Surviving Starvation: Proteomic and Lipidomic Profiling of Nutrient Deprivation in the Smallest Known Free-Living Eukaryote

 ,
, 
Abstract
:
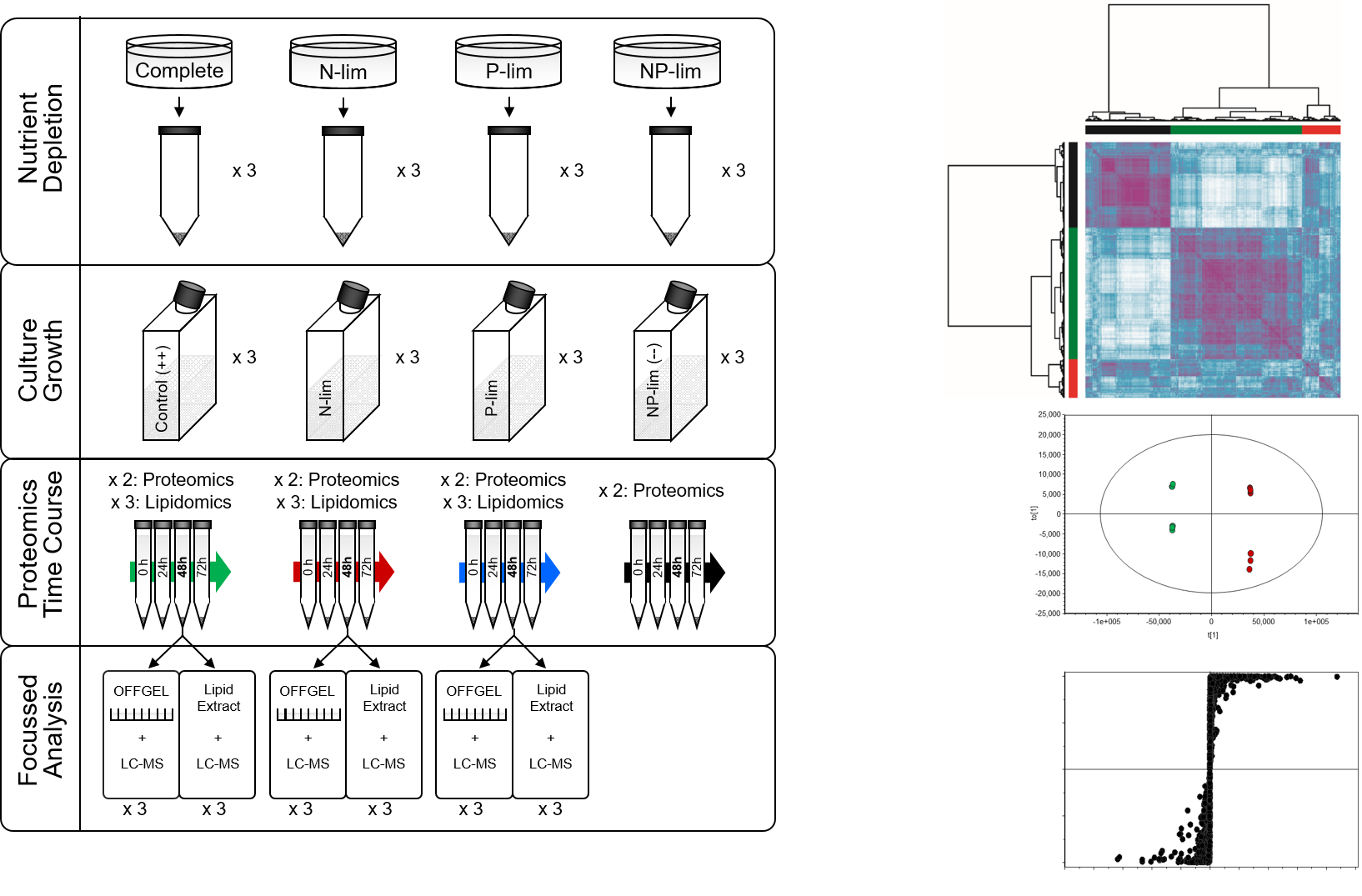{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Temporal Profiling of O. tauri Proteome under Nutrient Stress
2.2. Fractionation Provides In-Depth Characterisation of Differentially Expressed O. tauri Proteins Induced by Nutrient Depletion
2.3. Lipid Profiling of O. tauri under Nitrogen- and Phosphorus-Limiting Conditions
3. Materials and Methods
3.1. Culturing and Harvesting of O. tauri
3.2. In-Solution Trypsin Digestion
3.3. Fractionation of Peptides
3.4. Protein Identification and Data Analysis
3.5. Lipid Extraction
3.6. Analysis of Lipids and Data Processing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Parker, M.S.; Mock, T.; Armbrust, E.V. Genomic insights into marine microalgae. Ann. Rev. Genet. 2008, 42, 619–645. [Google Scholar] [CrossRef] [PubMed]
- Longhurst, A.; Sathyendranath, S.; Platt, T.; Caverhill, C. An estimate of global primary production in the ocean from satellite radiometer data. J. Plankton Res. 1995, 17, 1245–1271. [Google Scholar] [CrossRef] [Green Version]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courties, C.; Vaquer, A.; Troussellier, M.; Lautier, J.; Chretiennotdinet, M.J.; Neveux, J.; Machado, C.; Claustre, H. Smallest eukaryotic organism. Nature 1994, 370, 255. [Google Scholar] [CrossRef]
- Henderson, G.P.; Gan, L.; Jensen, G.J. 3-D ultrastructure of O. tauri: Electron cryotomography of an entire eukaryotic cell. PLoS ONE 2007, 2, e749. [Google Scholar] [CrossRef] [Green Version]
- Derelle, E.; Ferraz, C.; Rombauts, S.; Rouzé, P.; Worden, A.Z.; Robbens, S.; Partensky, F.; Degroeve, S.; Echeynié, S.; Cooke, R.; et al. Genome analysis of the smallest free-living eukaryote Ostreococcus tauri unveils many unique features. Proc. Natl. Acad. Sci. USA 2006, 103, 11647–11652. [Google Scholar] [CrossRef] [Green Version]
- Demir-Hilton, E.; Sudek, S.; Cuvelier, M.L.; Gentemann, C.L.; Zehr, J.P.; Worden, A.Z. Global distribution patterns of distinct clades of the photosynthetic picoeukaryote Ostreococcus. ISME J. 2011, 5, 1095–1107. [Google Scholar] [CrossRef]
- Ryther, J.H.; Dunstan, W.M. Nitrogen, phosphorus, and eutrophication in the coastal marine environment. Science 1971, 171, 1008–1013. [Google Scholar] [CrossRef] [Green Version]
- Malone, T.C.; Conley, D.J.; Fisher, T.R.; Glibert, P.M.; Harding, L.W.; Sellner, K.G. Scales of nutrient-limited phytoplankton productivity in Chesapeake Bay. Estuaries 1996, 19, 371–385. [Google Scholar] [CrossRef]
