Lateral Flow Immunoassay for Rapid Detection of Grapevine Leafroll-Associated Virus

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
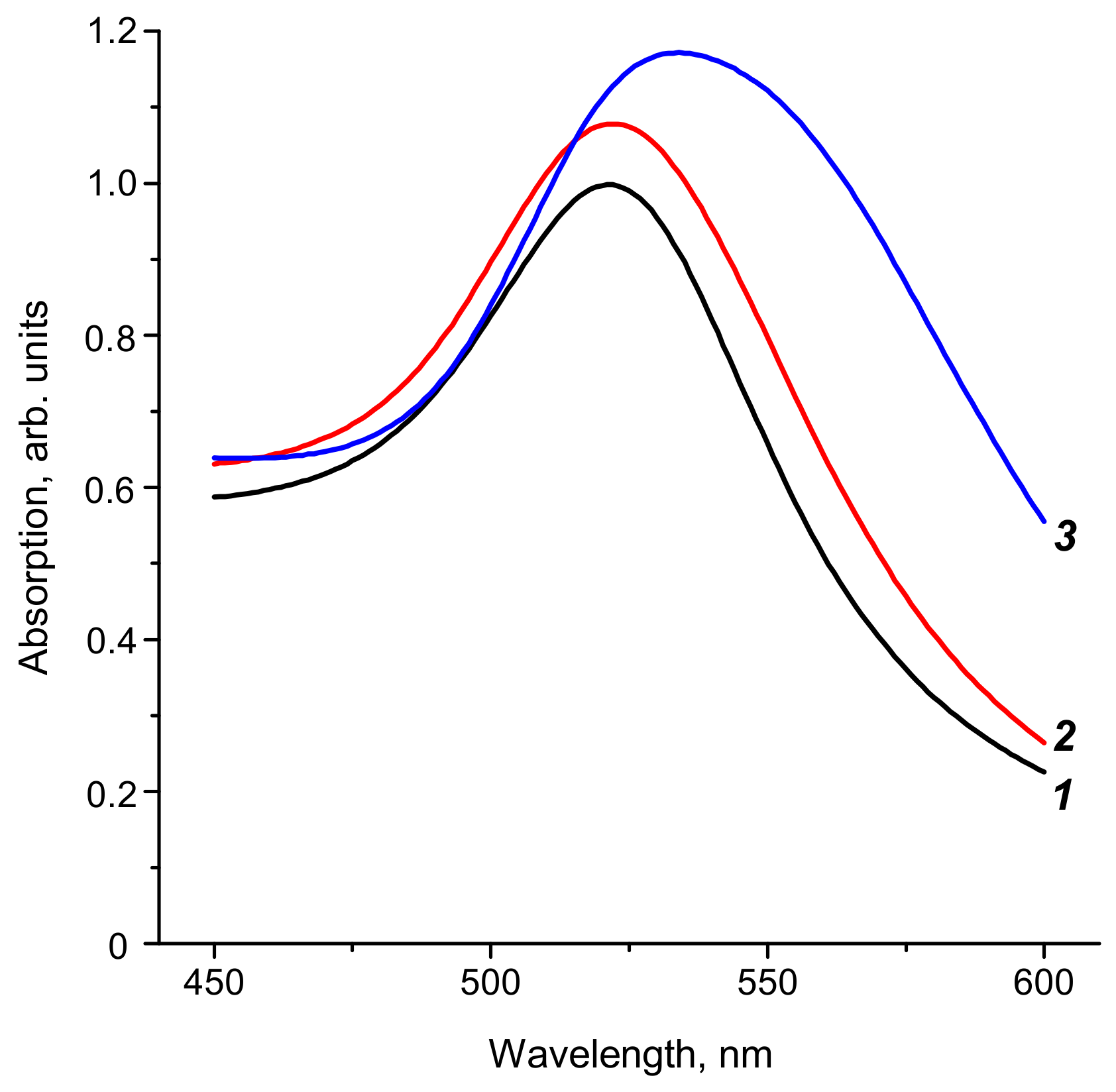
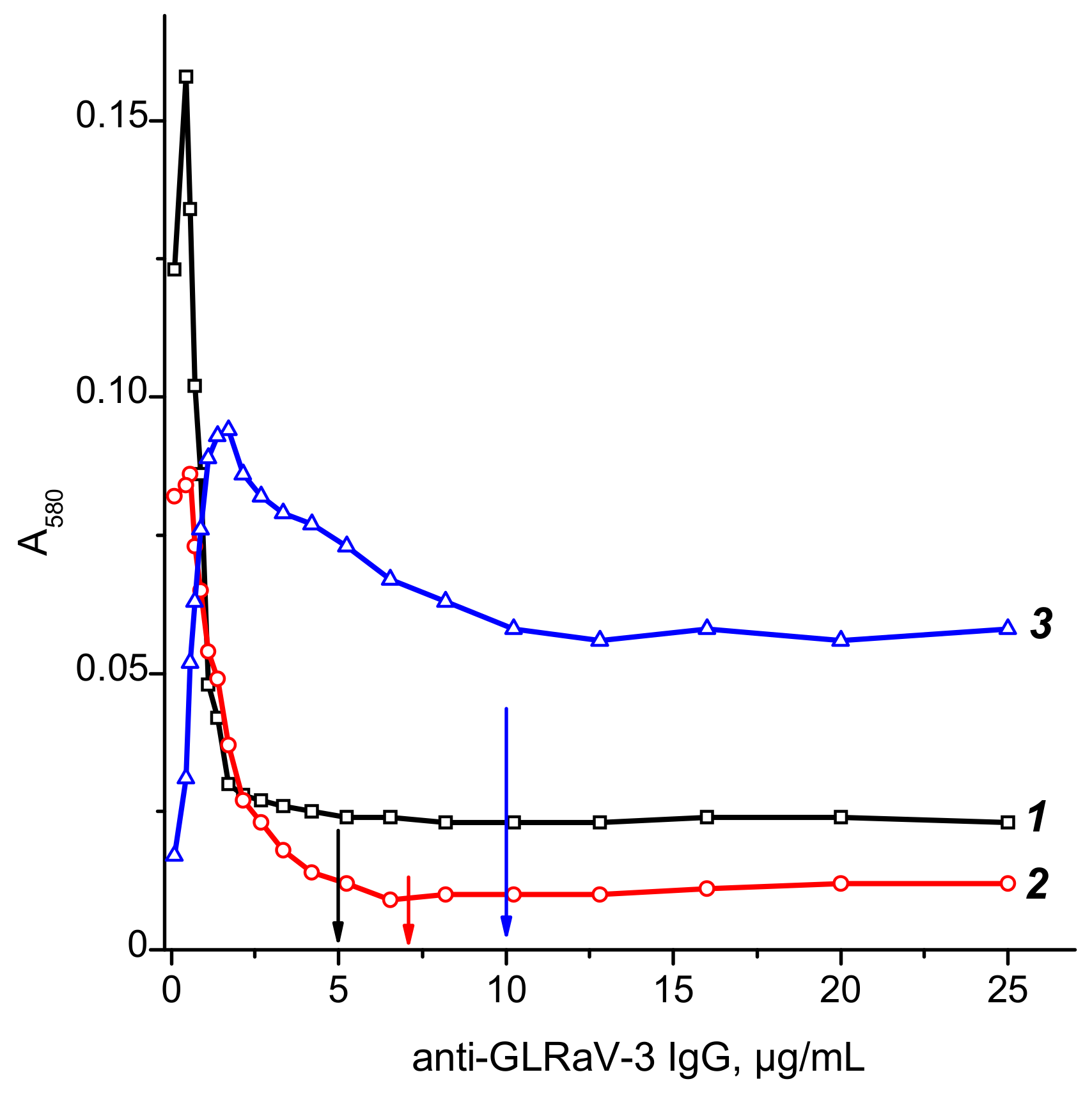
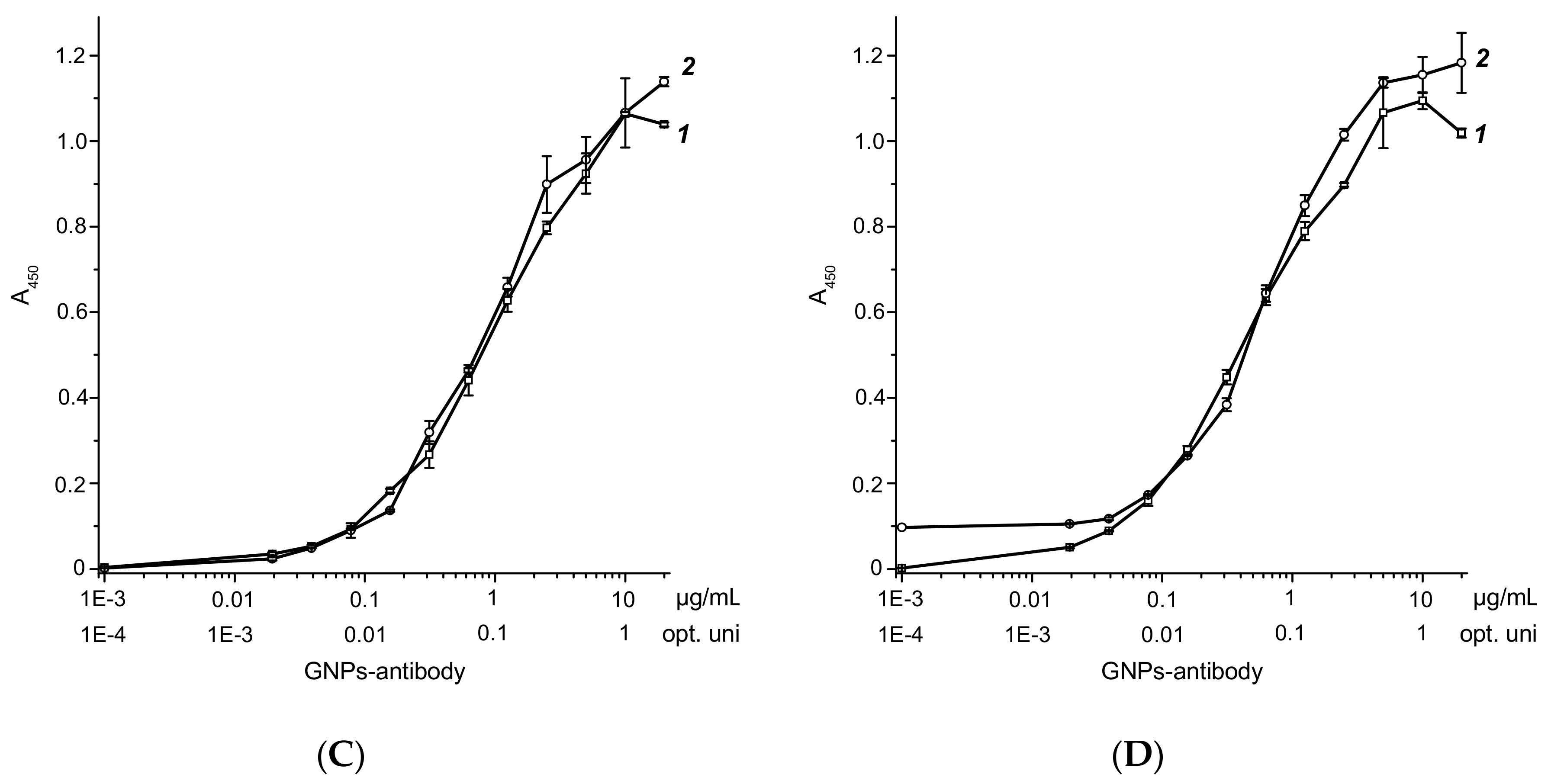
2.1. Synthesis and Characterization of Reagents for ICA
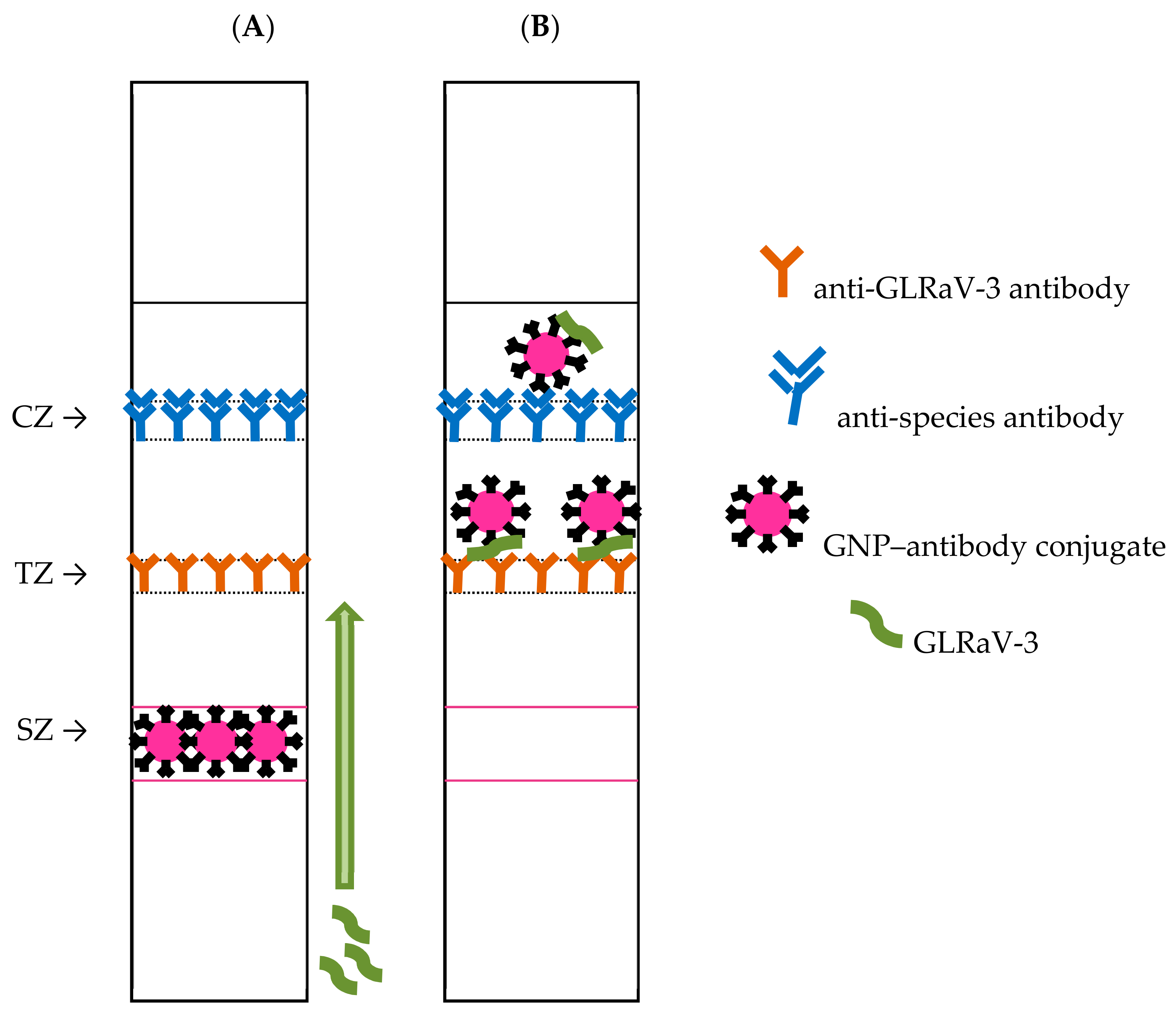
2.2. Preparation of Test Strips
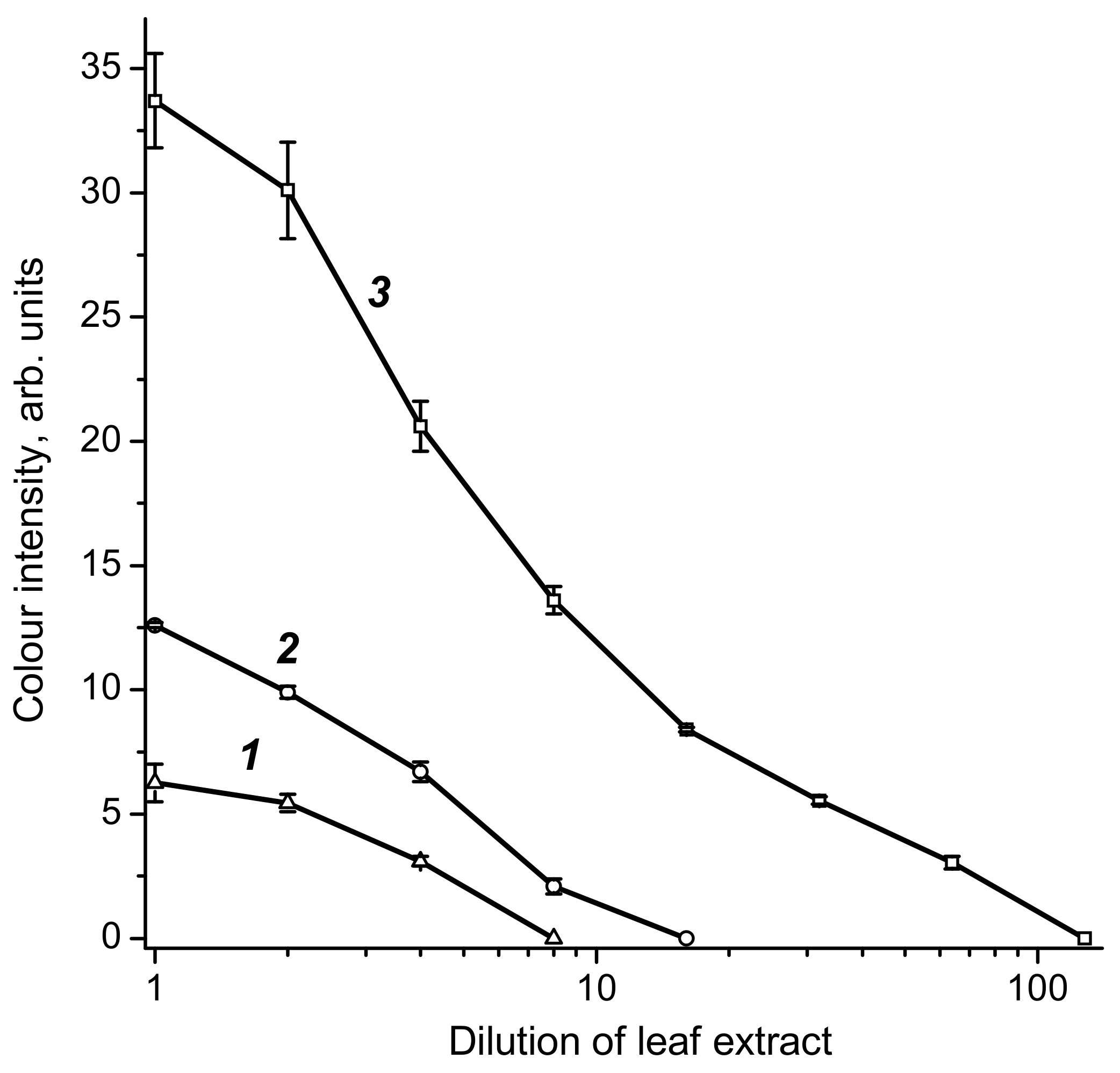
2.3. Preparation of Pooled Positive Sample of Grape Leaf Extracts
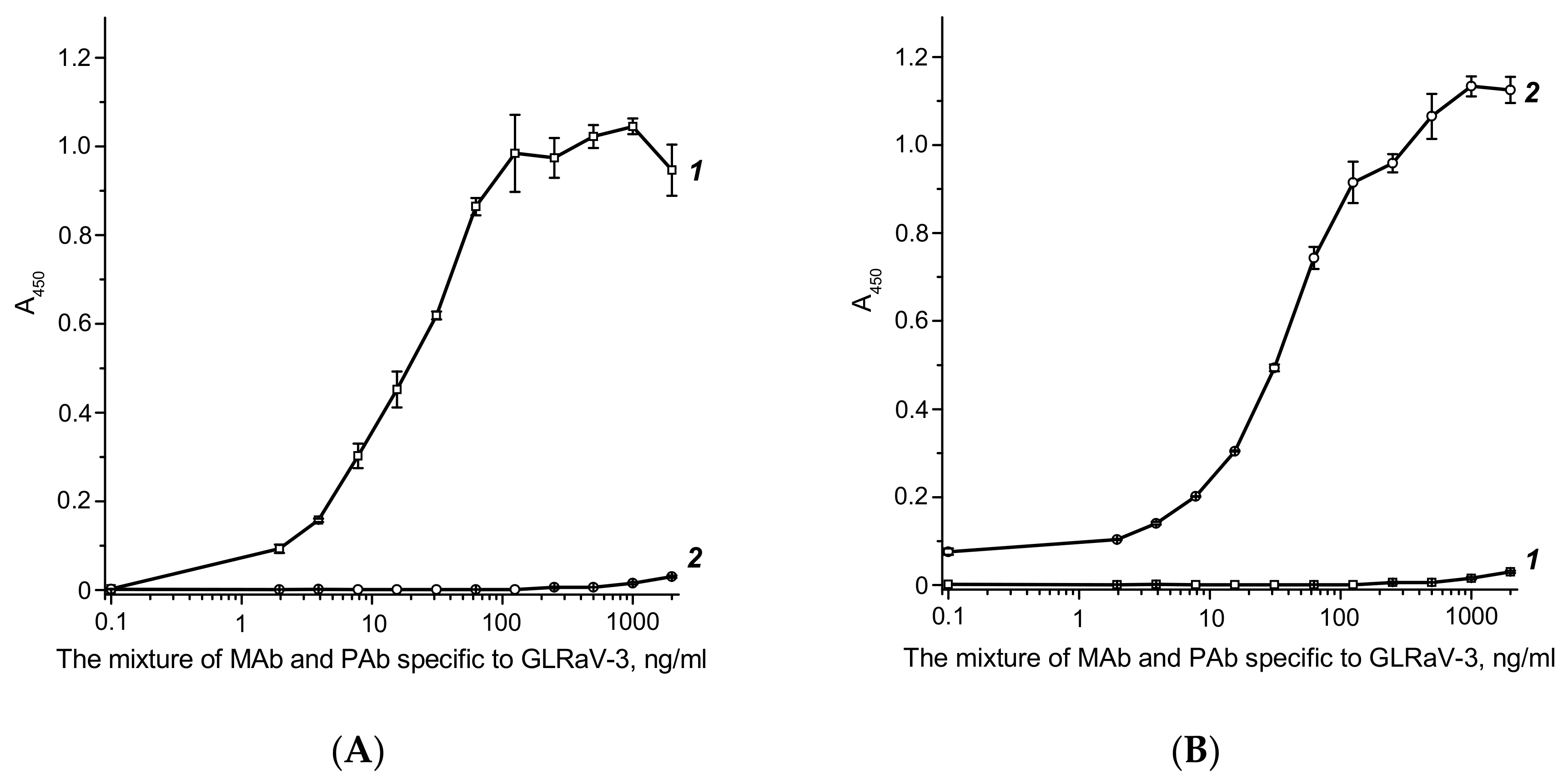
2.4. Characterization of the Developed Test System
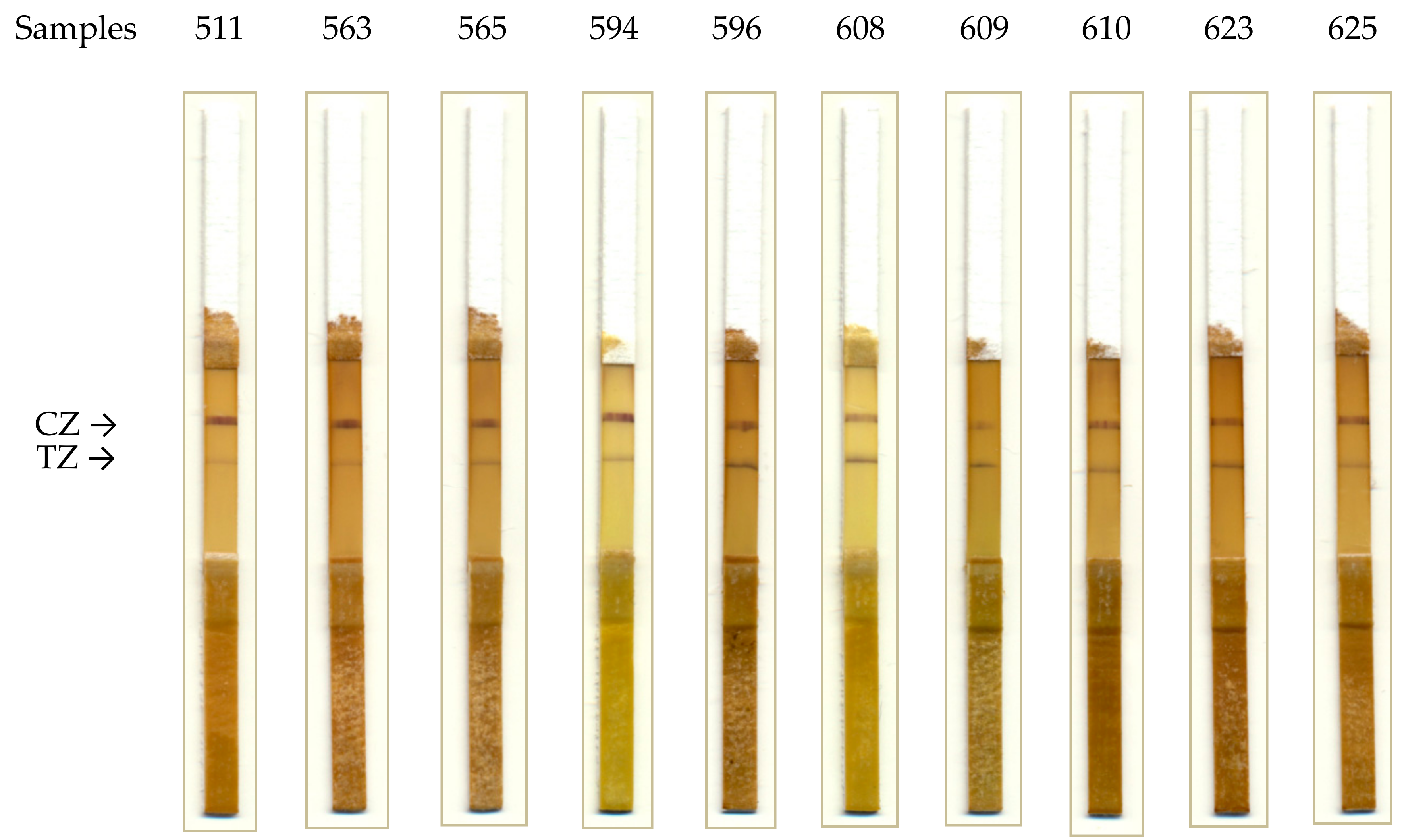
