C-Type Natriuretic Peptide (CNP) Inhibition of Interferon-γ-Mediated Gene Expression in Human Endothelial Cells In Vitro



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Flow Cytometry
2.4. cGMP Assay
2.5. Mass Spectrometry for Tryptophan Metabolites
2.6. Data Analysis
3. Results
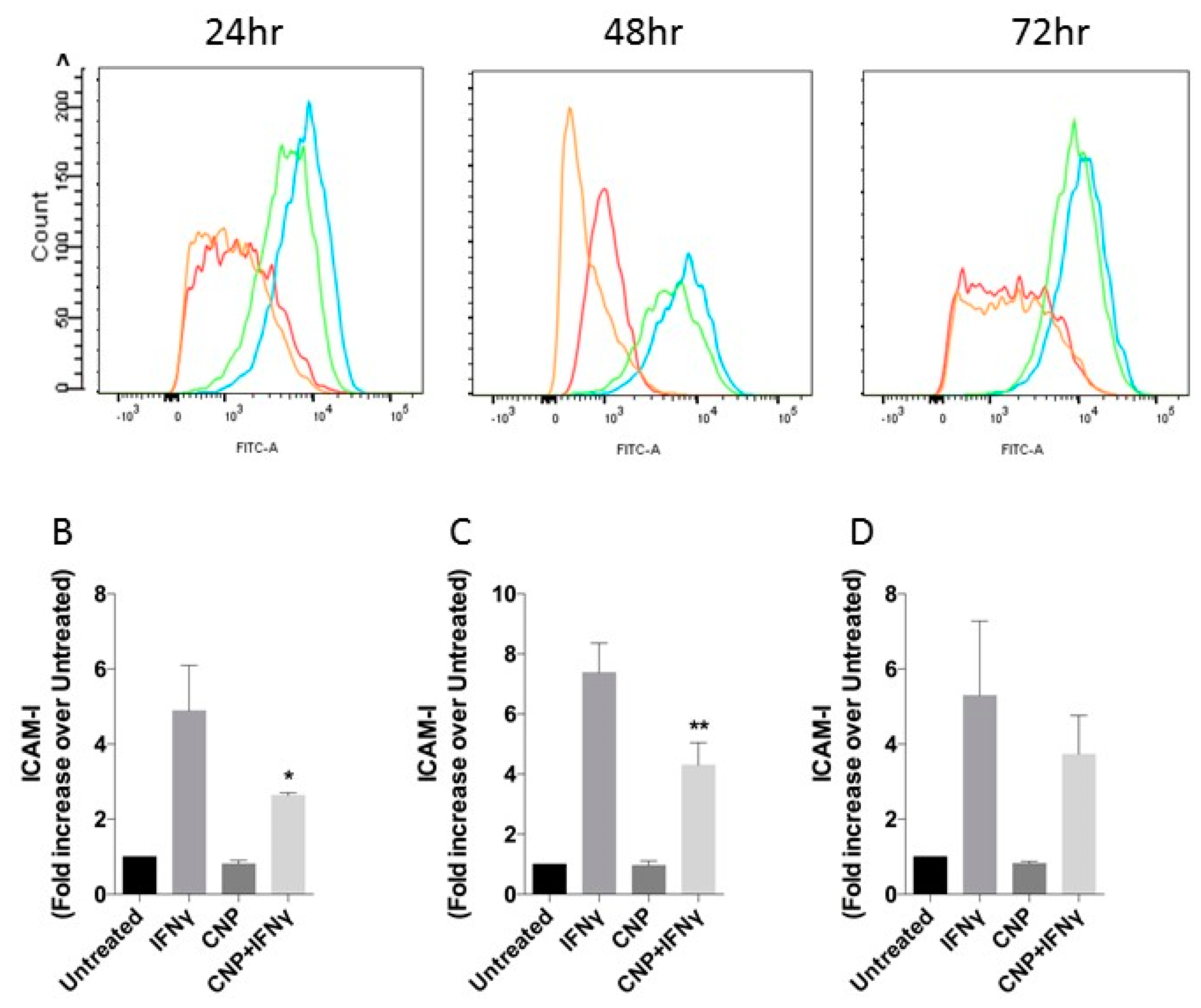
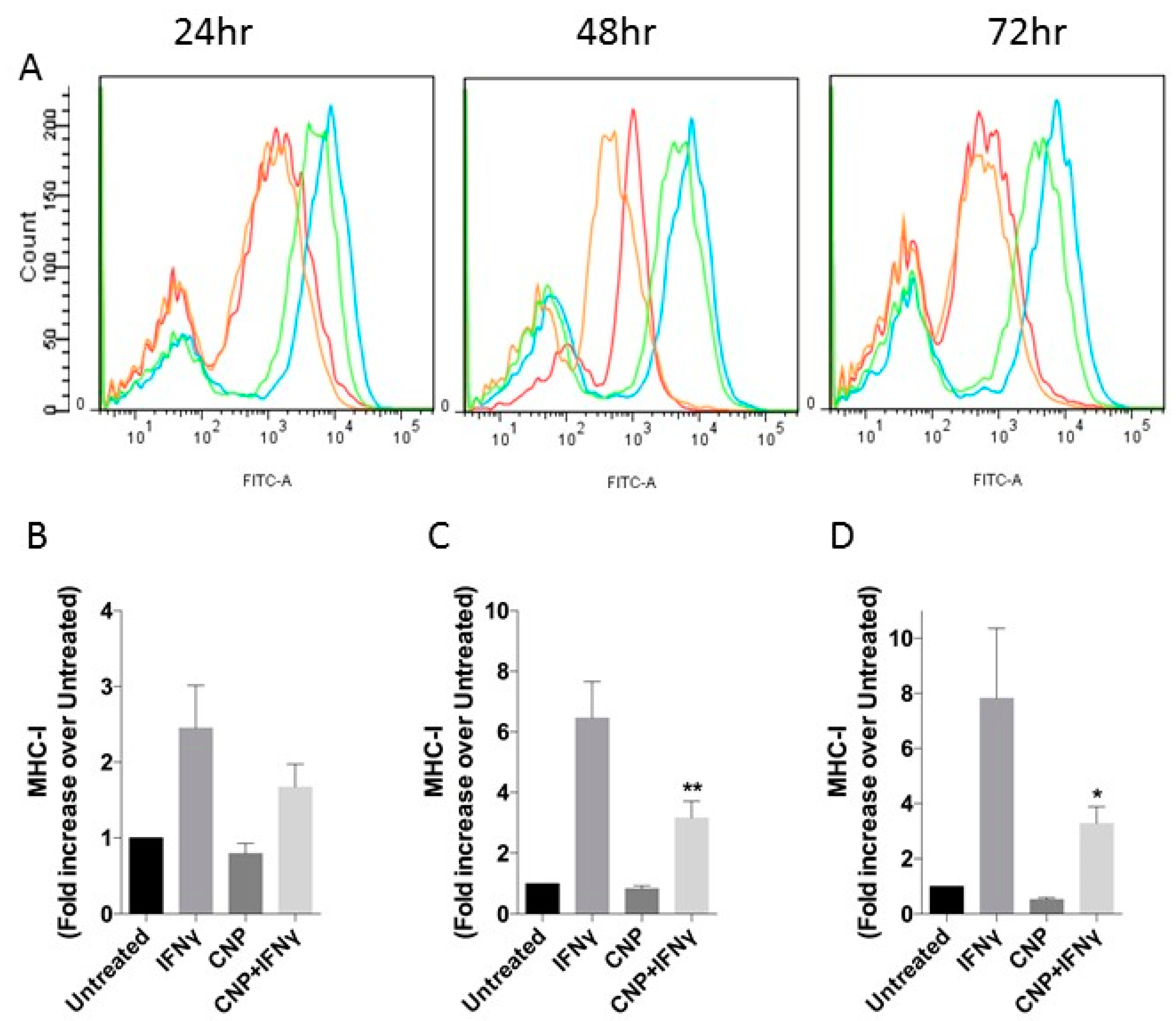
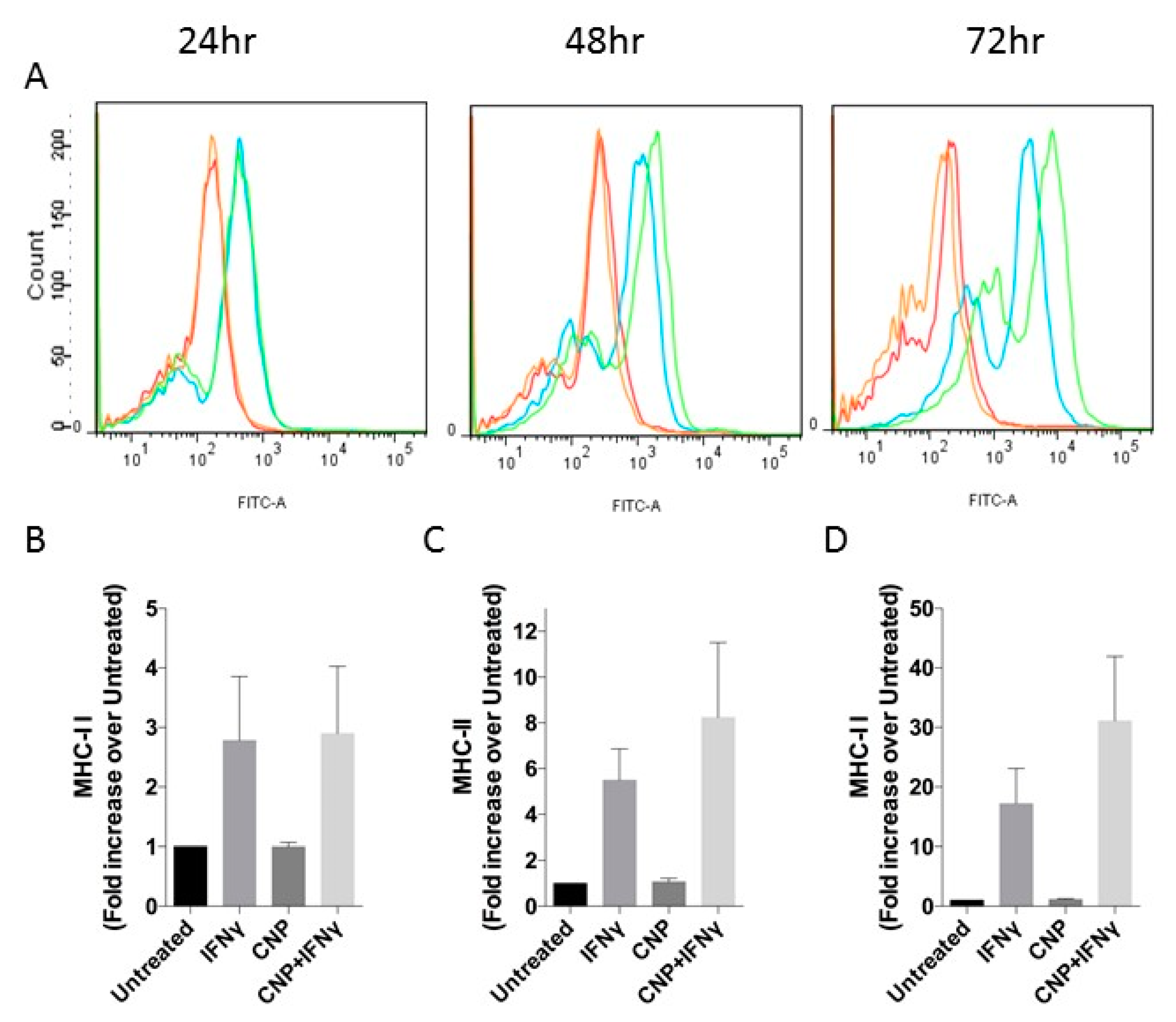
3.1. CNP Reduces IFN-γ Mediated Expression of Pro-Inflammatory Molecules on the Surface of HUVEC.
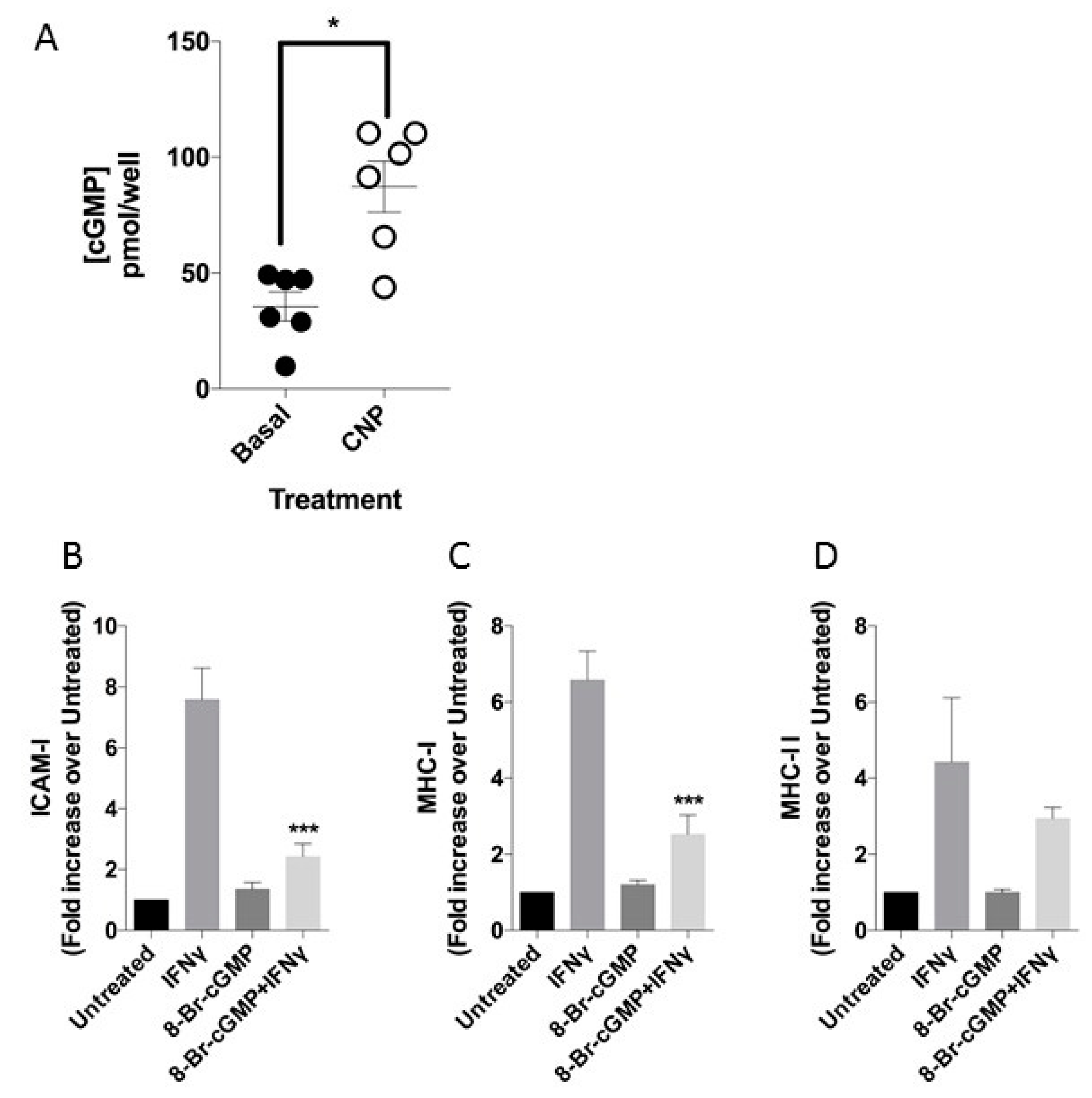
3.2. CNP Induces cGMP Release in HUVEC
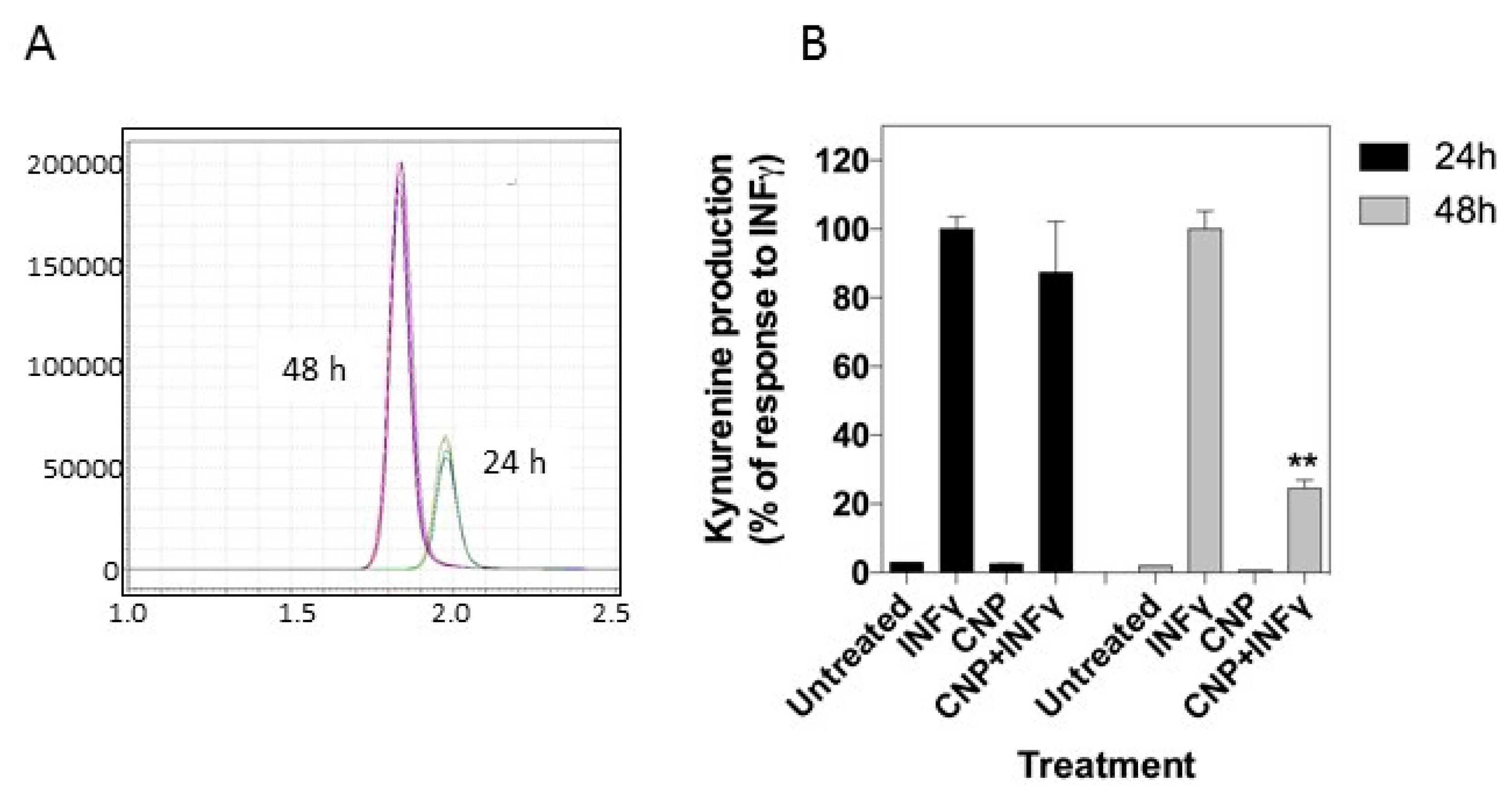
3.3. CNP Inhibits IFN-γ Induced Upregulation of Tryptophan Metabolism
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mozaffarian, D.; Benjamin, E.J.; Go, A.S.; Arnett, D.K.; Blaha, M.J.; Cushman, M.D.; Das, S.R.; De Ferranti, S.; Cushman, M.; Despr’s, J.P.; et al. Executive Summary: Heart Disease and Stroke Statistics-2016 Update: A Report From the American Heart Association. Circulation 2016, 133, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, E.J.; Virani, S.S.; Callaway, C.W.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Chiuve, S.E.; Cushman, M.; Delling, F.N.; Deo, R. Heart Disease and Stroke Statistics-2018 Update: A Report From the American Heart Association. Circulation 2018, 137, e67–e492. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K.; Libby, P.; Tabas, I. Inflammation and plaque vulnerability. J. Intern. Med. 2015, 278, 483–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, F.A.; Murphy, R.P.; Cummins, P.M. Thrombomodulin and the vascular endothelium: Insights into functional, regulatory, and therapeutic aspects. Am. J. Physiol. Heart Circ. Physiol. 2013, 304, H1585–H1597. [Google Scholar] [CrossRef] [PubMed]