- Conley, D.J. Biogeochemical nutrient cycles and nutrient management strategies. Hydrobiologia 1999, 410, 87–96. [Google Scholar] [CrossRef]
- Delaney, M. Phosphorus accumulation in marine sediments and the oceanic phosphorus cycle. Glob. Biogeochem. Cycles 1998, 12, 563–572. [Google Scholar] [CrossRef]
- Tyrrell, T. The relative influences of nitrogen and phosphorus on oceanic primary production. Nature 1999, 400, 525–531. [Google Scholar] [CrossRef]
- Altabet, M.A.; Higginson, M.J.; Murray, D.W. The effect of millennial-scale changes in Arabian Sea denitrification on atmospheric CO2. Nature 2002, 415, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Redfield, A. On the proportions of organic derivatives in sea water and their relation to the composition of plankton. In James Johnstone Memorial Volume; Daniel, R.J., Ed.; University Press of Liverpool: Liverpool, UK, 1934; pp. 177–192. [Google Scholar]
- Sanudo-Wilhelmy, S.A.; Kustka, A.B.; Gobler, C.J.; Hutchins, D.A.; Yang, M.; Lwiza, K.; Burns, J.; Capone, D.G.; Raven, J.A.; Carpenter, E.J. Phosphorus limitation of nitrogen fixation by Trichodesmium in the central Atlantic Ocean. Nature 2001, 411, 66–69. [Google Scholar] [CrossRef]
- Wu, J.; Sunda, W.; Boyle, E.A.; Karl, D.M. Phosphate depletion in the western North Atlantic Ocean. Science 2000, 289, 759–762. [Google Scholar] [CrossRef] [Green Version]
- Conley, D.J.; Paerl, H.W.; Howarth, R.W.; Boesch, D.F.; Seitzinger, S.P.; Havens, K.E.; Lancelot, C.; Likens, G.E. Controlling eutrophication: Nitrogen and phosphorus. Science 2009, 323, 1014–1015. [Google Scholar] [CrossRef]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef]
- Liu, B.; Benning, C. Lipid metabolism in microalgae distinguishes itself. Curr. Opin. Biotechnol. 2013, 24, 300–309. [Google Scholar] [CrossRef]
- Gong, Y.; Guo, X.; Wan, X.; Liang, Z.; Jiang, M. Triacylglycerol accumulation and change in fatty acid content of four marine oleaginous microalgae under nutrient limitation and at different culture ages. J. Basic Microbiol. 2013, 53, 29–36. [Google Scholar] [CrossRef]
- Van Mooy, B.A.S.; Fredricks, H.F.; Pedler, B.E.; Dyhrman, S.T.; Karl, D.M.; Koblízek, M.; Lomas, M.W.; Mincer, T.J.; Moore, L.R.; Moutin, T.; et al. Phytoplankton in the ocean use non-phosphorus lipids in response to phosphorus scarcity. Nature 2009, 458, 69–72. [Google Scholar] [CrossRef]
