
Correlation of Cerebral Microdialysis with Non-Invasive Diffuse Optical Cerebral Hemodynamic Monitoring during Deep Hypothermic Cardiopulmonary Bypass

 ,
,  , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Animal Model
2.2. Neurological Monitoring
2.3. Cardiopulmonary Bypass and Deep Hypothermia
2.4. Diffuse Optical Monitoring of Cerebral Hemodynamics
2.5. Cerebral Microdialysis
2.6. Statistical Analysis
2.6.1. Longitudinal Changes in Cerebral Physiology
2.6.2. Correlation between Cerebral Hemodynamics and Biomarkers of Neurological Injury
2.6.3. Non-Invasive Predictors of Cerebral Metabolic Distress and Injury
3. Results
3.1. Summary of Experimental Characteristics
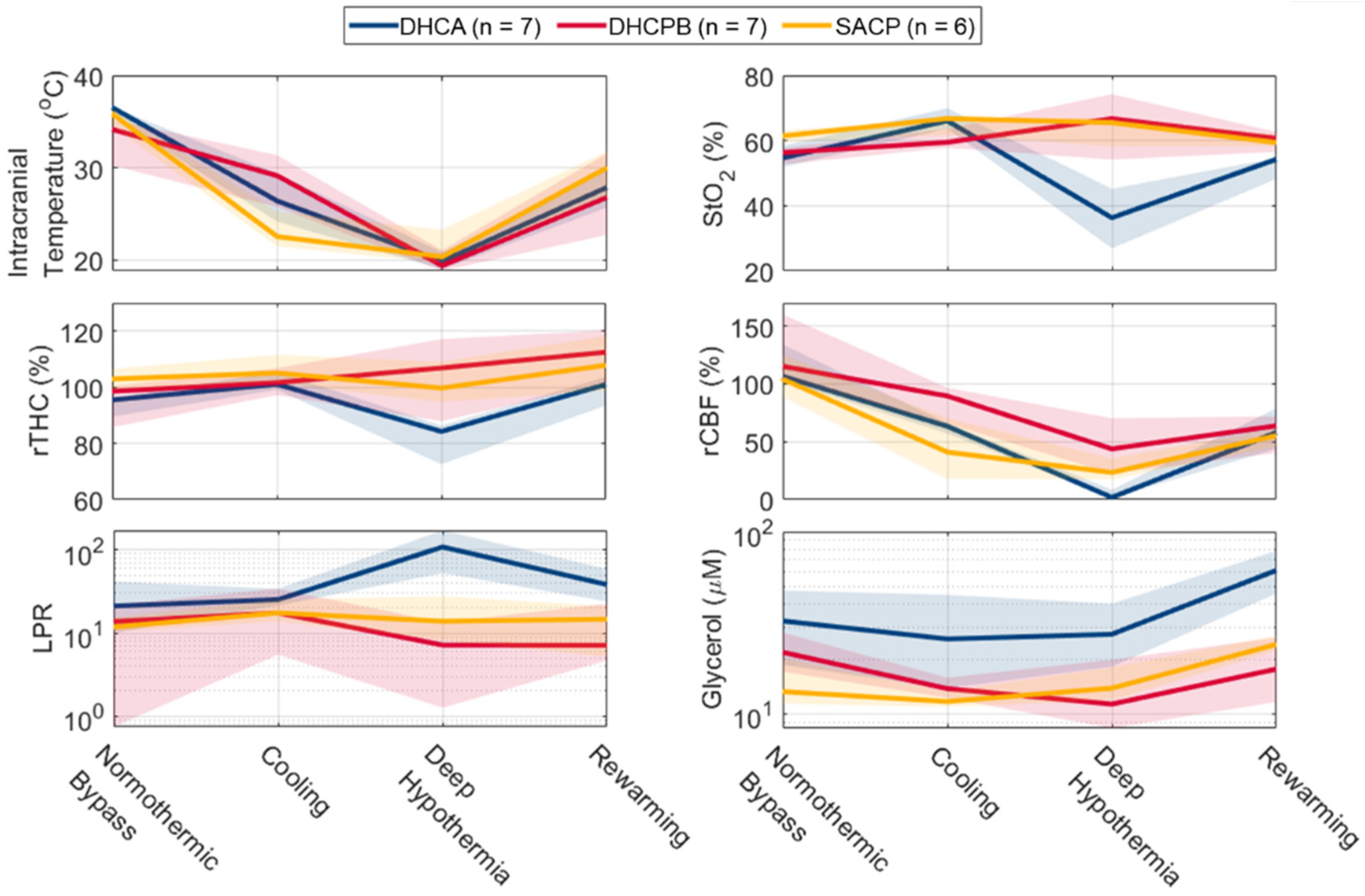
3.2. Longitudinal Changes in Cerebral Physiology
3.3. Correlation between Cerebral Hemodynamics and Biomarkers of Neurological Injury
3.3.1. Lactate–Pyruvate Ratio (LPR)
3.3.2. Glycerol
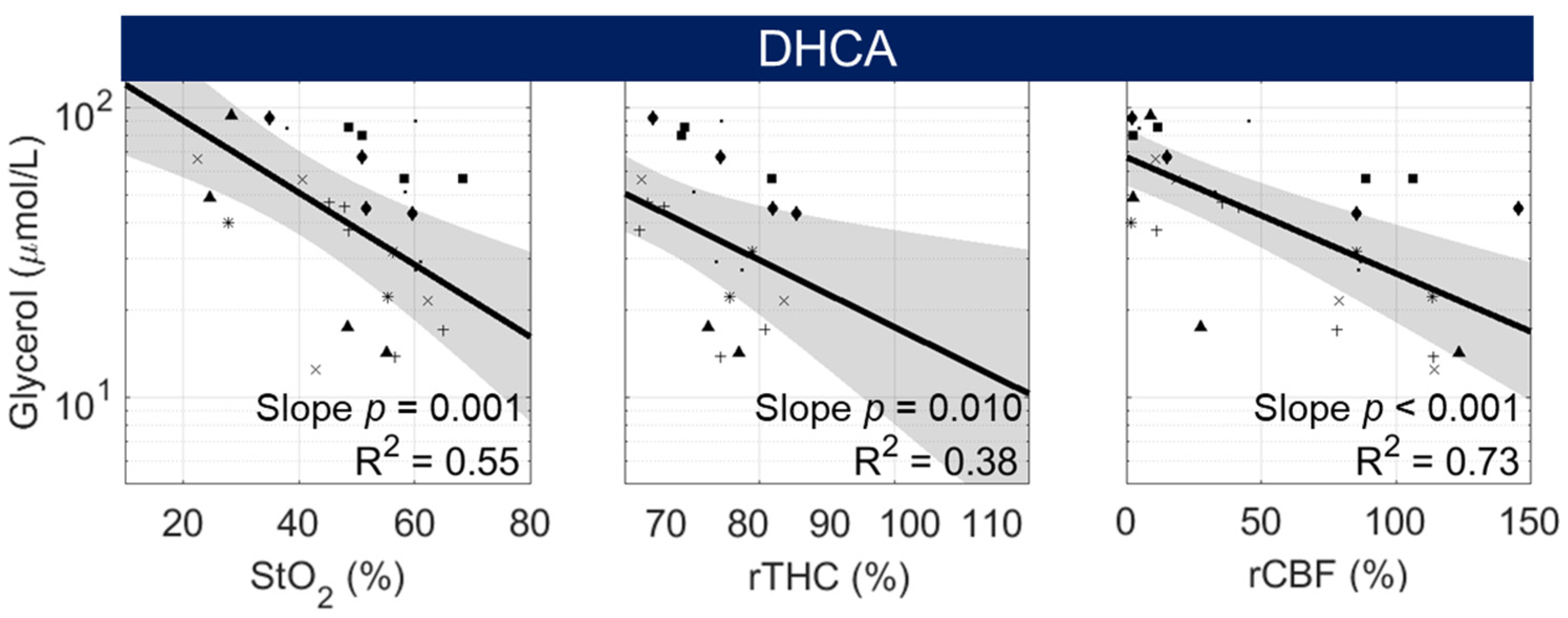
3.4. Non-Invasive Predictors of Cerebral Metabolic Distress and Injury
4. Discussion
Limitations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Owen, M.; Shevell, M.; Donofrio, M.; Majnemer, A.; McCarter, R.; Vezina, G.; Bouyssi-Kobar, M.; Evangelou, I.; Freeman, D.; Weisenfeld, N.; et al. Brain Volume and Neurobehavior in Newborns with Complex Congenital Heart Defects. J. Pediatr. 2014, 164, 1121–1127.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaynor, J.W.; Stopp, C.; Wypij, D.; Andropoulos, D.B.; Atallah, J.; Atz, A.M.; Beca, J.; Donofrio, M.T.; Duncan, K.; Ghanayem, N.S.; et al. Neurodevelopmental Outcomes After Cardiac Surgery in Infancy. Pediatrics 2015, 135, 816–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Licht, D.J.; Shera, D.M.; Clancy, R.R.; Wernovsky, G.; Montenegro, L.M.; Nicolson, S.C.; Zimmerman, R.A.; Spray, T.L.; Gaynor, J.W.; Vossough, A. Brain Maturation Is Delayed in Infants with Complex Congenital Heart Defects. J. Thorac. Cardiovasc. Surg. 2009, 137, 529–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinney, H.C. The Near-Term (Late Preterm) Human Brain and Risk for Periventricular Leukomalacia: A Review. Semin. Perinatol. 2006, 30, 81–88. [Google Scholar] [CrossRef]