- Ketelhuth, D.F.; Hansson, G.K. Cellular immunity, low-density lipoprotein and atherosclerosis: Break of tolerance in the artery wall. Thromb. Haemost. 2011, 106, 779–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thornhill, M.H.; Wellicome, S.M.; Mahiouz, D.L.; Lanchbury, J.S.S.; Kyan-Aung, U.; Haskard, D.O. Tumor necrosis factor combines with IL-4 or IFN-gamma to selectively enhance endothelial cell adhesiveness for T cells. The contribution of vascular cell adhesion molecule-1-dependent and -independent binding mechanisms. J. Immunol. 1991, 146, 592–598. [Google Scholar] [PubMed]
- Lidington, E.A.; Moyes, D.L.; McCormack, A.M.; Rose, M.L. A comparison of primary endothelial cells and endothelial cell lines for studies of immune interactions. Transpl. Immunol. 1999, 7, 239–246. [Google Scholar] [CrossRef]
- Mbongue, J.C.; Nicholas, D.A.; Torrez, T.W.; Kim, N.-S.; Firek, A.F.; Langridge, W.H.R. The Role of Indoleamine 2,3-Dioxygenase in Immune Suppression and Autoimmunity. Vaccines 2015, 3, 703–729. [Google Scholar] [CrossRef] [PubMed]
- Mellor, A.L.; Keskin, D.B.; Johnson, T.; Chandler, P.; Munn, D.H. Cells expressing indoleamine 2,3-dioxygenase inhibit T cell responses. J. Immunol. 2002, 168, 3771–3776. [Google Scholar] [CrossRef] [PubMed]
- Mellor, A.L.; Munn, D.H. Tryptophan catabolism and T-cell tolerance: Immunosuppression by starvation? Immunol. Today 1999, 20, 469–473. [Google Scholar] [CrossRef]
- Taylor, M.W.; Feng, G.S. Relationship between interferon-gamma, indoleamine 2,3-dioxygenase, and tryptophan catabolism. FASEB J. 1991, 5, 2516–2522. [Google Scholar] [CrossRef] [PubMed]