- Cañavate, J.P.; Armada, I.; Hachero-Cruzado, I. Interspecific variability in phosphorus-induced lipid remodelling among marine eukaryotic phytoplankton. New Phytol. 2017, 213, 700–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Adarme-Vega, T.C.; Lim, D.K.Y.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Fact. 2012, 11, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, G.J.; Hill, D.R.; Olmstead, I.L.; Bergamin, A.; Shears, M.J.; Dias, D.A.; Kentish, S.E.; Scales, P.J.; Botté, C.Y.; Callahan, D.L. Lipid profle remodeling in response to nitrogen deprivation in the microalgae Chlorella sp. (Trebouxi-ophyceae) and Nannochloropsis sp. (Eustigmatophyceae). PLoS ONE 2014, 9, e103389. [Google Scholar] [CrossRef] [Green Version]
- White, D.A.; Rooks, P.A.; Kimmance, S.; Tait, K.; Jones, M.; Tarran, G.A.; Cook, C.; Llewellyn, C.A. Modulation of polar lipid profiles in Chlorella sp. in response to nutrient limitation. Metabolites 2019, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Wen, F.; Liu, J. Transcriptomic and lipidomic analysis of an EPA-containing Nannochloropsis sp. PJ12 in response to nitrogen deprivation. Sci. Rep. 2019, 9, 4540. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Song, D.; Kind, T.; Ma, Y.; Hoefkens, J.; Fiehn, O. Lipidomic analysis of Chlamydomonas reinhardtii under nitrogen and sulfur deprivation. PLoS ONE 2015, 10, e0137948. [Google Scholar] [CrossRef] [Green Version]
- Abida, H.; Dolch, L.J.; Meï, C.; Villanova, V.; Conte, M.; Block, M.A.; Finazzi, G.; Bastien, O.; Tirichine, L.; Bowler, C.; et al. Membrane glycerolipid remodeling triggered by nitrogen and phosphorus starvation in Phaeodactylum tricornutum. Plant Physiol. 2015, 167, 118–136. [Google Scholar] [CrossRef] [Green Version]
- Matich, E.K.; Ghafari, M.; Camgoz, E.; Caliskan, E.; Pfeifer, B.A.; Haznedaroglu, B.Z.; Atilla-Gokcumen, G.E. Time-series lipidomic analysis of the oleaginous green microalga species Ettlia oleoabundans under nutrient stress. Biotechnol. Biofuels 2018, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Shemi, A.; Schatz, D.; Fredricks, H.F.; Van Mooy, B.A.; Porat, Z.; Vardi, A. Phosphorus starvation induces membrane remodeling and recycling in Emiliania huxleyi. New Phytol. 2016, 211, 886–898. [Google Scholar] [CrossRef] [Green Version]
- Degraeve-Guilbault, C.; Bréhélin, C.; Haslam, R.; Sayanova, O.; Marie-Luce, G.; Jouhet, J.; Corellou, F. Glycerolipid characterization and nutrient deprivation associated changes in the green picoalga Ostreococcus tauri. Plant Physiol. 2017, 173, 2060–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, P.; Zedler, J.B.; Donohoe, R.M. Nitrogen vs. phosphorus limitation of algal biomass in shallow coastal lagoons. Limnol. Oceanogr. 1993, 38, 906–923. [Google Scholar] [CrossRef]
- Alexa, A.; Rahnenfuhrer, J. TopGO: Enrichment Analysis for Gene Ontology. R Package Version 2.24.0. 2016. Available online: https://bioconductor.org/packages/release/bioc/html/topGO.html (accessed on 2 June 2020).
