
Physical Training Increases Erythroferrone Levels in Men



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Physical Activity Assessment
2.3. Blood Analysis
2.4. Statistical Analysis
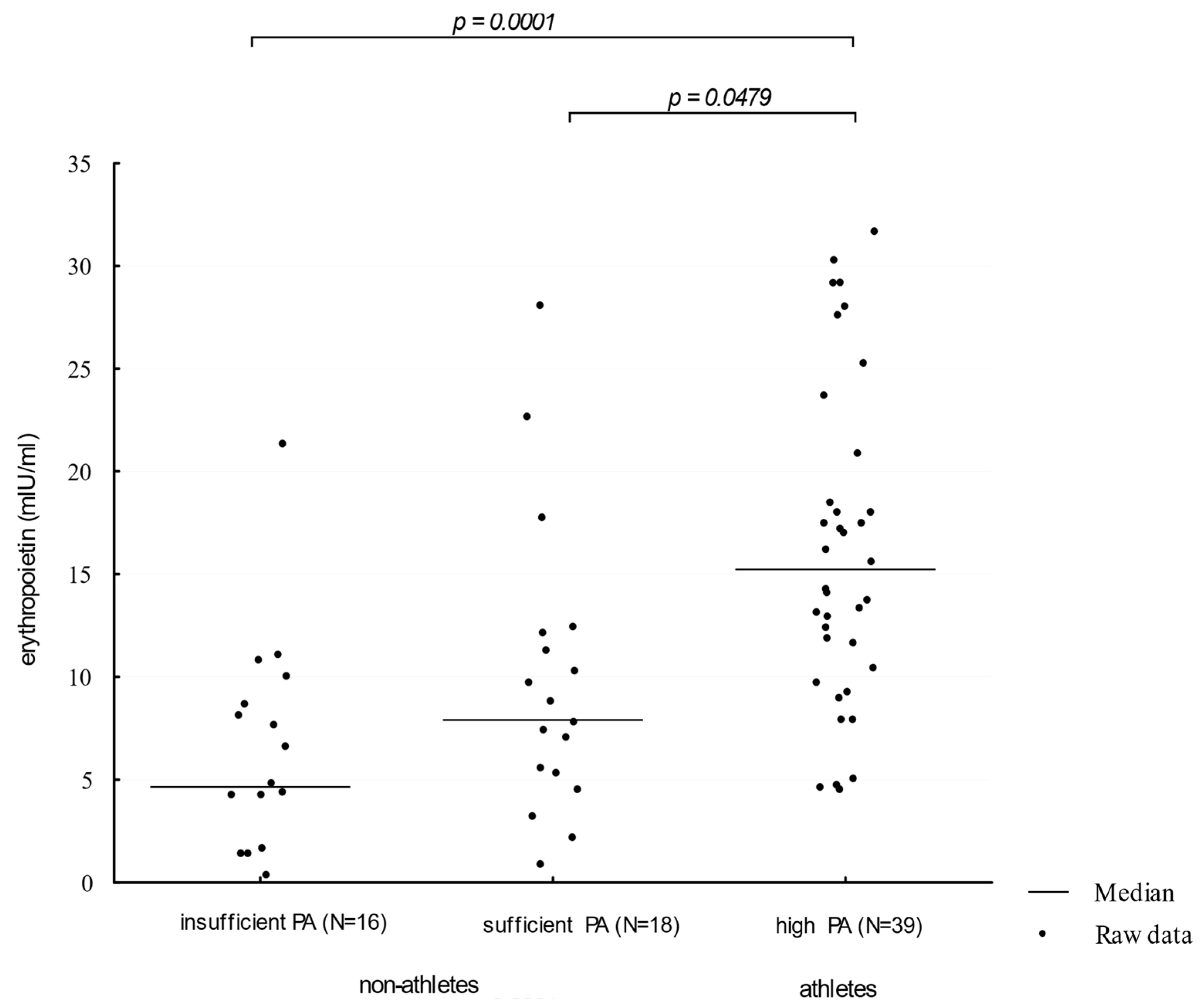
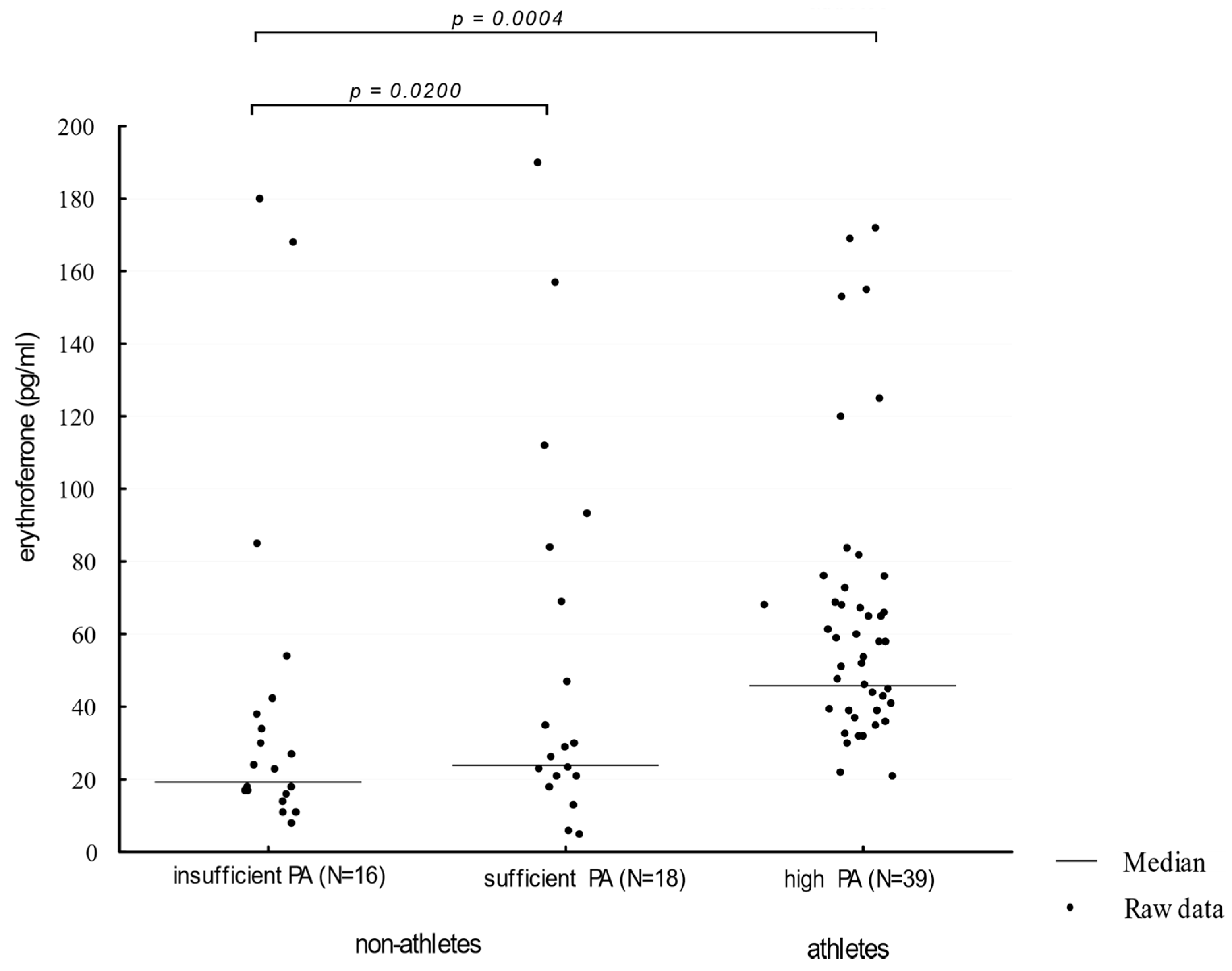
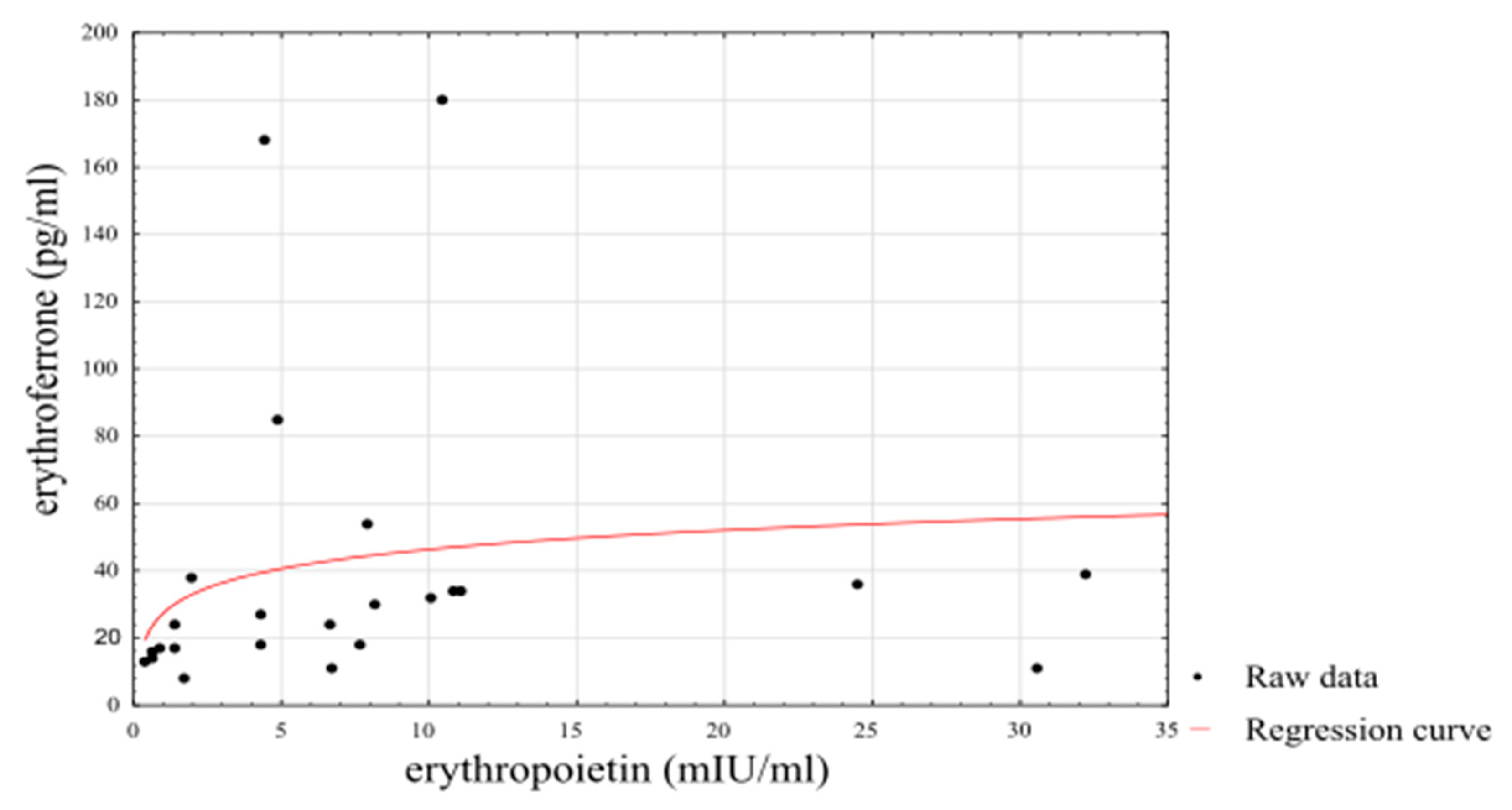
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lombardi, G.; Colombini, A.; Lanteri, P.; Banfi, G. Reticulocytes in sports medicine: An update. Adv. Clin. Chem. 2013, 59, 125–153. [Google Scholar]
- Rybinska, I.; Cairo, G. Mutual Cross Talk Between Iron Homeostasis and Erythropoiesis. Vitam. Horm. 2017, 105, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Park, W.; Lim, K. Living High-Training Low for 21 Days Enhances Exercise Economy, Hemodynamic Function, and Exercise Performance of Competitive Runners. J. Sports Sci. Med. 2019, 18, 427–437. [Google Scholar]
- Montero, D.; Breenfeldt-Andersen, A.; Oberholzer, L.; Haider, T.; Goetze, J.P.; Meinild-Lundby, A.-K.; Lundby, C. Erythropoiesis with endurance training: Dynamics and mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R894–R902. [Google Scholar] [CrossRef] [PubMed]
- Lobigs, L.M.; Sharpe, K.; Garvican-Lewis, L.A.; Gore, C.J.; Peeling, P.; Dawson, B.; Schumacher, Y.O. The athlete’s hematological response to hypoxia: A meta-analysis on the influence of altitude exposure on key biomarkers of erythropoiesis. Am. J. Hematol. 2018, 93, 74–83. [Google Scholar] [CrossRef]
- Campbell, J.P.; Turner, J.E. Debunking the myth of exercise-induced immune suppression: Redefining the impact of exercise on immunological health across the lifespan. Front. Immunol. 2018, 9, 648. [Google Scholar] [CrossRef]
- Domínguez, R.; Sánchez-Oliver, A.J.; Mata-Ordoñez, F.; Feria-Madueño, A.; Grimaldi-Puyana, M.; López-Samanes, Á.; Pérez-López, A. Effects of an Acute Exercise Bout on Serum Hepcidin Levels. Nutrients 2018, 10, 209. [Google Scholar] [CrossRef] [Green Version]