- Galli, K.K.; Zimmerman, R.A.; Jarvik, G.P.; Wernovsky, G.; Kuypers, M.K.; Clancy, R.R.; Montenegro, L.M.; Mahle, W.T.; Newman, M.F.; Saunders, A.M.; et al. Periventricular Leukomalacia Is Common after Neonatal Cardiac Surgery. J. Thorac. Cardiovasc. Surg. 2004, 127, 692–704. [Google Scholar] [CrossRef] [Green Version]
- Beca, J.; Gunn, J.K.; Coleman, L.; Hope, A.; Reed, P.W.; Hunt, R.W.; Finucane, K.; Brizard, C.; Dance, B.; Shekerdemian, L.S. New White Matter Brain Injury after Infant Heart Surgery Is Associated with Diagnostic Group and the Use of Circulatory Arrest. Circulation 2013, 127, 971–979. [Google Scholar] [CrossRef] [Green Version]
- Algra, S.O.; Jansen, N.J.G.; Van Der Tweel, I.; Schouten, A.N.J.; Groenendaal, F.; Toet, M.; Van Oeveren, W.; Van Haastert, I.C.; Schoof, P.H.; De Vries, L.S.; et al. Neurological Injury after Neonatal Cardiac Surgery: A Randomized, Controlled Trial of 2 Perfusion Techniques. Circulation 2014, 129, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Mavroudis, C.D.; Ko, T.; Volk, L.E.; Smood, B.; Morgan, R.W.; Lynch, J.M.; Davarajan, M.; Boorady, T.W.; Licht, D.J.; Gaynor, J.W.; et al. Does Supply Meet Demand? A Comparison of Perfusion Strategies on Cerebral Metabolism in a Neonatal Swine Model. J. Thorac. Cardiovasc. Surg. 2022, 163, e47–e58. [Google Scholar] [CrossRef]
- Alsoufi, B. Commentary: Everyone Is Winging It, Some Just Do It More Confidently. J. Thorac. Cardiovasc. Surg. 2022, 163, e59–e60. [Google Scholar] [CrossRef]
- Huisenga, D.; La Bastide-Van Gemert, S.; Van Bergen, A.; Sweeney, J.; Hadders-Algra, M. Developmental Outcomes after Early Surgery for Complex Congenital Heart Disease: A Systematic Review and Meta-Analysis. Dev. Med. Child Neurol. 2021, 63, 29–46. [Google Scholar] [CrossRef]
- Mavroudis, C.D.; Karlsson, M.; Ko, T.S.; Hefti, M.; Gentile, J.I.; Morgan, R.W.; Plyler, R.; Mensah-Brown, K.G.; Boorady, T.W.; Melchior, R.W.; et al. Cerebral Mitochondrial Dysfunction Associated with Deep Hypothermic Circulatory Arrest in Neonatal Swine. Eur. J. Cardio-Thorac. Surg. 2018, 54, 162–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley, S.M.; Chai, P.J.; Miller, S.E.; Mault, J.R.; Jaggers, J.J.; Tsui, S.S.; Lodge, A.J.; Lefurgey, A.; Ungerleider, R.M. Intermittent Perfusion Protects the Brain during Deep Hypothermic Circulatory Arrest. Ann. Thorac. Surg. 1999, 68, 4–12. [Google Scholar] [CrossRef]
