholar] [CrossRef]
- Redish, A.D.; Elga, A.N.; Touretzky, D.S. A coupled attractor model of the rodent head direction system. Netw. Comput. Neural Syst. 1996, 7, 671–685. [Google Scholar] [CrossRef]
- Skaggs, W.E.; Knierim, J.J.; Kudrimoti, H.S.; McNaughton, B.L. A model of the neural basis of the rat’s sense of direction. Adv. Neural Inf. Process. Syst. 1995, 7, 173–180. [Google Scholar]
- Mao, D.; Molina, L.A.; Bonin, V.; McNaughton, B.L. Vision and Locomotion Combine to Drive Path Integration Sequences in Mouse Retrosplenial Cortex. Curr. Biol. 2020, 30, 1680–1688.e4. [Google Scholar] [CrossRef]
- Chen, G.F.; King, J.A.; Burgess, N.; O’Keefe, J. How vision and movement combine in the hippocampal place code. Proc. Natl. Acad. Sci. USA 2013, 110, 378–383. [Google Scholar] [CrossRef]
- Zheng, Q.H.; Zhou, L.X.; Gu, Y. Temporal synchrony effects of optic flow and vestibular inputs on multisensory heading perception. Cell Rep. 2021, 37, 15. [Google Scholar] [CrossRef] [PubMed]
- So, P.T.C.; Dong, C.Y.; Masters, B.R.; Berland, K.M. Two-photon excitation fluorescence microscopy. Annu. Rev. Biomed. Eng. 2000, 2, 399–429. [Google Scholar] [CrossRef]
- Holtmaat, A.; Bonhoeffer, T.; Chow, D.K.; Chuckowree, J.; De Paola, V.; Hofer, S.B.; Hubener, M.; Keck, T.; Knott, G.; Lee, W.C.A.; et al. Long-term, high-resolution imaging in the mouse neocortex through a chronic cranial window. Nat. Protoc. 2009, 4, 1128–1144. [Google Scholar] [CrossRef]
- Kornienko, O.; Latuske, P.; Bassler, M.; Kohler, L.; Allen, K. Non-rhythmic head-direction cells in the parahippocampal region are not constrained by attractor network dynamics. eLife 2018, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.H.; Song, Y.L.; Lu, B.T.; Wang, Y.D.; Dai, Y.C.; Xie, J.Y.; He, E.H.; Xu, Z.J.; Yang, G.C.; Mo, F.; et al. PtNPs/PEDOT:PSS-Modified Microelectrode Arrays Reveal Electrophysiological Activities of Different Neurons in Medial Amygdala of Mice under Innate Fear. Front. Neurosci. 2022, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.J.; Song, Y.L.; Wang, L.; Zhang, S.; Luo, J.P.; Xu, S.W.; Cai, X.X. An implantable microelectrode array for simultaneous L-glutamate and electrophysiological recordings in vivo. Microsyst. Nanoeng. 2015, 1, 6. [Google Scholar] [CrossRef]
- Wang, Y.D.; Song, Y.L.; Dai, Y.C.; Li, X.R.; Xie, J.Y.; Luo, J.P.; Yang, C.; Fan, P.H.; Xiao, G.H.; Luo, Y.; et al. The burst of electrophysiological signals in the suprachiasmatic nucleus of mouse during the arousal detected by microelectrode arrays. Front. Bioeng. Biotechnol. 2022, 10, 12. [Google Scholar] [CrossRef]
- Ferguson, M.; Sharma, D.; Ross, D.; Zhao, F. A Critical Review of Microelectrode Arrays and Strategies for Improving Neural Interfaces. Adv. Healthc. Mater. 2019, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Song, Y.L.; Wang, M.X.; Zhang, Z.M.; Fan, X.Y.; Song, X.T.; Zhuang, P.; Yue, F.; Chan, P.; Cai, X.X. A silicon based implantable microelectrode array for electrophysiological and dopamine recording from cortex to striatum in the non-human primate brain. Biosens. Bioelectron. 2016, 85, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Kozai, T.D.Y.; Catt, K.; Du, Z.H.; Na, K.; Srivannavit, O.; Haque, R.U.M.; Seymour, J.; Wise, K.D.; Yoon, E.; Cui, X.T. Chronic In Vivo Evaluation of PEDOT/CNT for Stable Neural Recordings. IEEE Trans. Biomed. Eng. 2016, 63, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Dijk, G.; Rutz, A.L.; Malliaras, G.G. Stability of PEDOT:PSS-Coated Gold Electrodes in Cell Culture Conditions. Adv. Mater. Technol. 2020, 5, 6. [Google Scholar] [CrossRef]
- Kadir, S.N.; Goodman, D.F.M.; Harris, K.D. High-Dimensional Cluster Analysis with the Masked EM Algorithm. Neural Comput. 2014, 26, 2379–2394. [Google Scholar] [CrossRef] [PubMed]
