
Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review

Abstract
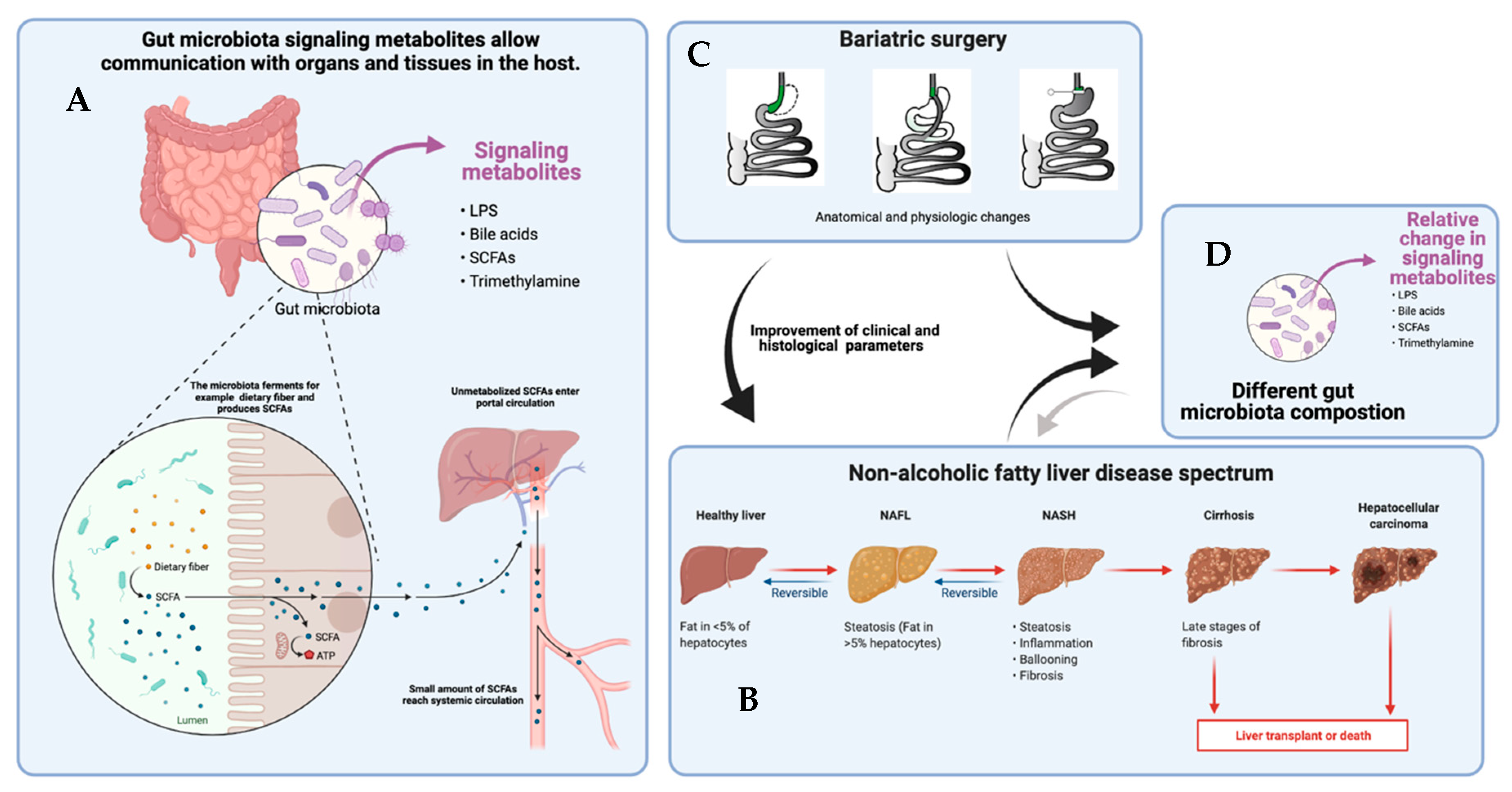
:1. Introduction
2. Gut Microbiome and Metabolites
3. Gut Microbiome and Metabolites in NAFLD
3.1. Fatty Liver Disease
3.2. Gut Microbiome in Non-Alcoholic Fatty Liver Diesease
3.3. Metabolites in Fatty Liver Disease
3.4. Causality
4. Gut Microbiome and Metabolites after Bariatric Surgery and Other Weight Loss Interventions
4.1. Bariatric Surgery and Other Weight Loss Interventions
4.2. Gut Microbiome in Obese Population and after Bariatric Surgery
4.3. Plasma Metabolites after Bariatric Surgery
5. NAFLD after Bariatric Surgery
6. Discussion and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sayiner, M.; Koenig, A.; Henry, L.; Younossi, Z.M. Epidemiology of Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis in the United States and the Rest of the World. Clin. Liver Dis. 2016, 20, 205–214. [Google Scholar] [CrossRef]
- Sberna, A.L.; Bouillet, B.; Rouland, A.; Brindisi, M.C.; Nguyen, A.; Mouillot, T.; Duvillard, L.; Denimal, D.; Loffroy, R.; Vergès, B.; et al. European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1388–1402. [Google Scholar]
- Setiawan, V.W.; Stram, D.O.; Porcel, J.; Lu, S.C.; Le Marchand, L.; Noureddin, M. Prevalence of chronic liver disease and cirrhosis by underlying cause in understudied ethnic groups: The multiethnic cohort. Hepatology 2016, 64, 1969–1977. [Google Scholar] [CrossRef] [Green Version]
- Chalasani, N.; Younossi, Z.; LaVine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef]
- Sayiner, M.; Younossi, Z.M. Nonalcoholic Steatohepatitis Is Becoming a Top Indication for Liver Transplantation Worldwide. Liver Transplant. 2019, 25, 10–11. [Google Scholar] [CrossRef] [Green Version]
- Syn, N.L.; Cummings, D.E.; Wang, L.Z.; Lin, D.J.; Zhao, J.J.; Loh, M.; Koh, Z.J.; Chew, C.A.; Loo, Y.E.; Tai, B.C.; et al. Association of metabolic–bariatric surgery with long-term survival in adults with and without diabetes: A one-stage meta-analysis of matched cohort and prospective controlled studies with 174 772 participants. Lancet 2021, 397, 1830–1841. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Prifti, E.; Belda, E.; Ichou, F.; Kayser, B.D.; Dao, M.C.; O Verger, E.; Hedjazil, L.; Bouillot, J.-L.; Chevallier, J.-M.; et al. Major microbiota dysbiosis in severe obesity: Fate after bariatric surgery. Gut 2019, 68, 70–82. [Google Scholar] [CrossRef]
- De Groot, P.; Scheithauer, T.; Bakker, G.J.; Prodan, A.; Levin, E.; Khan, M.T.; Herrema, H.; Ackermans, M.; Serlie, M.J.M.; de Brauw’, M.; et al. Gut microbiota Donor metabolic characteristics drive effects of faecal microbiota transplantation on recipient insulin sensitivity, energy expenditure and intestinal transit time. Gut 2019, 69, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Chen, C.; Cui, J.; Lu, J.; Yan, C.; Wei, X.; Zhao, X.; Li, N.; Li, S.; Xue, G.; et al. Fatty Liver Disease Caused by High-Alcohol-Producing Klebsiella pneumoniae. Cell Metab. 2019, 30, 675–688. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Abrahamsson, T.R.; Jenmalm, M.C.; Harris, K.; Quince, C.; Jernberg, C.; Björkstén, B.; Engstrand, L.; Andersson, A.F. Decreased gut microbiota diversity, delayed Bacteroidetes colonisation and reduced Th1 responses in infants delivered by Caesarean section. Gut 2014, 63, 559–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Stearns, J.C.; Lynch, M.D.J.; Senadheera, D.B.; Tenenbaum, H.C.; Goldberg, M.B.; Cvitkovitch, D.G.; Croitoru, K.; Moretno-Hagelsieb, G.; Neufeld, J.D. Bacterial biogeography of the human digestive tract. Sci. Rep. 2011, 1, 170. [Google Scholar] [CrossRef] [Green Version]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Al Khatib, H.; et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef]
- Wang, J.; Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 2020, 19, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Tabone, M.; Bressa, C.; García-Merino, J.A.; Moreno-Pérez, D.; Van, E.C.; Castelli, F.A.; Fetnaille, F.; Lalrrosa, M. The effect of acute moderate-intensity exercise on the serum and fecal metabolomes and the gut microbiota of cross-country endurance athletes. Sci. Rep. 2021, 11, 3558. [Google Scholar] [CrossRef]
- Overby, H.B.; Ferguson, J.F. Gut Microbiota-Derived Short-Chain Fatty Acids Facilitate Microbiota:Host Cross talk and Modulate Obesity and Hypertension. Curr. Hypertens. Rep. 2021, 23, 8. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe. 2015, 17, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Gibala, M.J. Protein metabolism and endurance exercise. Sports Med. 2007, 34, 337–340. [Google Scholar] [CrossRef]
- Zhou, M.; Shao, J.; Wu, C.-Y.; Shu, L.; Dong, W.; Liu, Y.; Chen, M.; Wynn, R.M.; Wang, J.; Wang, J.; et al. Targeting BCAA catabolism to treat obesity-associated insulin resistance. Diabetes 2019, 68, 1730–1746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyoötylaäinen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef]
- Meijnikman, A.S.; Gerdes, V.E.; Nieuwdorp, M.; Herrema, H. Evaluating causality of gut microbiota in obesity and diabetes in humans. Endocr. Rev. 2018, 39, 133–153. [Google Scholar] [CrossRef] [PubMed]
- Aydin, Ö.; Nieuwdorp, M.; Gerdes, V. The Gut Microbiome as a Target for the Treatment of Type 2 Diabetes. Curr. Diabetes Rep. 2018, 18, 55. [Google Scholar] [CrossRef] [Green Version]
- Hoyles, L.; Fernández-Real, J.M.; Federici, M.; Serino, M.; Abbott, J.; Charpentier, J.; Heymes, C.; Luque, J.L.; Anthony, E.; Barton, R.H.; et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat. Med. 2018, 24, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Sherriff, J.L.; O’Sullivan, T.A.; Properzi, C.; Oddo, J.L.; Adams, L.A. Choline, its potential role in nonalcoholic fatty liver disease, and the case for human and bacterial genes. Adv. Nutr. 2016, 7, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Wahlström, A.; Kovatcheva-Datchary, P.; Ståhlman, M.; Bäckhed, F.; Marschall, H.U. Crosstalk between Bile Acids and Gut Microbiota and Its Impact on Farnesoid X Receptor Signalling. Dig. Dis. 2017, 35, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Molinero, N.; Ruiz, L.; Sánchez, B.; Margolles, A.; Delgado, S. Intestinal bacteria interplay with bile and cholesterol metabolism: Implications on host physiology. Front. Physiol. 2019, 10, 185. [Google Scholar] [CrossRef] [Green Version]