2.5. Approbation of the Developed Test System
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Synthesis of GNPs
4.3. Transmission Electron Microscopy
4.4. Obtaining Flocculation Curves
4.5. Synthesis of Gnps–Antibody Conjugates
4.6. Functional Characterization of The Conjugate Gnps–Antibody
4.7. Preparation of Test Strips
4.8. Preparation of Grapevine Leaves Extracts
4.9. Lateral Flow Immunoassay
4.10. Reference ELISA Assay
4.11. PCR Analysis
4.12. Analysis of Approbation Data for the Developed Test Strips
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maliogka, V.I.; Martelli, G.P.; Fuchs, M.; Katis, N.I. Control of viruses infecting grapevine. In Advance in Virus Research, Control of Plant Virus Diseases Vegetatively-Propagated Crops; Loebenstein, G., Katis, N.I., Eds.; Academic Press: Boston, MA, USA, 2015; Volume 91, pp. 175–227. [Google Scholar]
- Meng, B.; Martelli, G.P.; Golino, D.A.; Fuchs, M. (Eds.) Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer: Heidelberg, Germany, 2017; ISBN 978-3-319-57706-7. [Google Scholar]
- Perrone, I.; Chitarra, W.; Boccacci, P.; Gambio, G. Grapevine-virus-environment interactions: An intriguing puzzle to solve. New Phytol. 2017, 213, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Maree, H.J.; Almeida, R.P.P.; Bester, R.; Chooi, K.M.; Cohen, D.; Dolja, V.V.; Fuchs, M.F.; Golino, D.A.; Jooste, A.E.C.; Martelli, G.P.; et