- Lee, D.Y.; Park, J.J.; Barupal, D.K.; Fiehn, O. System response of metabolic networks in Chlamydomonas reinhardtii to total available ammonium. Mol. Cell. Proteom. 2012, 11, 973–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Li, F.; Ge, F.; Tao, N.; Zhou, Q.; Wong, M. Mechanisms of ammonium assimilation by Chlorella vulgaris F1068: Isotope fractionation and proteomic approaches. Bioresour. Technol. 2015, 190, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Shang, C.; Zhu, S.; Wang, Z.; Qin, L.; Alam, M.A.; Xie, J.; Yuan, Z. Proteome response of Dunaliella parva induced by nitrogen limitation. Algal Res. 2017, 23, 196–202. [Google Scholar] [CrossRef]
- Chung, C.-C.; Hwang, S.-P.L.; Chang, J. Identification of a high-affinity phosphate transporter gene in a prasinophyte alga, Tetraselmis chui, and its expression under nutrient limitation. Appl. Environ. Microbiol. 2003, 69, 754–759. [Google Scholar] [CrossRef] [Green Version]
- Sawada, S.; Usuda, H.; Tsukui, T. Participation of inorganic orthophosphate in regulation of the ribulose-1,5-bisphosphate carboxylase activity in response to changes in the photosynthetic source-sink balance. Plant Cell Physiol. 1992, 33, 943–949. [Google Scholar]
- Kriventseva, E.V.; Kuznetsov, D.; Tegenfeldt, F.; Manni, M.; Dias, R.; Simão, F.A.; Zdobnov, E.M. OrthoDB v10: Sampling the diversity of animal, plant, fungal, protist, bacterial and viral genomes for evolutionary and functional annotations of orthologs. Nucleic Acids Res. 2019, 47, D807–D811. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Ferjani, A.; Suzuki, I.; Murata, N. The SphS-SphR two component system is the exclusive sensor for the induction of gene expression in response to phosphate limitation in synechocystis. J. Biol. Chem. 2004, 279, 13234–13240. [Google Scholar] [CrossRef] [Green Version]
- Dyhrman, S.T.; Chappell, P.D.; Haley, S.T.; Moffett, J.W.; Orchard, E.D.; Waterbury, J.B.; Webb, E.A. Phosphonate utilization by the globally important marine diazotroph Trichodesmium. Nature 2006, 439, 68–71. [Google Scholar] [CrossRef]
- Ral, J.P.; Colleoni, C.; Wattebled, F.; Dauvillée, D.; Nempont, C.; Deschamps, P.; Li, Z.; Morell, M.K.; Chibbar, R.; Purton, S.; et al. Circadian clock regulation of starch metabolism establishes GBSSI as a major contributor to amylopectin synthesis in Chlamydomonas reinhardtii. Plant Physiol. 2006, 142, 305–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Groot, C.C.; Van Den Boogaard, R.; Marcelis, L.F.; Harbinson, J.; Lambers, H. Contrasting effects of N and P deprivation on the regulation of photosynthesis in tomato plants in relation to feedback limitation. J. Exp. Bot. 2003, 54, 1957–1967. [Google Scholar] [CrossRef] [PubMed]
- Boot, R.G.A.; Schildwacht, P.M.; Lambers, H. Partitioning of nitrogen and biomass at a range of N-addition rates and their consequences for growth and gas exchange in to perennial grasses from inland dunes. Physiol. Plant 1992, 86, 152–160. [Google Scholar] [CrossRef]
- Guerrero, M.G.; Vega, J.M.; Losada, M. The assimilatory nitrate—Reducing system and its regulation. Annu. Rev. Plant Physiol. 1981, 32, 169–204. [Google Scholar] [CrossRef]
- Florencio, F.J.; Vega, J.M. Regulation of the assimilation of nitrate in Chlamydomonas reinhardii. Phytochemistry 1982, 21, 1195–1200. [Google Scholar] [CrossRef]