- Curry, F.-R.E.; Clark, J.F.; Jiang, Y.; Kim, M.-H.; Adamson, R.H.; Simon, S.I. The role of atrial natriuretic peptide to attenuate inflammation in a mouse skin wound and individually perfused rat mesenteric microvessels. Physiol. Rep. 2016, 4, e12968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoury, E.E.; Kinaneh, S.; Aronson, D.; Amir, O.; Ghanim, D.; Volinsky, N.; Azzam, Z.; Abassi, Z. Natriuretic peptides system in the pulmonary tissue of rats with heart failure: Potential involvement in lung edema and inflammation. Oncotarget 2018, 9, 21715–21730. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M. Endothelial actions of atrial and B-type natriuretic peptides. Br. J. Pharmacol. 2012, 166, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, A.; Nojiri, T.; Konishi, K.; Tokudome, T.; Miura, K.; Hosoda, H.; Hino, J.; Miyazato, M.; Kyomoto, Y.; Asai, K. Atrial natriuretic peptide protects against bleomycin-induced pulmonary fibrosis via vascular endothelial cells in mice: ANP for pulmonary fibrosis. Respir. Res. 2017, 18, 1. [Google Scholar] [CrossRef] [PubMed]
- Sudoh, T.; Minamino, N.; Kangawa, K.; Matsuo, H. C-type natriuretic peptide (CNP): A new member of natriuretic peptide family identified in porcine brain. Biochem. Biophys. Res. Commun. 1990, 168, 863–870. [Google Scholar] [CrossRef]
- Ahluwalia, A.; Hobbs, A.J. Endothelium-derived C-type natriuretic peptide: More than just a hyperpolarizing factor. Trends Pharmacol. Sci. 2005, 26, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Scotland, R.S.; Ahluwalia, A.; Hobbs, A.J. C-type natriuretic peptide in vascular physiology and disease. Pharmacol. Ther. 2005, 105, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Izumiya, Y.; Araki, S.; Usuku, H.; Rokutanda, T.; Hanatani, S.; Ogawa, H. Chronic C-Type Natriuretic Peptide Infusion Attenuates Angiotensin II-Induced Myocardial Superoxide Production and Cardiac Remodeling. Int. J. Vasc. Med. 2012, 2012, 246058. [Google Scholar] [CrossRef] [PubMed]