- Chiang, J.Y.L.; Ferrell, J.M. Bile acid metabolism in liver pathobiology. Gene Expr. 2018, 18, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, T.R.; Haeusler, R.A. Bile acids in glucose metabolism and insulin signalling—mechanisms and research needs. Nat. Rev. Endocrinol. 2019, 15, 701–712. [Google Scholar] [CrossRef]
- Thomas, C.; Gioiello, A.; Noriega, L.; Strehle, A.; Oury, J.; Rizzo, G.; Macchiarulo, A.; Yamamoto, H.; Mataki, C.; Pruzanski, M.; et al. TGR5-Mediated Bile Acid Sensing Controls Glucose Homeostasis. Cell Metab. 2009, 10, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Sayin, S.I.; Wahlström, A.; Felin, J.; Jäntti, S.; Marschall, H.U.; Bamberg, K.; Angelin, B.; Hyötyläinen, T.; Orešič, M.; Bäckhed, M. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 2013, 17, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Cai, J.; Gonzalez, F.J. The role of farnesoid X receptor in metabolic diseases, and gastrointestinal and liver cancer. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 1–13. [Google Scholar] [CrossRef]
- Kuipers, F.; Bloks, V.W.; Groen, A.K. Beyond intestinal soap—Bile acids in metabolic control. Nat. Rev. Endocrinol. 2014, 10, 488–498. [Google Scholar] [CrossRef] [PubMed]
- van Nierop, F.S.; Scheltema, M.J.; Eggink, H.M.; Pols, T.W.; Sonne, D.P.; Knop, F.K.; Soeters, M.R. Clinical relevance of the bile acid receptor TGR5 in metabolism. Lancet Diabetes Endocrinol. 2017, 5, 224–233. [Google Scholar] [CrossRef]
- Stenman, L.K.; Holma, R.; Eggert, A.; Korpela, R. A novel mechanism for gut barrier dysfunction by dietary fat: Epithelial disruption by hydrophobic bile acids. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G227–G234. [Google Scholar] [CrossRef] [Green Version]
- Heianza, Y.; Ma, W.; Manson, J.A.E.; Rexrode, K.M.; Qi, L. Gut microbiota metabolites and risk of major adverse cardiovascular disease events and death: A systematic review and meta-analysis of prospective studies. J. Am. Heart Assoc. 2017, 6, e004947. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.C.Y.; Tyler, A.D. Systematic evaluation of supervised machine learning for sample origin prediction using metagenomic sequencing data. Biol. Direct. 2020, 15, 1–12. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Ziemski, M.; Robeson, M.S.; Kaehler, B.D. Measuring the microbiome: Best practices for developing and benchmarking microbiomics methods. Comput. Struct. Biotechnol. J. 2020, 18, 4048–4062. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.M.; Hanson, B.M.; Agresta, O.M.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]
- Herrema, H.; Niess, J.H. Intestinal microbial metabolites in human metabolism and type 2 diabetes. Diabetologia 2020, 63, 2533–2547. [Google Scholar] [CrossRef]
- Pittayanon, R.; Lau, J.T.; Leontiadis, G.I.; Tse, F.; Yuan, Y.; Surette, M.; Moayyetdi, P. Differences in Gut Microbiota in Patients With vs. Without Inflammatory Bowel Diseases: A Systematic Review. Gastroenterology 2020, 158, 930–946.e1. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver (EASL). EASL–EASD–EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. Diabetologia 2016, 59, 1121–1140. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Jiang, X.; Cao, M.; Ge, J.; Bao, Q.; Tang, L.; Chen, Y.; Li, L. Altered fecal microbiota correlates with liver biochemistry in nonobese patients with non-alcoholic fatty liver disease. Sci. Rep. 2016, 6, 32002. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Zheng, R.D.; Sun, X.Q.; Ding, W.J.; Wang, X.Y.; Fan, J.G. Gut microbiota dysbiosis in patients with non-alcoholic fatty liver disease. Hepatobiliary Pancreat Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef]
- Augustyn, M.; Grys, I.; Kukla, M. Small intestinal bacterial overgrowth and nonalcoholic fatty liver disease. Clin. Exp. Hepatol. 2019, 5, 1–10. [Google Scholar] [CrossRef]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NAS.H. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab. 2017, 25, 1054–1062.e5. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clément, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Raman, M.; Ahmed, I.; Gillevet, P.M.; Probert, C.S.; Ratcliffe, N.M.; Smith, S.; Greenwood, R.; Sikaroodi, M.; Lam, V.; Crotty, P.; et al. Fecal microbiome and volatile organic compound metabolome in obese humans with nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 2013, 11, 868–875.e3. [Google Scholar] [CrossRef]
- Ruuskanen, M.O.; Åberg, F.; Männistö, V.; Havulinna, A.S.; Méric, G.; Liu, Y.; Loomba, R.; Vázquez-Baeza, Y.; Tripathi, A.; Valsta, L.M.; et al. Links between gut microbiome composition and fatty liver disease in a large population sample. Gut Microbes 2021, 13, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Miele, L.; Valenza, V.; La Torre, G.; Montalto, M.; Cammarota, G.; Ricci, R.; Mascianà, R.; Forgione, A.; Gabrieli, M.L.; Perotti, G.; et al. Increased intestinal permeability and tight junction alterations in nonalcoholic fatty liver disease. Hepatology 2009, 49, 1877–1887. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, a.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Arrese, M.; Cabrera, D.; Kalergis, A.M.; Feldstein, A.E. Innate Immunity and Inflammation in NAFLD/NAS.H. Dig. Dis. Sci. 2016, 61, 1294–1303. [Google Scholar] [CrossRef] [Green Version]
- Fei, N.; Bruneau, A.; Zhang, X.; Wang, R.; Wang, J.; Rabot, S.; Gerard, P.; Zhao, L.P. Endotoxin producers overgrowing in human gut microbiota as the causative agents for nonalcoholic fatty liver disease. MBio 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Wen, S.W.; Kaminga, A.C.; Liu, A. Gut metabolites and inflammation factors in non-alcoholic fatty liver disease: A systematic review and meta-analysis. Sci. Rep. 2020, 10, 8848. [Google Scholar] [CrossRef]
- Caussy, C.; Tripathi, A.; Humphrey, G.; Bassirian, S.; Singh, S.; Faulkner, C.; Bettencourt, R.; Rizo, E.; Richards, L.; Xu, Z.Z.; et al. A gut microbiome signature for cirrhosis due to nonalcoholic fatty liver disease. Nat. Commun. 2019, 10, 1406. [Google Scholar] [CrossRef] [PubMed]