- Harris, K.D.; Henze, D.A.; Csicsvari, J.; Hirase, H.; Buzsaki, G. Accuracy of tetrode spike separation as determined by simultaneous intracellular and extracellular measurements. J. Neurophysiol. 2000, 84, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, P.Y.; Casali, G.; Spieser, L.; Page, H.; Overington, D.; Jeffery, K. An independent, landmark-dominated head-direction signal in dysgranular retrosplenial cortex. Nat. Neurosci. 2017, 20, 173–175. [Google Scholar] [CrossRef]
- Butler, W.N.; Smith, K.S.; van der Meer, M.A.A.; Taube, J.S. The Head-Direction Signal Plays a Functional Role as a Neural Compass during Navigation. Curr. Biol. 2017, 27, 2406. [Google Scholar] [CrossRef]
- Bassett, J.P.; Wills, T.J.; Cacucci, F. Self-Organized Attractor Dynamics in the Developing Head Direction Circuit. Curr. Biol. 2018, 28, 609–615.e3. [Google Scholar] [CrossRef]
- Johnson, D.H.; Gruner, C.M. Information-theoretic analysis of neural coding. In Proceedings of the IEEE International Conference on Acoustics, Speech and Signal Processing (ICASSP 98), Seattle, WA, USA, 12–15 May 1998; pp. 1937–1940. [Google Scholar]
- Olypher, A.V.; Lansky, P.; Muller, R.U.; Fenton, A.A. Quantifying location-specific information in the discharge of rat hippocampal place cells. J. Neurosci. Methods 2003, 127, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Hennestad, E.; Witoelar, A.; Chambers, A.R.; Vervaeke, K. Mapping vestibular and visual contributions to angular head velocity tuning in the cortex. Cell Rep. 2021, 37, 23. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Adachi, A.; Shah, K.K.; Hirokawa, J.D.; Magani, P.S.; Maimon, G. A neural circuit architecture for angular integration in Drosophila. Nature 2017, 546, 101–106. [Google Scholar] [CrossRef]
- Wang, Q.X.; Burkhalter, A. Area map of mouse visual cortex. J. Comp. Neurol. 2007, 502, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Blair, H.T.; Sharp, P.E. Anticipatory head direction signals in anterior thalamus—Evidence for a thalamocortical circuit that integrates angular head motion to compute head direction. J. Neurosci. 1995, 15, 6260–6270. [Google Scholar] [CrossRef]
- Park, E.H.; Keeley, S.; Savin, C.; Ranck, J.B.; Fenton, A.A. How the Internally Organized Direction Sense Is Used to Navigate. Neuron 2019, 101, 285–293.e5. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Wang, Y.; Xu, Z.; Zhang, X.; Ruan, W.; Mo, F.; Lu, B.; Fan, P.; Dai, Y.; He, E.; et al. PtNPs/PEDOT:PSS-Modified Microelectrode Arrays for Detection of the Discharge of Head Direction Cells in the Retrosplenial Cortex of Rats under Dissociation between Visual and Vestibular Inputs. Biosensors 2023, 13, 496. https://doi.org/10.3390/bios13050496
Yang G, Wang Y, Xu Z, Zhang X, Ruan W, Mo F, Lu B, Fan P, Dai Y, He E, et al. PtNPs/PEDOT:PSS-Modified Microelectrode Arrays for Detection of the Discharge of Head Direction Cells in the Retrosplenial Cortex of Rats under Dissociation between Visual and Vestibular Inputs. Biosensors. 2023; 13(5):496. https://doi.org/10.3390/bios13050496
Chicago/Turabian StyleYang, Gucheng, Yiding Wang, Zhaojie Xu, Xue Zhang, Wang Ruan, Fan Mo, Botao Lu, Penghui Fan, Yuchuan Dai, Enhui He, and et al. 2023. "PtNPs/PEDOT:PSS-Modified Microelectrode Arrays for Detection of the Discharge of Head Direction Cells in the Retrosplenial Cortex of Rats under Dissociation between Visual and Vestibular Inputs" Biosensors 13, no. 5: 496. https://doi.org/10.3390/bios13050496
APA StyleYang, G., Wang, Y., Xu, Z., Zhang, X., Ruan, W., Mo, F., Lu, B., Fan, P., Dai, Y., He, E., Song, Y., Wang, C., Liu, J., & Cai, X. (2023). PtNPs/PEDOT:PSS-Modified Microelectrode Arrays for Detection of the Discharge of Head Direction Cells in the Retrosplenial Cortex of Rats under Dissociation between Visual and Vestibular Inputs. Biosensors, 13(5), 496. https://doi.org/10.3390/bios13050496