al. Grapevine leafroll-associated virus 3. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, J.T.; Maree, H.J.; Gouveia, P.; Naidu, R.A. Grapevine leafroll-associated virus 3. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Meng, B., Martelli, G.P., Golino, D.A., Fuchs, M., Eds.; Springer: Heidelberg, Germany, 2017; pp. 167–196. ISBN 978-3-319-57706-7. [Google Scholar]
- Rayapati, A.N.; O’Neal, S.; Walsh, D. Grapevine leafroll disease. WSU Ext. Bull. 2008, EB2027E, 1–17. [Google Scholar]
- Almeida, R.P.P.; Daane, K.M.; Bell, V.A.; Blaisdell, G.K.; Cooper, M.L.; Herrbach, E.; Pietersen, G. Ecology and management of grapevine leafroll disease. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martelli, G.P. Directory of virus and virus-like diseases of the grapevine and their agents. J. Plant. Pathol. 2014, 96. [Google Scholar] [CrossRef]
- Alabi, O.J.; Casassa, L.F.; Gutha, L.R.; Larsen, R.C.; Henick-Kling, T.; Harbertson, J.F.; Naidu, R.A. Impacts of grapevine leafroll disease on fruit yield and grape and wine chemistry in a wine grape (Vitis vinifera L.) cultivar. PLoS ONE 2016, 11, e0149666. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P. Grapevine-infecting viruses. Eur. J. Plant. Pathol. 2014, 96, 7–8. [Google Scholar]
- Naidu, R.; Rowhani, A.; Fuchs, M.; Golino, D.; Martelli, G.P. Grapevine leafroll: A complex viral disease affecting a high-value fruit crop. Plant Dis. 2014, 98, 1172–1185. [Google Scholar] [CrossRef]
- Naidu, R.A.; Maree, H.J.; Burger, J.T. Grapevine leafroll disease and associated viruses: A unique pathosystem. Annu. Rev. Phytopathol. 2015, 53, 613–634. [Google Scholar] [CrossRef] [PubMed]
- EPPO Standard PP 2/23 (1) Grapevine. EPPO Bull. 2002, 32, 371–392. Available online: https://onlinelibrary.wiley.com/doi/epdf/10.1046/j.1365-2338.2002.00571.x (accessed on 21 October 2018).