- Priestley, M.A.; Golden, J.A.; O’Hara, I.B.; McCann, J.; Kurth, C.D. Comparison of Neurologic Outcome after Deep Hypothermic Circulatory Arrest with Alpha-Stat and PH-Stat Cardiopulmonary Bypass in Newborn Pigs. J. Thorac. Cardiovasc. Surg. 2001, 121, 336–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, T.; Hatsuoka, S.; Stock, U.A.; Duebener, L.F.; Lidov, H.G.W.; Holmes, G.L.; Sperling, J.S.; Munakata, M.; Laussen, P.C.; Jonas, R.A. Prediction of Safe Duration of Hypothermic Circulatory Arrest by Near-Infrared Spectroscopy. J. Thorac. Cardiovasc. Surg. 2001, 122, 339–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagino, I.; Anttila, V.; Zurakowski, D.; Duebener, L.F.; Lidov, H.G.W.; Jonas, R.A. Tissue Oxygenation Index Is a Useful Monitor of Histologic and Neurologic Outcome after Cardiopulmonary Bypass in Piglets. J. Thorac. Cardiovasc. Surg. 2005, 130, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Mavroudis, C.D.; Mensah-Brown, K.G.; Ko, T.S.; Boorady, T.W.; Massey, S.L.; Abend, N.S.; Nicolson, S.C.; Morgan, R.W.; Mascio, C.E.; Gaynor, J.W.; et al. Electroencephalographic Response to Deep Hypothermic Circulatory Arrest in Neonatal Swine and Humans. Ann. Thorac. Surg. 2018, 106, 1841–1846. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Eucker, S.a.; Durduran, T.; Yu, G.; Ralston, J.; Friess, S.H.; Ichord, R.N.; Margulies, S.S.; Yodh, A.G. Diffuse Optical Monitoring of Hemodynamic Changes in Piglet Brain with Closed Head Injury. J. Biomed. Opt. 2009, 14, 034015. [Google Scholar] [CrossRef]
- Fantini, S.; Hueber, D.; Franceschini, M.A.; Gratton, E.; Rosenfeld, W.; Stubblefield, P.G.; Maulik, D.; Stankovic, M.R. Non-Invasive Optical Monitoring of the Newborn Piglet Brain Using Continuous-Wave and Frequency-Domain Spectroscopy. Phys. Med. Biol. 1999, 44, 1543–1563. [Google Scholar] [CrossRef]