- Lanthier, N.; Rodriguez, J.; Nachit, M.; Hiel, S.; Trefois, P.; Neyrinck, A.M.; Cani, P.D.; Bindels, L.B.; Thissen, J.-P.; Delzenne, N.M. Microbiota analysis and transient elastography reveal new extra-hepatic components of liver steatosis and fibrosis in obese patients. Sci. Rep. 2021, 11, 659. [Google Scholar] [CrossRef] [PubMed]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.V.; Fizanne, L.; Araujo-Perez, F.; Guy, C.D.; Seed, P.C.; Rawls, J.F.; David, L.A.; et al. The severity of nonalcoholic fatty liver disease is associated with gut dysbiosis and shift in the metabolic function of the gut microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef] [Green Version]
- Schwimmer, J.B.; Johnson, J.S.; Angeles, J.E.; Behling, C.; Belt, P.H.; Borecki, I.; Bross, C.; Durelle, J.; Goyal, N.P.; Hamilton, G.; et al. Microbiome Signatures Associated With Steatohepatitis and Moderate to Severe Fibrosis in Children With Nonalcoholic Fatty Liver Disease. Gastroenterology 2019, 157, 1109–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astbury, S.; Atallah, E.; Vijay, A.; Aithal, G.P.; Grove, J.I.; Valdes, A.M. Lower gut microbiome diversity and higher abundance of proinflammatory genus Collinsella are associated with biopsy-proven nonalcoholic steatohepatitis. Gut Microbes 2020, 11, 569–580. [Google Scholar] [CrossRef]
- Lee, G.; You, H.J.; Bajaj, J.S.; Joo, S.K.; Yu, J.; Park, S.; Kang, H.; Park, J.H.; Kim, J.H.; Lee, D.H.; et al. Distinct signatures of gut microbiome and metabolites associated with significant fibrosis in non-obese NAFLD. Nat. Commun. 2020, 11, 4982. [Google Scholar] [CrossRef] [PubMed]
- Masarone, M.; Troisi, J.; Aglitti, A.; Torre, P.; Colucci, A.; Dallio, M.; Fetderico, A.; Balsano, C.; Persico, M. Untargeted metabolomics as a diagnostic tool in NAFLD: Discrimination of steatosis, steatohepatitis and cirrhosis. Metabolomics 2021, 17, 12. [Google Scholar] [CrossRef]
- Mardinoglu, A.; Agren, R.; Kampf, C.; Asplund, A.; Uhlen, M.; Nielsen, J. Genome-scale metabolic modelling of hepatocytes reveals serine deficiency in patients with non-alcoholic fatty liver disease. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mardinoglu, A.; Shoaie, S.; Bergentall, M.; Ghaffari, P.; Zhang, C.; Larsson, E.; Bäckhetd, F.; Nielsen, J. The gut microbiota modulates host amino acid and glutathione metabolism in mice. Mol. Syst. Biol. 2015, 11, 834. [Google Scholar] [CrossRef]
- Rom, O.; Liu, Y.; Liu, Z.; Zhao, Y.; Wu, J.; Ghrayeb, A.; Villacorta, L.; Fan, Y.; Chang, L.; Wang, L.; et al. Glycine-based treatment ameliorates NAFLD by modulating fatty acid oxidation, glutathione synthesis, and the gut microbiome. Sci. Transl. Med. 2020, 12, eaaz2841. [Google Scholar] [CrossRef] [PubMed]
- Coppola, S.; Avagliano, C.; Calignano, A.; Canani, R.B. The Protective Role of Butyrate against Obesity and Obesity-Related Diseases. Molecules 2021, 26, 682. [Google Scholar] [CrossRef] [PubMed]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.I.; Shankaran, M.; Yoshino, M.; Schweitzer, G.G.; Chondronikola, M.; Beals, J.W.; Okunade, A.L.; Patterson, B.W.; Nyangau, E.; Field, T.; et al. Insulin resistance drives hepatic de novo lipogenesis in nonalcoholic fatty liver disease. J. Clin. Invest. 2020, 130, 1453–1460. [Google Scholar] [CrossRef]
- Adams, L.A.; Wang, Z.; Liddle, C.; Melton, P.E.; Ariff, A.; Chandraratna, H.; Tan, J.; Ching, H.; Coulter, S.; de Boer, B.; et al. Bile acids associate with specific gut microbiota, low-level alcohol consumption and liver fibrosis in patients with non-alcoholic fatty liver disease. Liver Int. 2020, 40, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Nimer, N.; Choucair, I.; Wang, Z.; Nemet, I.; Li, L.; Gukasyan, J.; Weeks, T.L.; Alkhouri, N.; Nizar Zein, W.H.; Tang, W.; et al. Bile acids profile, histopathological indices and genetic variants for non-alcoholic fatty liver disease progression. Metabolism 2021, 116, 154457. [Google Scholar] [CrossRef]
- Puri, P.; Daita, K.; Joyce, A.; Mirshahi, F.; Santhekadur, P.K.; Cazanave, S.; A Luketic, V.; Siddiqui, M.S.; Boyett, S.; Min, H.; et al. The presence and severity of nonalcoholic steatohepatitis is associated with specific changes in circulating bile acids. Hepatology 2018, 67, 534–548. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.W.M.; Dallinga-Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozee, R.; et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef]
- Kootte, R.S.; Levin, E.; Salojärvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of Insulin Sensitivity after Lean Donor Feces in Metabolic Syndrome Is Driven by Baseline Intestinal Microbiota Composition. Cell Metab. 2017, 26, 611–619.e6. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Esteve, E.; Tremaroli, V.; Khan, M.T.; Caesar, R.; Mannerås-Holm, L.; Ståhlman, M.; Olsson, L.M.; Serino, M.; Planas-Fèlix, M.; et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat. Med. 2017, 23, 850–858. [Google Scholar] [CrossRef]
- B Barrow, F.; Khan, S.; Fredrickson, G.; Wang, H.; Dietsche, K.; Parthiban, P.; Robert, S.; Kaiser, T.; Winer, S.; Herman, A.; et al. Microbiota-Driven Activation of Intrahepatic B Cells Aggravates Nonalcoholic Steatohepatitis through Innate and Adaptive Signaling. Hepatology 2021. [Google Scholar] [CrossRef]
- Witjes, J.J.; Smits, L.P.; Pekmez, C.T.; Prodan, A.; Meijnikman, A.S.; Troelstra, M.A.; Bouter, K.E.C.; Herrema, H.; Levin, E.; Holleboom, A.G.; et al. Donor Fecal Microbiota Transplantation Alters Gut Microbiota and Metabolites in Obese Individuals with Steatohepatitis. Hepatol. Commun. 2020, 4, 1578–1590. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Xu, Z.; Mak, J.W.Y.; Yang, K.; Liu, Q.; Zuo, T.; Tang, W.; Lau, L.; Lui, R.N.; Wong, S.H.; et al. Microbiota engraftment after faecal microbiota transplantation in obese subjects with type 2 diabetes: A 24-week, double-blind, randomised controlled trial. Gut 2021. [Google Scholar] [CrossRef] [PubMed]
- Belgaumkar, A.P.; Vincent, R.P.; Carswell, K.A.; Hughes, R.D.; Alaghband-Zadeh, J.; Mitry, R.R.; Le Roux, C.W.; Pael, A.G. Changes in Bile Acid Profile After Laparoscopic Sleeve Gastrectomy are Associated with Improvements in Metabolic Profile and Fatty Liver Disease. Obes. Surg. 2016, 26, 1195–1202. [Google Scholar] [CrossRef]