- Scheffer, D.D.L.; Latini, A. Exercise-induced immune system response: Anti-inflammatory status on peripheral and central organs. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165823. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Muraoka, I. Exercise-Induced Oxidative Stress and the Effects of Antioxidant Intake from a Physiological Viewpoint. Antioxidants 2018, 7, 119. [Google Scholar] [CrossRef] [Green Version]
- Yusof, A.; Leithauser, R.M.; Roth, H.J.; Finkernagel, H.; Wilson, M.T.; Beneke, R. Exercise-induced hemolysis is caused by protein modification and most evident during the early phase of an ultraendurance race. J. Appl. Physiol. 2007, 102, 582–586. [Google Scholar] [CrossRef] [Green Version]
- Płoszczyca, K.; Langfort, J.; Czuba, M. The Effects of Altitude Training on Erythropoietic esponse and Hematological Variables in Adult Athletes: A Narrative Review. Front. Physiol. 2018, 9, 375. [Google Scholar] [CrossRef] [Green Version]
- Kautz, L.; Jung, G.; Valore, E.V.; Rivella, S.; Nemeth, E.; Ganz, T. Identification of erythroferrone as an erythroid regulator of iron metabolism. Nat. Genet. 2014, 46, 678–684. [Google Scholar] [CrossRef] [Green Version]
- Ramirez Cuevas, K.; Schobinger, C.; Gottardo, E.; Voss, S.C.; Kuuranne, T.; Tissot, J.-D.; Favrat, B.; Townsend, N.; Leuenberger, N. Erythroferrone as a sensitive biomarker to detect stimulation of erythropoiesis. Drug Test. Anal. 2019, 12, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T. Erythropoietic regulators of iron metabolism. Free Radic. Biol. Med. 2019, 133, 69–74. [Google Scholar] [CrossRef]
- Bergier, J.; Kapka-Skrzypczak, L.; Biliński, P.; Paprzycki, P.; Wojtyla, A. Physical activity of Polish adolescents and young adults according to IPAQ: A population based study. Ann. Agric. Environ. Med. 2012, 19, 109–115. [Google Scholar] [PubMed]