- EPPO Standard PM 4/8 Pathogen-Tested Material of Grapevine Varieties and Rootstocks. EPPO Bull. 2008, 38, 422–429. Available online: https://onlinelibrary.wiley.com/doi/epdf/10.1111/j.1365-2338.2008.01258.x (accessed on 21 October 2018). [CrossRef]
- Rowhani, A.; Uyemoto, J.K.; Golino, D.A. A comparison between serological and biological assays in detecting grapevine leafroll associated viruses. Plant Dis. 1997, 81, 799–801. [Google Scholar] [CrossRef]
- Martelli, G.P.; Uyemoto, J.K. Plant virus diseases: Fruit trees and grapevine. In Encyclopedia of Virology, 3rd ed.; Mahy, B.W.J., Van Regenmortel, M.H.V., Eds.; Academic Press: San Diego, CA, USA, 2008; Volume 4, pp. 201–207. ISBN 978-0-12-374410-4. [Google Scholar]
- Gambino, G.; Angelini, E.; Gribaudo, I. Field assessment and diagnostic methods for detection of grapevine viruses. In Methodologies and Results in Grapevine Research; Springer: Berlin, Germany, 2010; pp. 211–228. [Google Scholar]
- Fang, Y.; Ramasamy, R.P. Current and prospective methods for plant disease detection. Biosensors 2015, 5, 537–561. [Google Scholar] [CrossRef] [PubMed]
- Gambino, G.; Gribaudo, I. Simultaneous detection of nine grapevine viruses by multiplex reverse transcription-polymerase chain reaction with coamplification of a plant RNA as internal control. Phytopathology 2006, 96, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Poojari, S.; Alabi, O.J.; Okubara, P.A.; Naidu, R.A. SYBR® Green-based real-time quantitative reverse-transcription PCR for detection and discrimination of grapevine viruses. J. Virol. Methods 2016, 235, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Aloisio, M.; Morelli, M.; Elicio, V.; Saldarelli, P.; Ura, B.; Bortot, B.; Severini, G.M.; Minafra, A. Detection of four regulated grapevine viruses in a qualitative, single tube real-time PCR with melting curve analysis. J. Virol. Methods 2018, 257, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Rubinson, E.; Galiakparov, N.; Radian, S.; Sela, I.; Tanne, E.; Gafny, R. Serological detection of grapevine virus a using antiserum to a nonstructural protein, the putative movement protein. Phytopathology 1997, 87, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Abdullahi, I.; Rott, M. Microarray immunoassay for the detection of grapevine and tree fruit viruses. J. Virol. Methods 2009, 160, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Koolivand, D.; Bashir, N.S.; Behjatnia, S.A.; Joozani, R.J. Production of polyclonal antibody against grapevine fanleaf virus movement protein expressed in Escherichia coli. Plant Pathol. J. 2016, 32, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Olmos, A.; Bertolini, E.; Ruiz-García, A.B.; Martínez, C.; Peiró, R.; Vidal, E. Modeling the accuracy of three detection methods of grapevine leafroll-associated virus 3 during the dormant period using a bayesian approach. Phytopathology 2016, 106, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Chiriacò, M.S.; Luvisi, A.; Primiceri, E.; Sabella, E.; De Bellis, L.; Maruccio, G. Development of a lab-on-a-chip method for rapid assay of Xylella fastidiosa subsp. pauca strain CoDiRO. Sci. Rep. 2018, 8, 7376. [Google Scholar] [CrossRef] [PubMed]
- Nezhad, A.S. Future of portable devices for plant pathogen diagnosis. Lab Chip 2014, 14, 2887–2904. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.R; Fuchs, M.; Fischer, K.F.; Perry, K.L. Macroarray detection of grapevine leafroll-associated viruses. J. Virol. Methods 2012, 183, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.L.; Aguilar, Z.P.; Xu, H.; Lai, W.; Xiong, Y. Membrane-based lateral flow immunochromatographic strip with nanoparticles as reporters for detection: A review. Biosens. Bioelectron. 2016, 75, 166–180. [Google Scholar] [CrossRef] [PubMed]
- De Boer, S.H.; Lopez, M.M. New grower-friendly methods for plant pathogen monitoring. Annu. Rev. Phytopathol. 2012, 50, 197–218. [Google Scholar] [CrossRef] [PubMed]
- Dzantiev, B.B.; Byzova, N.A.; Urusov, A.E.; Zherdev, A.V. Immunochromatographic methods in food analysis. TrAC Trends Anal. Chem. 2014, 55, 81–93. [Google Scholar] [CrossRef]
- Liebenberg, A.; Freeborough, M.-J.; Visser, C.J.; Bellstedt, D.U.; Burger, J.T. Genetic variability within the coat protein gene of grapevine fanleaf virus isolates from South Africa and the evaluation of RT-PCR, DAS-ELISA and ImmunoStrips as virus diagnostic assays. Virus Res. 2009, 142, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Huseynova, I.; Sultanova, N.; Mammadov, A.; Kosayeva, N.; Khanishova, M.; Aliyev, J. Detection of grapevine leafroll-assosiated virus type 3 (GLRaV-3) in Azerbaijan and study of some histopathological changes in leaves of infected plants. In Proceedings of the 18th Conference of the International Council for the Study of Virus and Virus-like Diseases of the Grapevine, Ankara, Turkey, 7–11 September 2015; pp. 48–49. [Google Scholar]
- Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. Correlation between the composition of multivalent antibody conjugates with colloidal gold nanoparticles and their affinity. J. Immunol. Methods 2010, 357, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. Factors influencing the detection limit of the lateral-flow sandwich immunoassay: A case study with potato virus X. Anal. Bioanal. Chem. 2012, 403, 1595–1605. [Google Scholar] [CrossRef] [PubMed]
- Makhsin, S.R.; Razak, K.A.; Noordin, R.; Zakaria, N.D.; Chun, T.S. The effects of size and synthesis methods of gold nanoparticle-conjugated M alpha HIgG4 for use in an immunochromatographic strip test to detect brugian filariasis. Nanotechnology 2012, 23, 495719. [Google Scholar] [CrossRef] [PubMed]
- Dykman, L.; Khlebtsov, N. Gold nanoparticles in biomedical applications: Recent advances and perspectives. Chem. Soc. Rev. 2012, 41, 2256–2282. [Google Scholar] [CrossRef] [PubMed]
- Wild, D. (Ed.) The Immunoassay Handbook: Theory and Applications of Ligand Binding, ELISA and Related Techniques, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9780080970370. [Google Scholar]
- Hermanson, G.T. Bioconjugate Techniques, 3rd ed.; Academic Press: Boston, MA, USA, 2013; ISBN 9780123822390. [Google Scholar]
- Byzova, N.A.; Safenkova, I.V.; Slutskaya, E.S.; Zherdev, A.V.; Dzantiev, B.B. Less is more: A comparison of antibody—Gold nanoparticle conjugates of different ratios. Bioconj. Chem. 2017, 28, 2737–2746. [Google Scholar] [CrossRef] [PubMed]
- Bioreba Product Information: DAS-ELISA Grapevine Leafroll-Associated Virus 3 (GLRaV-3). Available online: www.bioreba.ch/popup.php?docFile=http://www.bioreba.ch/files/Product_Info/ELISA_Reagents/GLRaV_3_DAS_ELISA.pdf (accessed on 21 October 2018).