- Scotland, R.S.; Cohen, M.; Foster, P.; Lovell, M.; Mathur, A.; Ahluwalia, A.; Hobbs, A.J. C-type natriuretic peptide inhibits leukocyte recruitment and platelet-leukocyte interactions via suppression of P-selectin expression. Proc. Natl. Acad. Sci. USA 2005, 102, 14452–14457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, J.; Haruno, A.; Asada, Y.; Nishida, T.; Saito, Y.; Matsuda, T.; Ueno, H. Local expression of C-type natriuretic peptide suppresses inflammation, eliminates shear stress-induced thrombosis, and prevents neointima formation through enhanced nitric oxide production in rabbit injured carotid arteries. Circ. Res. 2002, 91, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Wang, J.; Hu, B.; Lu, L.; Xuan, Q.; Qin, Y. Increased urinary C-type natriuretic peptide excretion may be an early marker of renal tubulointerstitial fibrosis. Peptides 2012, 37, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Y.; Li, G.; Li, B.; Liu, Y.; Li, X.; Liu, A. Inhibitory effects of C-type natriuretic peptide on the differentiation of cardiac fibroblasts, and secretion of monocyte chemoattractant protein-1 and plasminogen activator inhibitor-1. Mol. Med. Rep. 2015, 11, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Ichiki, T.; Boerrigter, G.; Huntley, B.K.; Sangaralingham, S.J.; McKie, P.M.; Harty, G.J.; Harders, G.E.; Burnett, J.C., Jr. Differential expression of the pro-natriuretic peptide convertases corin and furin in experimental heart failure and atrial fibrosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R102–R109. [Google Scholar] [CrossRef] [PubMed]