- Caussy, C.; Hsu, C.; Lo, M.T.; Liu, A.; Bettencourt, R.; Ajmera, V.H.; Bassirian, S.; Hooker, J.; Sy, E.; Richards, L.; et al. Link between gut-microbiome derived metabolite and shared gene-effects with hepatic steatosis and fibrosis in NAFL.D. Hepatology 2018, 68, 918–932. [Google Scholar] [CrossRef] [Green Version]
- Sjöström, L.; Narbro, K.; Sjöström, C.D.; Karason, K.; Larsson, B.; Wedel, H.; Lystig, T.; Sullivan, M.; Bouchard, C.; Carlsson, B.; et al. Effects of bariatric surgery on mortality in Swedish obese subjects. N. Engl. J. Med. 2007, 357, 741–752. [Google Scholar] [CrossRef] [Green Version]
- Adams, T.D.; Davidson, L.E.; Litwin, S.E.; Kim, J.; Kolotkin, R.L.; Nanjee, M.N.; Gutierrez, J.M.; Frogley, S.J.; Ibele, A.R.; Brinton, E.A.; et al. Weight and metabolic outcomes 12 years after gastric bypass. N. Engl. J. Med. 2017, 377, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, L.; Chen, Y.; Lian, G.; Wang, J.; Zhang, J. Role of Gut Microbiome and Microbial Metabolites in Alleviating Insulin Resistance After Bariatric Surgery. Obes. Surg. 2021, 31, 327–336. [Google Scholar] [CrossRef]
- Yoshino, M.; Kayser, B.D.; Yoshino, J.; Stein, R.I.; Reeds, D.; Eagon, J.C.; Eckhouse, S.R.; Watrous, J.D.; Jain, M.; Knight, R.; et al. Effects of Diet versus Gastric Bypass on Metabolic Function in Diabetes. N. Engl. J. Med. 2020, 383, 721–732. [Google Scholar] [CrossRef]
- Meijnikman, A.S.; Aydin, O.; Prodan, A.; Tremaroli, V.; Herrema, H.; Levin, E.; Acherman, Y.; Bruin, S.; Gerdes, V.; Backhed, F.; et al. Distinct differences in gut microbial composition and functional potential from lean to morbidly obese subjects. J. Intern. Med. 2020, 288, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Hong, J.; Xu, X.; Feng, Q.; Zhang, D.; Gu, Y.; Shi, J.; Zhao, S.; Liu, W.; Wang, X.; et al. Gut microbiome and serum metabolome alterations in obesity and after weight-loss intervention. Nat. Med. 2017, 23, 859–868. [Google Scholar] [CrossRef]
- Koffert, J.; Lahti, L.; Nylund, L.; Salminen, S.; Hannukainen, J.C.; Salminen, P.; De Vos, W.M.; Nuutila, P. Partial restoration of normal intestinal microbiota in morbidly obese women six months after bariatric surgery. PeerJ. 2020, 8, e10442. [Google Scholar] [CrossRef]
- Steenackers, N.; Vanuytsel, T.; Augustijns, P.; Tack, J.; Mertens, A.; Lannoo, M.; Van der Schueren, B.; Matthys, C. Adaptations in gastrointestinal physiology after sleeve gastrectomy and Roux-en-Y gastric bypass. Lancet Gastroenterol. Hepatol. 2021, 6, 225–237. [Google Scholar] [CrossRef]
- Karami, R.; Kermansaravi, M.; Pishgahroudsari, M.; Talebi, M.; Mohammadzadeh, N.; Pazouki, A. Changes in gut microbial flora after Roux-en-Y gastric bypass and sleeve gastrectomy and their effects on post-operative weight loss. Updates Surg. 2020. [Google Scholar] [CrossRef] [PubMed]
- Steinert, R.E.; Rehman, A.; Souto Lima, E.J.; Agamennone, V.; Schuren, F.H.J.; Gero, D.; Schirnier, P.; Vonlanthen, R.; Ismaeil, A.; Tzafos, S.; et al. Roux-en-Y gastric bypass surgery changes fungal and bacterial microbiota in morbidly obese patients-A pilot study. PLoS ONE 2020, 15, e0236936. [Google Scholar] [CrossRef] [PubMed]
- Farin, W.; Oñate, F.P.; Plassais, J.; Bonny, C.; Beglinger, C.; Woelnerhanssen, B.; Nocca, D.; Magloues, F.; Le Chatelier, E.; Pons, N.; et al. Impact of laparoscopic Roux-en-Y gastric bypass and sleeve gastrectomy on gut microbiota: A metagenomic comparative analysis. Surg. Obes. Relat. Dis. 2020, 16, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Palleja, A.; Kashani, A.; Allin, K.H.; Nielsen, T.; Zhang, C.; Li, Y.; Brach, T.; Liang, S.; Feng, Q.; Jørgensen, N.B.; et al. Roux-en-Y gastric bypass surgery of morbidly obese patients induces swift and persistent changes of the individual gut microbiota. Genome. Med. 2016, 8, 67. [Google Scholar] [CrossRef] [Green Version]
- Barcenilla, A.; Pryde, S.E.; Martin, J.C.; Duncan, S.H.; Stewart, C.S.; Henderson, C.; Flint, H.J. Phylogenetic relationships of butyrate-producing bacteria from the human gut. Appl. Environ. Microbiol. 2000, 66, 1654–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Siles, M.; Duncan, S.H.; Garcia-Gil, L.J.; Martinez-Medina, M. Faecalibacterium prausnitzii: From microbiology to diagnostics and prognostics. ISME J. 2017, 11, 841–852. [Google Scholar] [CrossRef]
- Lenoir, M.; Martín, R.; Torres-Maravilla, E.; Chadi, S.; González-Dávila, P.; Sokol, H.; Langella, P.; Chain, F.; Bermúdez-Humarán, L.G. Butyrate mediates anti-inflammatory effects of Faecalibacterium prausnitzii in intestinal epithelial cells through Dact3. Gut Microbes. 2020, 12, 1826748. [Google Scholar] [CrossRef]
- Zhang, M.; Zhou, L.; Wang, Y.; Dorfman, R.G.; Tang, D.; Xu, L.; Pan, Y.D.; Zhou, Q.; Li, Y.; Yin, Y.Y.; et al. Faecalibacterium prausnitzii produces butyrate to decrease c-Myc-related metabolism and Th17 differentiation by inhibiting histone deacetylase 3. Int. Immunol. 2019, 31, 499–514. [Google Scholar] [CrossRef]
- Tabasi, M.; Eybpoosh, S.; Siadat, S.D.; Elyasinia, F.; Soroush, A.; Bouzari, S. Modulation of the Gut Microbiota and Serum Biomarkers After Laparoscopic Sleeve Gastrectomy: A 1-Year Follow-Up Study. Obes. Surg. 2021, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Karlsson, F.; Werling, M.; Ståhlman, M.; Kovatcheva-Datchary, P.; Olbers, T.; Fändriks, L.; Roux, C.W.; Nielsen, J.; Bäckhed, F. Roux-en-Y Gastric Bypass and Vertical Banded Gastroplasty Induce Long-Term Changes on the Human Gut Microbiome Contributing to Fat Mass Regulation. Cell Metab. 2015, 22, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Faria, S.L.; Santos, A.; Magro, D.O.; Cazzo, E.; Assalin, H.B.; Guadagnini, D.; Vieira, F.T.; Dutra, E.S.; Saad, M.J.A.; Ito, M.K. Gut Microbiota Modifications and Weight Regain in Morbidly Obese Women After Roux-en-Y Gastric Bypass. Obes. Surg. 2020, 30, 4958–4966. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.; Tsai, P.; Jüllig, M.; Liu, A.; Plank, L.; Booth, M. Differential Changes in Gut Microbiota After Gastric Bypass and Sleeve Gastrectomy Bariatric Surgery Vary According to Diabetes Remission. Obes. Surg. 2017, 27, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Shu, X.O.; Howard, E.F.; Long, J.; English, W.J.; Flynn, C.R. Fecal metagenomics and metabolomics reveal gut microbial changes after bariatric surgery. Surg. Obes. Relat. Dis. 2020, 16, 1772–1782. [Google Scholar] [CrossRef] [PubMed]