- Ganz, T.; Jung, G.; Naeim, A.; Ginzburg, Y.Z.; Pakbaz, Z.; Walter, P.B.; Kautz, L.; Nemeth, E. Immunoassay for human serum erythroferrone. Blood 2017, 130, 1243–1246. [Google Scholar] [CrossRef] [Green Version]
- Kautz, L.; Jung, G.; Du, X.; Gabayan, V.; Chapman, J.; Nasoff, M.; Nemeth, E.; Ganz, T. Erythroferrone contributes to hepcidin suppression and ironoverload in a mouse model of β-thalassemia. Blood 2015, 126, 2031–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kautz, L.; Jung, G.; Nemeth, E.; Ganz, T. Erythroferrone contributes to recovery from anemia of inflammation. Blood 2014, 124, 2569–2574. [Google Scholar] [CrossRef] [Green Version]
- Lawen, A. Is erythroferrone finally the long sought-after systemic erythroid regulator of iron? World J. Biol. Chem. 2015, 6, 78–82. [Google Scholar] [CrossRef]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A Red Carpet for Iron Metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [Green Version]
- Jelkmann, W. Erythropoietin. Front. Horm. Res. 2016, 47, 115–127. [Google Scholar]
- Lamon, S.; Russell, A.P. The role and regulation of erythropoietin (EPO) and its receptor in skeletal muscle: How much do we really know? Front. Physiol. 2013, 4, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimáková, P.; Solár, P.; Solárová, Z.; Komel, R.; Debeljak, N. Erythropoietin and Its Angiogenic Activity. Int. J. Mol. Sci. 2017, 18, 1519. [Google Scholar] [CrossRef]
- Kasperska, A.; Zembron-Lacny, A. The effect of intermittent hypoxic exposure on erythropoietic response and hematological variables in elite athletes. Physiol. Res. 2020, 69, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Rundqvist, H.; Rullman, E.; Sundberg, C.J.; Fischer, H.; Eisleitner, K.; Ståhlberg, M.; Sundblad, P.; Jansson, E.; Gustafsson, T. Activation of the erythropoietin receptor in human skeletal muscle. Eur. J. Endocrinol. 