- Hueber, D.M.; Franceschini, M.A.; Ma, H.Y.; Zhang, Q.; Ballesteros, J.R.; Fantini, S.; Walace, D.; Ntziachristos, V.; Chance, B. Non-Invasive and Quantitative near-Infrared Haemoglobin Spectrometry in the Piglet Brain during Hypoxic Stress, Using a Frequency-Domain Multidistance Instrument. Phys. Med. Biol. 2001, 46, 41–62. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Katz, A.; Alfano, R.R.; Kofinas, A.D.; Kofinas, D.A.; Stubblefield, P.G.; Rosenfeld, W.; Beyer, D.; Maulik, D.; Stankovic, M.R. Brain Perfusion Monitoring with Frequency-Domain and Continuous-Wave near-Infrared Spectroscopy: A Cross-Correlation Study in Newborn Piglets. Phys. Med. Biol. 2000, 45, 3143–3158. [Google Scholar] [CrossRef]
- Kurth, C.D.; Levy, W.J.; McCann, J. Near-Infrared Spectroscopy Cerebral Oxygen Saturation Thresholds for Hypoxia–Ischemia in Piglets. J. Cereb. Blood Flow Metab. 2002, 22, 335–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ko, T.S.; Mavroudis, C.D.; Baker, W.B.; Morano, V.C.; Mensah-Brown, K.; Boorady, T.W.; Schmidt, A.L.; Lynch, J.M.; Busch, D.R.; Gentile, J.; et al. Non-Invasive Optical Neuromonitoring of the Temperature-Dependence of Cerebral Oxygen Metabolism during Deep Hypothermic Cardiopulmonary Bypass in Neonatal Swine. J. Cereb. Blood Flow Metab. 2020, 40, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Ko, T.S.; Mavroudis, C.D.; Morgan, R.W.; Baker, W.B.; Marquez, A.M.; Boorady, T.W.; Devarajan, M.; Lin, Y.; Roberts, A.L.; Landis, W.P.; et al. Non-Invasive Diffuse Optical Neuromonitoring during Cardiopulmonary Resuscitation Predicts Return of Spontaneous Circulation. Sci. Rep. 2021, 11, 3828. [Google Scholar] [CrossRef] [PubMed]
- Mavroudis, C.D.; Ko, T.S.; Morgan, R.W.; Volk, L.E.; Landis, W.P.; Smood, B.; Xiao, R.; Hefti, M.; Boorady, T.W.; Marquez, A.; et al. Epinephrine’s Effects on Cerebrovascular and Systemic Hemodynamics during Cardiopulmonary Resuscitation. Crit. Care 2020, 24, 583. [Google Scholar] [CrossRef]
- Patterson, M.S.; Moulton, J.D.; Wilson, B.C.; Berndt, K.W.; Lakowicz, J.R. Frequency-Domain Reflectance for the Determination of the Scattering and Absorption Properties of Tissue. Appl. Opt. 1991, 30, 4474–4476. [Google Scholar] [CrossRef]
- Fantini, S.; Franceschini, M.-A.A.; Maier, J.S.; Walker, S.A.; Barbieri, B.B.; Gratton, E. Frequency-Domain Multichannel Optical Detector for Noninvasive Tissue Spectroscopy and Oximetry. Opt. Eng. 1995, 34, 32. [Google Scholar] [CrossRef] [Green Version]