- Laferrère, B.; Reilly, D.; Arias, S.; Swerdlow, N.; Gorroochurn, P.; Bawa, B.; Bose, M.; Teixeira, J.; Stevens, R.D.; Wenner, B.R.; et al. Differential metabolic impact of gastric bypass surgery versus dietary intervention in obese diabetic subjects despite identical weight loss. Sci. Transl. Med. 2011, 3, 80re2. [Google Scholar] [CrossRef] [Green Version]
- Pakiet, A.; Wilczynski, M.; Rostkowska, O.; Korczynska, J.; Jabłonska, P.; Kaska, L.; Proczko-Stepaniak, M.; Sobczak, E.; Stepnowski, P.; Magkos, P.; et al. The Effect of One Anastomosis Gastric Bypass on Branched-Chain Fatty Acid and Branched-Chain Amino Acid Metabolism in Subjects with Morbid Obesity. Obes. Surg. 2020, 30, 304–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhabeeb, H.; AlFaiz, A.; Kutbi, E.; AlShahrani, D.; Alsuhail, A.; AlRajhi, S.; Alotaibi, N.; Alotaibi, K.; AlAmri, S.; Alghamdi, S.; et al. Gut hormones in health and obesity: The upcoming role of short chain fatty acids. Nutrients 2021, 13, 481. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Van Der Beek, C.M.; Jocken, J.W.E.; Goossens, G.H.; Holst, J.J.; Olde Damink, S.W.M.O.; Lenaerts, K.; DeJong, C.H.C.; Blalak, E.E. Colonic infusions of short-chain fatty acid mixtures promote energy metabolism in overweight/obese men: A randomized crossover trial. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Sowah, S.A.; Hirche, F.; Milanese, A.; Johnson, T.S.; Grafetstätter, M.; Schübel, R.; Kirsten, R.; Ulrich, C.M.; Kaaks, R.; Zeller, G.; et al. Changes in plasma short-chain fatty acid levels after dietary weight loss among overweight and obese adults over 50 weeks. Nutrients 2020, 12, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monte, S.V.; Caruana, J.A.; Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Schentag, J.J.; Dandona, P. Reduction in endotoxemia, oxidative and inflammatory stress, and insulin resistance after Roux-en-Y gastric bypass surgery in patients with morbid obesity and type 2 diabetes mellitus. Surgery 2012, 151, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Eiken, A.; Fuglsang, S.; Eiken, M.; Svane, M.S.; Kuhre, R.E.; Wewer Albrechtsen, N.J.W.; Hansen, S.H.; Trammell, S.A.J.; Svenningsen, J.S.; Rehfeld, J.F.; et al. Bilio-enteric flow and plasma concentrations of bile acids after gastric bypass and sleeve gastrectomy. Int. J. Obes. 2020, 44, 1872–1883. [Google Scholar] [CrossRef] [PubMed]
- Nickel, F.; Tapking, C.; Benner, L.; Sollors, J.; Billeter, A.T.; Kenngott, H.G.; Bokhary, L.; Schmid, M.; von Frankenberg, M.; Fischer, L.; et al. Bariatric Surgery as an Efficient Treatment for Non-Alcoholic Fatty Liver Disease in a Prospective Study with 1-Year Follow-up: BariScan Study. Obes. Surg. 2018, 28, 1342–1350. [Google Scholar] [CrossRef]
- Cherla, D.V.; Rodriguez, N.A.; Vangoitsenhoven, R.; Singh, T.; Mehta, N.; McCullough, A.J.; Brethauer, S.A.; Schauer, P.R.; Aminian, A. Impact of sleeve gastrectomy and Roux-en-Y gastric bypass on biopsy-proven non-alcoholic fatty liver disease. Surg. Endosc. 2020, 34, 2266–2272. [Google Scholar] [CrossRef]
- von Schönfels, W.; Beckmann, J.H.; Ahrens, M.; Hendricks, A.; Röcken, C.; Szymczak, S.; Hampe, J.; Schalfmayer, C. Histologic improvement of NAFLD in patients with obesity after bariatric surgery based on standardized NAS (NAFLD activity score). Surg. Obes. Relat. Dis. 2018, 14, 1607–1616. [Google Scholar] [CrossRef]
- Garg, H.; Aggarwal, S.; Shalimar; Yadav, R.; Gupta, S.D.; Agarwal, L.; Agarwal, S. Utility of transient elastography (fibroscan) and impact of bariatric surgery on nonalcoholic fatty liver disease (NAFLD) in morbidly obese patients. Surg. Obes. Relat. Dis. 2018, 14, 81–91. [Google Scholar] [CrossRef]
- Wirth, K.M.; Sheka, A.C.; Kizy, S.; Irey, R.; Benner, A.; Sieger, G. Bariatric Surgery is Associated With Decreased Progression of Nonalcoholic Fatty Liver Disease to Cirrhosis. Ann. Surg. 2020, 272, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Mazzini, G.S.; Khoraki, J.; Dozmorov, M.; Browning, M.G.; Wijesinghe, D.; Wolfe, L.; Gurski, R.R.; Campos, G.M. Concomitant PPARα and FXR Activation as a Putative Mechanism of NASH Improvement after Gastric Bypass Surgery: A GEO Datasets Analysis. J. Gastrointest. Surg. 2019, 23, 51–57. [Google Scholar] [CrossRef]
- Rebibo, L.; Gerin, O.; Verhaeghe, P.; Dhahri, A.; Cosse, C.; Regimbeau, J.M. Laparoscopic sleeve gastrectomy in patients with NASH-related cirrhosis: A case-matched study. Surg. Obes. Relat. Dis. 2014, 10, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Pouwels, S.; Parmar, C.; Kassir, R.; de Luca, M.; Graham, Y.; Mahawar, K.; for the Global Bariatric Research Collaborative. Outcomes of Bariatric Surgery in Patients with Liver Cirrhosis: A Systematic Review. Obes. Surg. 2021, 31, 2255–2267. [Google Scholar] [CrossRef] [PubMed]
- Glynn, E.L.; Piner, L.W.; Huffman, K.M.; Slentz, C.A.; Elliot-Penry, L.; Abouassi, H.; White, P.; Bain, J.R.; Muehlbauer, M.J.; Ilkayeva, O.R.; et al. Impact of combined resistance and aerobic exercise training on branched-chain amino acid turnover, glycine metabolism and insulin sensitivity in overweight humans. Diabetologia 2015, 58, 2324–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P.J.; Lapworth, A.L.; McGarrah, R.W.; Kwee, L.C.; Crown, S.B.; Ilkayeva, O.; An, J.; Carson, M.W.; Christopher, B.A.; Ball, J.R.; et al. Muscle-Liver Trafficking of BCAA-Derived Nitrogen Underlies Obesity-Related Glycine Depletion. Cell Rep. 2020, 33, 108375. [Google Scholar] [CrossRef]
- Yu, D.; Richardson, N.E.; Green, C.L.; Spicer, A.B.; Murphy, M.E.; Flores, V.; Jang, C.; Kasza, I.; Nikodemova, M.; Wakai, M.H.; et al. The adverse metabolic effects of branched-chain amino acids are mediated by isoleucine and valine. Cell Metab. 2021, 33, 905–922.e6. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhou, Y.; Wu, Z.; Li, Y.; Zhou, W.; Wang, Y. Integrated Analysis of Key Genes and Pathways Involved in Nonalcoholic Steatohepatitis Improvement After Roux-en-Y Gastric Bypass Surgery. Front. Endocrinol. 2021, 11, 611213. [Google Scholar] [CrossRef] [PubMed]