2009, 161, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Edgett, B.A.; Fraquharson, L.; Romanova, N.; Brunt, K.R.; Gurd, B.J.; Simpson, J.A. Regulation of erythropoietin expression in response to exercise in mouse and human skeletal muscle. Fed. Am. Soc. Exp. Biol. J. 2017, 17, lb748. [Google Scholar]
- Nijholt, K.T.; Meems, L.M.G.; Ruifrok, W.P.T.; Maass, A.H.; Yurista, S.R.; Pavez-Giani, M.G.; Mahmoud, B.; Wolters, A.H.G.; van Veldhuisen, D.J.; van Gilst, W.H.; et al. The erythropoietin receptor expressed in skeletal muscle is essential for mitochondrial biogenesis and physiological exercise. Pflugers Arch. Eur. J. Physiol. 2021, 473, 1301–1313. [Google Scholar] [CrossRef] [PubMed]
- Luccia, T.P.B.; Natali, J.E.S.; Moreira, A.; Chaui-Berlinck, J.G.; Bicudo, J.E.P.W. Bouts of exercise elicit discordant testosterone:cortisol ratios in runners and non-runners. Arch. Endocrinol. Metab. 2018, 62, 325–331. [Google Scholar]
- Hayes, L.D.; Grace, F.M.; Baker, J.S.; Sculthorpe, N. Exercise-induced responses in salivary testosterone, cortisol, and their ratios in men: A meta-analysis. Sports Med. 2015, 45, 713–726. [Google Scholar] [CrossRef] [Green Version]
- Hayes, L.D.; Herbert, P.; Sculthorpe, N.F.; Grace, F.M. Exercise training improves free testosterone in lifelong sedentary aging men. Endocr. Connect. 2017, 6, 306–310. [Google Scholar] [CrossRef] [Green Version]
- Molinari, P.F. Erythropoietic mechanism of androgens: A critical review and clinical implications. Haematologica 1982, 67, 442–460. [Google Scholar] [PubMed]
- Kawabata, H. Transferrin and transferrin receptors update. Free Radic. Biol. Med. 2018, 133, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Peeling, P.; Sim, M.; Badenhorst, C.E.; Dawson, B.; Govus, A.D.; Abbiss, C.R.; Swinkels, D.W.; Trinder, D. Iron status and the acute post-exercise hepcidin response in athletes. PLoS ONE 2014, 9, e93002. [Google Scholar]