- Byzova, N.A.; Safenkova, I.V.; Chirkov, S.N.; Avdienko, V.G.; Guseva, A.N.; Mitrofanova, I.V.; Zherdev, A.V.; Dzantiev, B.B.; Atabekov, J.G. Interaction of plum pox virus with colloidal gold labeled antibodies and development of an immunochromatographic system for detection of the virus. Biochemistry (Moscow) 2010, 75, 1393–1403. [Google Scholar] [CrossRef] [PubMed]
- Byzova, N.A.; Smirnova, N.I.; Zherdev, A.V.; Eremin, S.A.; Shanin, I.A.; Lei, H.-T.; Sun, Y.; Dzantiev, B.B. Rapid immunochromatographic assay for ofloxacin in animal original foodstuffs using antisera labeled by colloidal gold. Talanta 2014, 119, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.C.; Tse, H.Y. Lateral Flow Immunoassay; Humana Press: New York, NY, USA, 2009; Volume 1318, pp. 127–137. [Google Scholar]
- Ling, K.S.; Zhu, H.Y.; Jiang, Z.Y.; Gonsalves, D. Effective application of DAS-ELISA for detection of grapevine leafroll associated closterovirus-3 using a polyclonal antiserum developed from recombinant coat protein. Eur. J. Plant Pathol. 2000, 106, 301–309. [Google Scholar] [CrossRef]
- Frens, G. Controlled nucleation for the regulation on particle size in monodisperse gold suspension. Nat. Phys. Sci. 1973, 241, 20–22. [Google Scholar] [CrossRef]
- Rott, M.E.; Jelkmann, W. Characterization and detection of several filamentous viruses of cherry, adaptation of an alternative cloning method (DOP-PCR), and modification of an RNA extraction protocol. Eur. J. Plant Pathol. 2001, 107, 411–420. [Google Scholar] [CrossRef]
- Gribaudo, I.; Gambino, G.; Cuozzo, D.; Mannini, F. Attempts to eliminate Grapevine rupestris stem pitting-associated virus from grapevine clones. J. Plant Pathol. 2006, 88, 293–298. [Google Scholar]
- Minafra, A.; Hadidi, A. Sensitive detection of grapevine viruses A, B, or leafroll-associated III from viruliferous mealybugs and infected tissue by cDNA amplification. J. Virol. Methods 1994, 47, 175–187. [Google Scholar] [CrossRef]
- Fawcett, T. An introduction to ROC analysis. Pattern Recognit. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ICA Testing | Confirmation by ELISA | Confirmation by PCR |
|---|---|---|
| Positive—14 | Positive—14 Negative—0 | Positive—13 Negative—1 |
| Negative—53 | Positive—4 Negative—49 | Positive—4 Negative—49 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byzova, N.A.; Vinogradova, S.V.; Porotikova, E.V.; Terekhova, U.D.; Zherdev, A.V.; Dzantiev, B.B. Lateral Flow Immunoassay for Rapid Detection of Grapevine Leafroll-Associated Virus. Biosensors 2018, 8, 111. https://doi.org/10.3390/bios8040111
Byzova NA, Vinogradova SV, Porotikova EV, Terekhova UD, Zherdev AV, Dzantiev BB. Lateral Flow Immunoassay for Rapid Detection of Grapevine Leafroll-Associated Virus. Biosensors. 2018; 8(4):111. https://doi.org/10.3390/bios8040111
Chicago/Turabian StyleByzova, Nadezhda A., Svetlana V. Vinogradova, Elena V. Porotikova, Uliana D. Terekhova, Anatoly V. Zherdev, and Boris B. Dzantiev. 2018. "Lateral Flow Immunoassay for Rapid Detection of Grapevine Leafroll-Associated Virus" Biosensors 8, no. 4: 111. https://doi.org/10.3390/bios8040111
APA StyleByzova, N. A., Vinogradova, S. V., Porotikova, E. V., Terekhova, U. D., Zherdev, A. V., & Dzantiev, B. B. (2018). Lateral Flow Immunoassay for Rapid Detection of Grapevine Leafroll-Associated Virus. Biosensors, 8(4), 111. https://doi.org/10.3390/bios8040111