- Van Olden, C.C.; Van de Laar, A.W.; Meijnikman, A.S.; Aydin, O.; Van Olst, N.; Hoozemans, J.B.; De Brauw, L.M.; Bruin, S.C.; Acherman, Y.I.Z.; Verheij, J.; et al. A Systems Biology approach to understand gut microbiota and host metabolism in morbid obesity: Design of the BARIA Longitudinal Cohort Study. J. Intern. Med. 2020. [Google Scholar] [CrossRef]
- Sookoian, S.; Salatino, A.; Castaño, G.O.; Landa, M.S.; Fijalkowky, C.; Garaycoechea, M.; Pirola, G.J. Intrahepatic bacterial metataxonomic signature in non-alcoholic fatty liver disease. Gut 2020, 69, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Burcelin, R.; Tremaroli, V. Liver tissue microbiome in NAFLD: Next step in understanding the gut-–liver axis? . Gut 2020, 69, 1373–1374. [Google Scholar] [CrossRef]
- Verdier, C.; Denis, S.; Gasc, C.; Boucinha, L.; Uriot, O.; Delmas, D.; Dore, J.; Le Camus, C.; Schwintner, C.; Blanquet-Diot, S. An Oral FMT Capsule as Efficient as an Enema for Microbiota Reconstruction Following Disruption by Antibiotics, as Assessed in an In Vitro Human Gut Model. Microorganisms 2021, 9, 358. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Scorletti, E.; Afolabi, P.R.; Miles, E.A.; Smith, D.E.; Almehmadi, A.; Alshathry, A.; Childs, C.E.; Del Fabbro, S.; Bilson, J.; Moyses, H.E.; et al. Synbiotics Alter Fecal Microbiomes, But Not Liver Fat or Fibrosis, in a Randomized Trial of Patients with Nonalcoholic Fatty Liver Disease. Gastroenterology 2020, 158, 1597–1610.e7. [Google Scholar] [CrossRef] [PubMed]
- Sherf-Dagan, S.; Zelber-Sagi, S.; Zilberman-Schapira, G.; Webb, M.; Buch, A.; Keidar, A.; Sakran, N.; Goitein, D.; Goldenberg, N.; Mahdi, J.A.; et al. Probiotics administration following sleeve gastrectomy surgery: A randomized double-blind trial. Int. J. Obes. 2018, 42, 147–155. [Google Scholar] [CrossRef]
- Taur, Y.; Pamer, E.G. Harnessing microbiota to kill a pathogen: Fixing the microbiota to treat Clostridium difficile infections. Nat. Med. 2014, 20, 246–247. [Google Scholar] [CrossRef]
- Tixier, E.N.; Verheyen, E.; Luo, Y.; Grinspan, L.T.; Du, C.H.; Ungaro, R.C.; Walsh, S.; Grinspan, A.M. Systematic Review with Meta-Analysis: Fecal Microbiota Transplantation for Severe or Fulminant Clostridioides difficile. Dig. Dis. Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Bozadjieva Kramer, N.; Evers, S.S.; Shin, J.H.; Silverwood, S.; Wang, Y.; Burant, C.F.; Sandoval, D.A.; Seeley, R.J. The Role of Elevated Branched-Chain Amino Acids in the Effects of Vertical Sleeve Gastrectomy to Reduce Weight and Improve Glucose Regulation. Cell Rep. 2020, 33, 108239. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, S.N.; Luo, J.N.; Harris, D.A.; Aliakbarian, H.; Yao, L.; Paik, D.; Subramaniam, R.; Adhikari, A.A.; Vernon, A.H.; Kiliç, A.; et al. A microbial metabolite remodels the gut-liver axis following bariatric surgery. Cell Host Microbe. 2021, 29, 408–424.e7. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Li, Y.; Wang, Y.; Jin, L.; Yang, L.; Wang, L.; Liao, J.; Wang, H.; Peng, Y.; Zhang, Z.; et al. Da-Chai-Hu Decoction Ameliorates High Fat Diet-Induced Nonalcoholic Fatty Liver Disease Through Remodeling the Gut Microbiota and Modulating the Serum Metabolism. Front. Pharmacol. 2020, 11, 584090. [Google Scholar] [CrossRef] [PubMed]
- Littmann, E.R.; Lee, J.J.; Denny, J.E.; Alam, Z.; Maslanka, J.R.; Zarin, I.; Matsuda, R.; Carter, R.A.; Susac, B.; Saffern, M.S.; et al. Host immunity modulates the efficacy of microbiota transplantation for treatment of Clostridioides difficile infection. Nat. Commun. 2021, 12, 755. [Google Scholar] [CrossRef]
- Mazzini, G.S.; Khoraki, J.; Browning, M.G.; Wu, J.; Zhou, H.; Price, E.T.; Wolfe, L.G.; Mangino, M.J.; Campos, G.M. Gastric Bypass Increases Circulating Bile Acids and Activates Hepatic Farnesoid X Receptor (FXR) but Requires Intact Peroxisome Proliferator Activator Receptor Alpha (PPARα) Signaling to Significantly Reduce Liver Fat Content. J. Gastrointest. Surg. 2021, 25, 871–879. [Google Scholar] [CrossRef]
- Du, F.; Huang, R.; Lin, D.; Wang, Y.; Yang, X.; Huang, X.; Zheng, B.; Chen, Z.; Huang, Y.; Wang, X.; et al. Resveratrol Improves Liver Steatosis and Insulin Resistance in Non-alcoholic Fatty Liver Disease in Association With the Gut Microbiota. Front. Microbiol. 2021, 12, 611323. [Google Scholar] [CrossRef] [PubMed]
- Burz, S.; Monnoye, M.; Philippe, C.; Farin, W.; Ratziu, V.; Strozzi, F.; Paillarse, J.-M.; Chêne, L.; Blottière, H.; Gérard, P. Fecal Microbiota Transplant from Human to Mice Gives Insights into the Role of the Gut Microbiota in Non-Alcoholic Fatty Liver Disease (NAFLD). Microorg. 2021, 9, 199. [Google Scholar] [CrossRef]
- Meijnikman, A.S.; Bruin, S.; Groen, A.K.; Nieuwdorp, M.; Herrema, H. Increased expression of key SARS-CoV-2 entry points in multiple tissues in individuals with NAFLD. J. Hepatol. 2020, 74, 748. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Author, Year | Study Descriptive (Population; Follow-up; Weight Loss) | Method | Microbiome | Metabolites |
|---|---|---|---|---|
| Belgaumkar, 2016 [84] | Prospective analysis; SG (n = 18) NAFLD defined by serum cytokeratin 18 (n = 14, 78% NAFL) FU 6 months; TWL-39.9 kg | Bile acids: LC/MS | Not described. | ↑ primary glycine- and ↑ taurine-conjugated BA, ↓ cholic acid decreased, and ↑ secondary BA, ↑ glycine-conjugated urodeoxycholic acid No change in total BA. |
| Boursier, 2016 [64] | Biopsy-proven NAFLD (n = 57) F0/FI n = 30 vs. F3/F4 n = 27 | Fecal microbiome: 16 S RNA sequencing analysis | Increased NAFLD severity: ↑ Bacteroidaceae, ↓ Prevotellacea; ↓ Erysipelotrichaceae. NASH (compared to no NASH): ↑ Bacteroides; ↓ Prevotella Significant fibrosis (F3/4) compared to F0/F1: ↑ Bacteroides; ↑ Ruminococcus; ↓ Prevotella. | Not described. |
| Loomba, 2017 [53] | Prospective analysis biopsy-proven NAFLD (n = 86): comparison mild/moderate (n = 72;) vs. advanced fibrosis (n = 14); | Fecal microbiome: whole-genome shotgun sequencing | NAFLD—mild/moderate: ↑ abundance Firmicutes; most abundant Eubacterium rectale, Bacteroides vulgates NAFLD-AF: ↑ abundance Proteobacteria; most abundant B. vulgates, Escherichia coli. ↓ Ruminococcus obeum CAG:39; R. obeum; E. rectale. | NAFLD—mild/moderate: serum: ↑ Hypoxanthine, ↑ Inosine; Stool: ↑ L-lactate; ↑ Acetate ↑ formate; NAFLD-AF: serum: ↑ Succinate; ↑ Malatae; ↑ alfa-ketoglutarate; ↑ Serine; ↑ Glutamine; ↑ Fumarate; ↑ Glutamate; ↑ Lactate; stool: ↑ butyrate, D-lactate, propionate, succinate |