- Burley, D.S.; Hamid, S.A.; Baxter, G.F. Cardioprotective actions of peptide hormones in myocardial ischemia. Heart Fail. Rev. 2007, 12, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Soeki, T.; Kishimoto, I.; Okumura, H.; Tokudome, T.; Horio, T.; Mori, K.; Kangawa, K. C-type natriuretic peptide, a novel antifibrotic and antihypertrophic agent, prevents cardiac remodeling after myocardial infarction. J. Am. Coll Cardiol. 2005, 45, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Ichiki, T.; Schirger, J.A.; Huntley, B.K.; Brozovich, F.V.; Maleszewski, J.J.; Sandberg, S.M.; Sangaralingham, S.J.; Park, S.J.; Burnett, J.C., Jr. Cardiac fibrosis in end-stage human heart failure and the cardiac natriuretic peptide guanylyl cyclase system: Regulation and therapeutic implications. J. Mol. Cell Cardiol. 2014, 75, 199–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangaralingham, S.J.; McKie, P.M.; Ichiki, T.; Scott, C.G.; Heublein, D.M.; Chen, H.H.; Bailey, K.R.; Redfield, M.M.; Rodeheffer, R.J.; Burnett, J.C., Jr. Circulating C-type natriuretic peptide and its relationship to cardiovascular disease in the general population. Hypertension 2015, 65, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, E.A.; Nachman, R.L.; Becker, C.G.; Minick, C.R. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J. Clin. Investing 1973, 52, 2745–2756. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.; Ainsworth, M.; Yacoub, M.; Rose, M. Ligation of ICAM-1 on endothelial cells leads to expression of VCAM-1 via a nuclear factor-kappaB-independent mechanism. J. Immunol. 1999, 162, 2990–2996. [Google Scholar] [PubMed]
- Pober, J.S.; Gimbrone, M.A.; Lapierre, L.A.; Mendrick, D.; Fiers, W.; Rothlein, R.; Springer, T. A Overlapping patterns of activation of human endothelial cells by interleukin 1, tumor necrosis factor, and immune interferon. J. Immunol. 1986, 137, 1893–1896. [Google Scholar] [PubMed]