- Kanter, M. High-Quality Carbohydrates and Physical Performance: Expert Panel Report. Nutr. Today 2018, 53, 35–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variable | Athletes | Non-Athletes | p | η2 |
|---|---|---|---|---|
| N = 39 | N = 34 | |||
| Me (Min–Max) | Me (Min–Max) | |||
| Age (years) | 21.00 (18.00–32.00) | 21.50 (18.00–29.00) | 0.5321 | 0.0031 |
| BMI (kg/m2) | 23.12 (17.28–27.15) | 23.61 (17.94–28.12) | 0.2431 | 0.0000 |
| Physical activity (MET × min/week) | 5120.00 (3100.00–6300.00) | 1230.00 (340.00–2100.00) | <0.0001 | 0.5706 |
| Years of training | 9.00 (4.00–14.00) | N/A | N/A | N/A |
| Serum EPO levels (mIU/mL) | 12.35 (0.90–32.25) | 5.68 (0.38–32.15) | 0.0001 | 0.1612 |
| Serum ERFE levels (pg/mL) | 40.00 (5.00–190.00) | 25.5 (8.00–180.00) | 0.0042 | 0.1861 |
| Serum hepcidin levels (pg/mL) | 8.43 (5.81–12.54) | 8.21 (5.92–12.32) | 0.5431 | 0.0001 |
| Serum ferritin levels (ng/mL) | 57.36 (20.58–158.32) | 45.28 (21.52–162.45) | 0.3412 | 0.0013 |
| Serum sTfR levels (µg/mL) | 1.28 (0.81–3.43) | 1.83 (1.47–2.78) | 0.0002 | 0.1014 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziembowska, I.; Wójcik, M.; Bukowski, J.; Żekanowska, E. Physical Training Increases Erythroferrone Levels in Men. Biology 2021, 10, 1215. https://doi.org/10.3390/biology10111215
Dziembowska I, Wójcik M, Bukowski J, Żekanowska E. Physical Training Increases Erythroferrone Levels in Men. Biology. 2021; 10(11):1215. https://doi.org/10.3390/biology10111215
Chicago/Turabian StyleDziembowska, Inga, Małgorzata Wójcik, Jakub Bukowski, and Ewa Żekanowska. 2021. "Physical Training Increases Erythroferrone Levels in Men" Biology 10, no. 11: 1215. https://doi.org/10.3390/biology10111215
APA StyleDziembowska, I., Wójcik, M., Bukowski, J., & Żekanowska, E. (2021). Physical Training Increases Erythroferrone Levels in Men. Biology, 10(11), 1215. https://doi.org/10.3390/biology10111215