| Caussy, 2018 [85] | Cross-sectional analysis twin family cohort, n = 156 validation cohort, n = 156 hepatic steatosis, n = 57 | Fecal microbiome: whole-genome shotgun metagenomic sequencing; Liver: MRI/MRE; Metabolites CG/MS and LC/MS/MS | Proteobacteria, Firmicutes, Bacteroidetes correlated with 3-(4-hydroxyphenyl)lactate and phenyllactate. | 6 microbial origins: 3-(4-hydroxyphenyl)lactate, N-formylmethionine, phenyllactate, mannitol, allantoine, N-(2-furoyl)glycine. 3-(4-hydroxyphenyl)lactate gut microbiome-linked metabolite assosated with liver fibrosis. |
| Caussy, 2019 [62] | Cross-sectional; n = 203 NAFLD-cirrhosis, NAFLD, without advanced fibrosis non-NAFLD controls | Fecal microbiome: 16S RNA sequencing analysis Liver: MRI/MRE. | NAFLD–cirrhosis: ↑ Streptococcus; ↑ Megashaera; most enriched abundance of family Enterobacteriaceae, genera Streptococci and Gallibacterium. NAFLD-AF: ↑ Streptococcus; ↑ Bacillus; ↑ Lactococcus Non-NAFLD: ↑ Bacillus; ↑ Lactococcus; ↑ Pseudomonas; ↑ Faecalibacterium prausnitzii, ↑ genus Catenibacterium; families ↑ Rikenellaceae, ↑ Mogibacterium, ↑ Peptostreptococcaceae | Not described. |
| Puri, 2018 [77] | Cross-sectional analysis biopsy-proven NAFLD and bile acids; n = 86 (controls n = 24. NAFL n = 25; NASH n = 37; BMI 31.9) | LC/MS | Not described. | NASH: ↑ total primary BAs; ↓ secondary BAs. NASH vs. NAFL, vs. controls: ↑ Total conjugated primary BAs ↑ conjugated/unconjugated chenodeoxycholate; ↑ cholate; ↑ total primary BAs. NAFL: ↑ Total cholate/chenodeoxycholate ratio ↑ total secondary/primary BA ratio -> ↓ likelihood of significant fibrosis (F ≥ 2) ↑ conjugated cholate -> ↑ likelihood of significant fibrosis (F ≥ 2). |
| Hoyles, 2018 [29] | Prospective analysis; obese women n= 105; liver biopsy (histology), NAFLD (n = 56); fecal microbiome (n =56, | Fecal microbiome: shotgun metagenomic sequencing; serum and urine Metabolites: LC/MS | Steatosis: ↑ Proteobacteria, ↑ Actinobacteria, Verrucomicrobia ↑ correlated Firmicutes and Euryarchaeta ↓ correlated. Species: ↑ Acidaminococcus, ↑ Escherichia; ↓ Lachnospiraceae, ↓ Ruminococcaceae Functional analysis: ↑ LPS and peptidoglycan biosynthesis. | Steatosis: Serum BCAAs: ↑ leucine, ↑ valine, ↑ isoleucine. ↑ phenylacetic acid (PAA) Urine: ↑ choline No-NAFLD: ↑ acetate; ↑ TMAO |
| Lee, 2020 [67] | Prospective analysis Non-obese NAFLD | Fecal metabolites: 16S RNA sequencing analysis | Elevated Ruminococcaceae and Veillonellaceae associated with fibrosis severity. | Fecal metabolites: bile acids and propionate elevated (especially with significant fibrosis). |
| Adams, 2020 [75] | Prospective analysis liver biopsy n = 122 (as part of clinical care or during bariatric surgery) | Fecal microbiome: 16S RNA sequencing analysis Metabolies: serum + fecal BA analysis: LCMS. | NAFLD-AF (F3/4): ↑ Firmicutes, ↑ Proteobacteria; ↑ Actinobacteria; ↓ Bacteriodetes. Family: ↑ Actinomycetaceae; ↑ Lachnospiraceae; ↓ Bacteroidaceae; ↓ unclassifiable of order Bacteroidales. | Progressive ↑ total serum BAs from controls, F0–2 NAFLD to F3/4 NAFLD. ↑ GCA (glycocholic acid); ↑ GDCA (glycodeoxycholic acid) Fecal BA: ↑ DCA (deoxycholic acid); ↑ LC (lithocholic acid). |
| Masarone, 2021 [68] | Cross-sectional analysis cohort biopsy-proven NAFLD n = 144 steatosis, n = 76, NASH n = 23, cirrhosis, n = 43 (NASH–cirrhosis n = 15, HCV n = 8, cryptogenic n = 20) | Serum metabolites GC/MS; machine learning model. | Not described. | Lower in controls and increase with disease progression: isocitric acid, isoleucine, not identified metabolite Higher in controls and decrease with disease progression: xanthine, glutathione, glycolic acid Valine, asparagine, 4-deoxy erythronic acid, propanoic acid, palmitic acid, butanoic acid, stearic acid, phenylalanine, taurocholic acid NASH-related cirrhosis, increased concentration of galactose, uric acid, glyceric acid, butanoic acid, histidine, phenylalanine, stearic acid, threonine and palmitic acid |
| Nimer, 2021 [76] | Prospective analysis; NAFLD n = 102 (30% simple steatosis, 43% borderline NASH, 27% NASH); controls n = 50 Liver biopsy; BMI 32.8 kg/m2 | Plasma bile acid profile: quantitative stable isotope dilution LC/MS/MS | Not described. | NAFLD vs. controls: ↑ almost all circulating BAs Fibrosis vs. NAFLD: ↑ glycine-conjugated primary BAs (↑ GCDCA, ↑ GCA), secondary BAs ↑ 7-keto-DCA, ↑ GUDCA NASH vs. simple steatosis: ↑ 7-keto-DCA, ↑ 7-keto, LCA |
| First author, year [Ref.] | Study Descriptive (Population; Follow-up; Weight Loss) | Method | Results Microbiome | Results Metabolites |
|---|---|---|---|---|
| Laferrere, 2011 [108] | Prospective analysis; diabetic patients; RYGB, n = 10 Diet intervention, n = 11 FU 1 month | Plasma metabolites: MS | Not described. | RYGB: ↓ BCAAs: ↓ leucine, ↓ isoleucine, ↓ valine; ↓ aromatic AAs: ↓ phenylalanine ↓ tyrosine; ↓ ornithine, ↓ citrulline, ↓ histidine. Diet: no differences |
| Tremaroli, 2015 [104] | Post-bariatric surgery, long-term effect: RYGB (n = 7) vs. VGB (n = 7); matched controls (severe obesity n = 7); FU 9.4 years | Fecal microbiome: shotgun metagenomic sequencing Metabolites: GC/MS, UPLC-MS-MS | RYGB vs. obese controls: ↑ Gammaproteobacteria; ↓ Firmicutes (↓ Clostridium difficile, ↓ Clostridium hiranonis, ↓ Gemella sanguinis); ↑ Proteobacteria (↑ Escherichia, ↑ Klebsiella, ↑ Pseudomonas) No significant differences VBG and controls. No significant differences VBG and RYGB. | RYGB, VBG: ↓SCFAs (↓ acetate, ↓ propionate, ↓ butyrate) BCFA (isobutryrate, isovaleratie): no change. RYGB: SCFA/BCFA ratio decreased |
| Palleja, 2016 [98] | Prospective analysis; RYGB, n = 13 FU 3 months, n = 12 12 months, n = 8 | Fecal microbiome: Shotgun metagenomic sequencing | Microbial diversity: ↑ 3 months and ↑ 1 year. ↑ Proteobacteria, ↑ Fusobacteria ↑ Escherichia coli, ↑ Klebsiella pneumoniae, ↑ Veillonella spp., ↑ Streptococcus spp., ↑ Alistipes spp., and ↑ Enterococcus faecalis; ↑ Bifidobacterium dentium; ↑ Fusobacterium nucleatum; ↑ Akkermansia muciniphila; ↓ Faecalibacterium prausnitzii | Not described. |