- Thornhill, M.H.; Haskard, D.O. IL-4 regulates endothelial cell activation by IL-1, tumor necrosis factor, or IFN-gamma. J. Immunol. 1990, 145, 865–872. [Google Scholar] [PubMed]
- Del Ry, S. C-type natriuretic peptide: A new cardiac mediator. Peptides 2013, 40, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, L.R. Regulation and therapeutic targeting of peptide-activated receptor guanylyl cyclases. Pharmacol. Ther. 2011, 130, 71–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbs, A.; Foster, P.; Prescott, C.; Scotland, R.; Ahluwalia, A. Natriuretic peptide receptor-C regulates coronary blood flow and prevents myocardial ischemia/reperfusion injury: Novel cardioprotective role for endothelium-derived C-type natriuretic peptide. Circulation 2004, 110, 1231–1235. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.D.; Nilsson, H.; Ahluwalia, A.; Hobbs, A.J. Release of C-type natriuretic peptide accounts for the biological activity of endothelium-derived hyperpolarizing factor. Proc. Natl. Acad. Sci. USA 2003, 100, 1426–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, C.; Wolf, S. ICAM-1 signaling in endothelial cells. Pharmacol. Rep. 2009, 61, 22–32. [Google Scholar] [CrossRef]
- Cole, J.E.; Astola, N.; Cribbs, A.P.; Goddard, M.E.; Park, I.; Green, P.; Davies, A.H.; Williams, R.O.; Feldmann, M.; Monaco, C. Indoleamine 2,3-dioxygenase-1 is protective in atherosclerosis and its metabolites provide new opportunities for drug development. Proc. Natl. Acad. Sci. USA 2015, 112, 13033–13038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadhu, C.; Ting, H.J.; Lipsky, B.; Hensley, K.; Garcia-Martinez, L.F.; Simon, S.I.; Staunton, D.E. CD11c/CD18: Novel ligands and a role in delayed-type hypersensitivity. J. Leukoc. Biol. 2007, 81, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, K.A.; Ovchinnikova, O.; Berg, M.; Baumgartner, R.; Agardh, H.; Pirault, J.; Gister, A.; Assinger, A.; Laguna-Fernandez, A.; Back, M. Inhibition of indoleamine 2,3-dioxygenase promotes vascular inflammation and increases atherosclerosis in Apoe-/- mice. Cardiovasc. Res. 2015, 106, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Osawa, H.; Yamabe, H.; Kaizuka, M.; Tamura, N.; Tsunoda, S.; Baba, Y.; Shirato, K.; Tateyama, F.; Okumura, K. C-Type natriuretic peptide inhibits proliferation and monocyte chemoattractant protein-1 secretion in cultured human mesangial cells. Nephron 2000, 86, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Dansky, H.M.; Barlow, C.B.; Lominska, C.; Sikes, J.L.; Kao, C.; Weinsaft, J.; Cybulsky, M.I.; Smith, J.D. Adhesion of monocytes to arterial endothelium and initiation of atherosclerosis are critically dependent on vascular cell adhesion molecule-1 gene dosage. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1662–1667. [Google Scholar] [CrossRef] [PubMed]
- Gewaltig, J.; Kummer, M.; Koella, C.; Cathomas, G.; Biedermann, B.C. Requirements for CD8 T-cell migration into the human arterial wall. Hum. Pathol. 2008, 39, 1756–1762. [Google Scholar] [CrossRef] [PubMed]
- Rautureau, Y.; Gowers, I.; Wheeler-Jones, C.P.D.; Baxter, G.F. C-type natriuretic peptide regulation of guanosine-3′,5′-cyclic monophosphate production in human endothelial cells. Auton. Autacoid Pharmacol. 2010, 30, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, H.; Froese, A.; Jonsson, P.; Schmidt, H.; Gorelik, J.; Nikolaev, V.O. Distinct submembrane localisation compartmentalises cardiac NPR1 and NPR2 signalling to cGMP. Nat. Commun. 2018, 9, 2446. [Google Scholar] [CrossRef] [PubMed]