- Franco, A.R.; Cárdenas, J.; Fernández, E. Regulation by ammonium of nitrate and nitrite assimilation in Chlamydomonas reinhardtii. Biochem. Biophys. Acta 1988, 951, 98–103. [Google Scholar] [CrossRef]
- García-Fernández, J.M.; Diez, J. Adaptive mechanisms of nitrogen and carbon assimilatory pathways in the marine cyanobacteria Prochlorococcus. Res. Microbiol. 2004, 155, 795–802. [Google Scholar] [CrossRef]
- Riekhof, W.R.; Sears, B.B.; Benning, C. Annotation of genes involved in glycerolipid biosynthesis in Chlamydomonas reinhardtii: Discovery of the betaine lipid synthase BTA1(Cr). Eukaryot. Cell 2005, 4, 242–252. [Google Scholar] [CrossRef] [Green Version]
- Vogel, G.; Eichenberger, W. Betaine lipids in lower plants: Biosynthesis of DGTS and DGTA in Ochromonas danica (Chrysophyceaae) and the possible role of DGTS in lipid metabolism. Plant Cell Physiol. 1992, 33, 427–436. [Google Scholar]
- Fulton, J.M.; Fredricks, H.F.; Bidle, K.D.; Vardi, A.; Kendrick, B.J.; DiTullio, G.R.; Van Mooy, B.A. Novel molecular determinants of viral susceptibility and resistance in the lipidome of Emiliania huxleyi. Environ. Microbiol. 2014, 16, 1137–1149. [Google Scholar] [CrossRef]
- Martin, S.F.; Munagapati, V.S.; Salvo-Chirnside, E.; Kerr, L.E.; Le Bihan, T. Proteome turnover in the green alga Ostreococcus tauri by time course 15N metabolic labeling mass spectrometry. J. Proteome Res. 2012, 11, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Le Bihan, T.; Grima, R.; Martin, S.; Forster, T.; Le Bihan, Y. Quantitative analysis of low-abundance peptides in HeLa cell cytoplasm by targeted liquid chromatography/mass spectrometry and stable isotope dilution: Emphasising the distinction between peptide detection and peptide identification. Rapid Commun. Mass Spectrom. 2010, 24, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Kanonidis, E.I.; Roy, M.M.; Deighton, R.F.; Le Bihan, T. Protein co-expression analysis as a strategy to complement a standard quantitative proteomics approach: Case of a glioblastoma multiforme study. PLoS ONE 2016, 11, e0161828. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lee, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, S.F.; Doherty, M.K.; Salvo-Chirnside, E.; Tammireddy, S.R.; Liu, J.; Le Bihan, T.; Whitfield, P.D. Surviving Starvation: Proteomic and Lipidomic Profiling of Nutrient Deprivation in the Smallest Known Free-Living Eukaryote. Metabolites 2020, 10, 273. https://doi.org/10.3390/metabo10070273
Martin SF, Doherty MK, Salvo-Chirnside E, Tammireddy SR, Liu J, Le Bihan T, Whitfield PD. Surviving Starvation: Proteomic and Lipidomic Profiling of Nutrient Deprivation in the Smallest Known Free-Living Eukaryote. Metabolites. 2020; 10(7):273. https://doi.org/10.3390/metabo10070273
Chicago/Turabian StyleMartin, Sarah F., Mary K. Doherty, Eliane Salvo-Chirnside, Seshu R. Tammireddy, Jiaxiuyu Liu, Thierry Le Bihan, and Phillip D. Whitfield. 2020. "Surviving Starvation: Proteomic and Lipidomic Profiling of Nutrient Deprivation in the Smallest Known Free-Living Eukaryote" Metabolites 10, no. 7: 273. https://doi.org/10.3390/metabo10070273
APA StyleMartin, S. F., Doherty, M. K., Salvo-Chirnside, E., Tammireddy, S. R., Liu, J., Le Bihan, T., & Whitfield, P. D. (2020). Surviving Starvation: Proteomic and Lipidomic Profiling of Nutrient Deprivation in the Smallest Known Free-Living Eukaryote. Metabolites, 10(7), 273. https://doi.org/10.3390/metabo10070273