| Liu, 2017 [92] | Prospective analysisSG, n = 23 obese individuals, n = 72 lean controls, n = 79 FU 1 and 3 months. | Shotgun metagenomic sequencing; serum plasma metabolites: LC/MS | ↑ : C. comes; D. longicatena; Clostridiales bacterium; Anaerotruncus colihominis; Akkermansia muciniphila; B. thetaiotaomicron. | ↓ : Aromatic amino acids ↓; methionine ↓; alanine ↓; lysine ↓; serine ↓; glutamate ↓ decreased ↑ : Acetylglycine ↑; glycine ↑increased. |
| Aron, 2019 [7)] | Prospective analysisRYGB, n = 41 gastric banding/AGB, n = 20) n = 24 had post-operative follow-up. | Fecal microbiome: shotgun metagenomic sequencing Serum metabolites: LC/MS | RYGB: ↑ Oscillibacter; ↑ Clostridium sp; ↑ Alistipes shahii; ↑ Butyricimonas; ↑ Fusobacterium nucleatum; ↑ Roseburia; ↑ Dialister sp; ↑ Butyricimonas virosa; ↑ Hungatella hatewayi; ↓ Coprobacillus sp.; ↓ Anaerostipes hadrus (butyrate producer)AGB: ↑ Oscillibacter; ↑ Butyricimonas virosa; ↑ Bacteroides finegoldii | Metabolites associated with microbial gene richness (MGR): glutamate, negatively correlated; 3-methoxyphenylacetic acid, phloretate, hippurate, 3-hydroxphenylacetate, L-histidin and three unidentified positively correlated.. RYGB: ↑ glycine, ↑ acetylglycine, ↑ methylmalonate After BS: - ↑ Acetlyglycine and ↑ glycine: - ↓ BCAA when insulin resistance ↑ |
| Steinert, 2020 [96] | Prospective n= 25, RYGB, n = 16 (RYGB), Controls, n= 9 FU 3 months, not described | Fecal microbiome: 16S RNA sequencing analysis | RYGB vs. pre-operation: ↓ Blautia, ↓ Roseburia and ↓ Faecalibacterium (Firmicutes); compared to controls more abundant RYGB: ↓ Bifidobacterium, which was increased compared to lean controls | Not described. |
| Farin, 2020 [97] | RYGB (n = 89) SG (n= 108) FU 6 months; not described | Fecal microbiome: shotgun metagenomic sequencing | Phylum: 15% gene enrichment: ↑ Bacteroidetes, ↑ Proteobacteria, ↑ Actinobacteria, ↑ Verrucomicrobia; ↓ Firmicutes (Firmicutes slight decrease after surgeries).Both: ↑ A. muciniphila; ↑ E. coli, ↑ H. parainfluenzae, ↑ Klebsiella pneuoniae. RYGB > SG: ↑ E. coli and ↑ K. pneumoniae, ↑ R. faecis and ↑ R. hominis, ↑ E. faecalis more enriched by RYGB than SG. ↓ Faecalibacterium preusnitzii less abundant after RYGB. SG > RYGB: ↑ A. hadrus, C. sp KLE, F. plautii, O. sp. KLE, R. gnavus (Firmicutes order Clostridiales) | Not described. |
| Karami, 2020 [95] | Prospective analysis n = 30 RYGB, n = 12 SG, n = 18 FU 6 months; EWL 57.72 ± 15.08% | Fecal microbiome: 16S RNA sequencing analysis | RYGB: Bacteroidetes ↓ compared to pre-surgery. RYGB or SG: no changes in Firmicutes count. Firmicutes to Bacteroidetes ratio not different from baseline; B. fragilis count not different to baseline or between groups. BS (RYGB + SG): ↓ Bacteroidetes; ↑ Firmicutes/Bacteroidetes ratio. | Not described. |
| Faria, 2020 [105] | Retrospective analysis; RYGB: with vs. without weight regain, vs. control; FU > 5 year post-operative; TWL% 25.8 vs. 33.1. | Fecal microbiome: 16S RNA sequencing analysis. | RYGB, non-regain vs. control and weight regain: ↑ Akkermansia genus. RYGB, non-regain vs. control: ↑ Phascolarctobacterium genus and ↓ SMB53 genus. RYGB vs. control: ↓ Bacteroidetes Control vs. RYGB: ↑ Bacteroides, ↑ SMB53 | Not described. |
| Pakiet, 2020 [109] | Prospective analysis OAGB, n = 50 lean controls, n = 32 | Serum metabolites GM/LM/MS | Not described. | Baseline vs. controls: ↓BCFAs; ↑ BCAAs OAGB: ↑ BCFA; ↓BCAA. IR correlated inversely with BCFAs and positively with BCAAs |
| Tabasi, 2021 [103] | Prospective analysis SG, n = 126 FU 3 and 12 months | Fecal microbiome: qPCR | M3: ↓ Prevotella; ↓ Bacteroides fragilis group; ↓ Firmicutes spp; ↑ Akkermansia muciniphila; ↑ Roseburia spp.; ↑ Bacteroidetes; ↑ Bifidobacterium M12: ↑ Actinobacteria (compared to M3 and baseline) | Not described. |
| Author, Year [Ref.] | Study Design | Parameters to Assess Liver Disease | Changes/Outcomes |
|---|---|---|---|
| Nickel, 2018 [115] | Prospective analysis, n = 100 SG vs. RYGB FU 1 year. | Transient elastography: liver stiffness Laboratory-based fibrosis score: AST/ALT ratio; NAFLD fibrosis score; APRI score; BARD score | 12.9 ± 10.4 vs. 7.1 ± 3.7 kPa; (RYGB > SG) 0.8 ± 0.3 vs. 1.1 ± 0.4 −1.0 ± 1.8 vs. −1.7 ± 1.3; 0.3 ± 0.2 vs. 0.3 ± 0.1; 2.3 ± 1.2 vs. 2.8 ± 1.1. |
| Garg, 2018 [118] | Prospective analysis Bariatric surgery population n = 76 FU 1 year (32 biopsies) | Controlled attenuation parameter (CAP) Transient elastography: liver stiffness Liver biopsy | Pre-operation: 326.5 (301–360.5) dB/m; Success rate pre-operation 87.9% Pre-operation 7.0 (5.0–9.5) kPa Improvement in hepatic steatosis, lobular inflammation, ballooning and fibrosis; NASH resolved in 3 out of 4 patients. |
| Cherla, 2020 [116] | Prospective analysis of biopsy-proven NAFLD SG, n = 62 RYGB, n = 248 FU 4 years (median) | Liver function tests AST, SG AST, RYGB ALT, SG ALT, RYGB | 84% normalized after bariatric surgery 49.1 ± 21.5 to 28.0 ± 16.5 49.3 ± 22.0 to 26.5 ± 15.5 61.7 ± 30.0 to 27.2 ± 21.5 59.4 ± 24.9 to 26.1 ± 19.2 |
| Wirth, 2020 [119] | Retrospective analysis; Bariatric surgery, n = 2942 Control, n = 5884 FU 31 months | Risk of NAFLD progression to cirrhosis after bariatric surgery | Reduced risk: HR 0.31 (95% CI 0.19–0.52) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoozemans, J.; de Brauw, M.; Nieuwdorp, M.; Gerdes, V. Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review. Metabolites 2021, 11, 353. https://doi.org/10.3390/metabo11060353
Hoozemans J, de Brauw M, Nieuwdorp M, Gerdes V. Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review. Metabolites. 2021; 11(6):353. https://doi.org/10.3390/metabo11060353
Chicago/Turabian StyleHoozemans, Jacqueline, Maurits de Brauw, Max Nieuwdorp, and Victor Gerdes. 2021. "Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review" Metabolites 11, no. 6: 353. https://doi.org/10.3390/metabo11060353
APA StyleHoozemans, J., de Brauw, M., Nieuwdorp, M., & Gerdes, V. (2021). Gut Microbiome and Metabolites in Patients with NAFLD and after Bariatric Surgery: A Comprehensive Review. Metabolites, 11(6), 353. https://doi.org/10.3390/metabo11060353