- Moyes, A.J.; Khambata, R.S.; Villar, I.; Bubb, K.J.; Baliga, R.S.; Lumsden, N.G.; Xiao, F.; Gane, P.J.; Rebstock, A.-S.; Worthington, R.J.; et al. Endothelial C-type natriuretic peptide maintains vascular homeostasis. J. Clin. Investing 2014, 124, 4039–4051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, M.A.; Elesgaray, R.; Balaszczuk, A.M.; Arranz, C. Role of NPR-C natriuretic receptor in nitric oxide system activation induced by atrial natriuretic peptide. Regul. Pept. 2006, 135, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Mcdouall, R.M.; Batten, P.; Mccormack, A.; Yacoub, M.H.; Rose, M.L. MHC class II expression on human heart microvascular endothelial cells: Exquisite sensitivity to interferon-gamma and natural killer cells. Transplantation 1997, 64, 1175–1180. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Day, A.; Jameson, Z.; Hyde, C.; Simbi, B.; Fowkes, R.; Lawson, C. C-Type Natriuretic Peptide (CNP) Inhibition of Interferon-γ-Mediated Gene Expression in Human Endothelial Cells In Vitro. Biosensors 2018, 8, 86. https://doi.org/10.3390/bios8030086
Day A, Jameson Z, Hyde C, Simbi B, Fowkes R, Lawson C. C-Type Natriuretic Peptide (CNP) Inhibition of Interferon-γ-Mediated Gene Expression in Human Endothelial Cells In Vitro. Biosensors. 2018; 8(3):86. https://doi.org/10.3390/bios8030086
Chicago/Turabian StyleDay, Amy, Zoe Jameson, Carolyn Hyde, Bigboy Simbi, Robert Fowkes, and Charlotte Lawson. 2018. "C-Type Natriuretic Peptide (CNP) Inhibition of Interferon-γ-Mediated Gene Expression in Human Endothelial Cells In Vitro" Biosensors 8, no. 3: 86. https://doi.org/10.3390/bios8030086
APA StyleDay, A., Jameson, Z., Hyde, C., Simbi, B., Fowkes, R., & Lawson, C. (2018). C-Type Natriuretic Peptide (CNP) Inhibition of Interferon-γ-Mediated Gene Expression in Human Endothelial Cells In Vitro. Biosensors, 8(3), 86. https://doi.org/10.3390/bios8030086