- Kreis, R.; Ernst, T.; Ross, B.D. Development of the Human Brain: In Vivo Quantification of Metabolite and Water Content with Proton Magnetic Resonance Spectroscopy. Magn. Reson. Med. 1993, 30, 424–437. [Google Scholar] [CrossRef]
- Durduran, T.; Choe, R.; Baker, W.B.; Yodh, A.G. Diffuse Optics for Tissue Monitoring and Tomography. Rep. Prog. Phys. 2010, 73, 076701. [Google Scholar] [CrossRef] [Green Version]
- Jain, V.; Buckley, E.M.; Licht, D.J.; Lynch, J.M.; Schwab, P.J.; Naim, M.Y.; Lavin, N.A.; Nicolson, S.C.; Montenegro, L.M.; Yodh, A.G.; et al. Cerebral Oxygen Metabolism in Neonates with Congenital Heart Disease Quantified by MRI and Optics. J. Cereb. Blood Flow Metab. 2014, 34, 380–388. [Google Scholar] [CrossRef] [Green Version]
- Irwin, D.; Dong, L.; Shang, Y.; Cheng, R.; Kudrimoti, M.; Stevens, S.D.; Yu, G. Influences of Tissue Absorption and Scattering on Diffuse Correlation Spectroscopy Blood Flow Measurements. Biomed. Opt. Express 2011, 2, 1969. [Google Scholar] [CrossRef] [Green Version]
- Hutchinson, P.J.; Jalloh, I.; Helmy, A.; Carpenter, K.L.H.; Rostami, E.; Bellander, B.M.; Boutelle, M.G.; Chen, J.W.; Claassen, J.; Dahyot-Fizelier, C.; et al. Consensus Statement from the 2014 International Microdialysis Forum. Intensive Care Med. 2015, 41, 1517–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hägglund, L.; Olivecrona, M.; Koskinen, L.O.D. Correlation of Cerebral and Subcutaneous Glycerol in Severe Traumatic Brain Injury and Association with Tissue Damage. Neurocrit. Care 2021, 36, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Marcoux, J.; McArthur, D.A.; Miller, C.; Glenn, T.C.; Villablanca, P.; Martin, N.A.; Hovda, D.A.; Alger, J.R.; Vespa, P.M. Persistent Metabolic Crisis as Measured by Elevated Cerebral Microdialysis Lactate-Pyruvate Ratio Predicts Chronic Frontal Lobe Brain Atrophy after Traumatic Brain Injury. Crit. Care Med. 2008, 36, 2871–2877. [Google Scholar] [CrossRef] [PubMed]
- Paraforou, T.; Paterakis, K.; Fountas, K.; Paraforos, G.; Chovas, A.; Tasiou, A.; Mpakopoulou, M.; Papadopoulos, D.; Karavellis, A.; Komnos, A. Cerebral Perfusion Pressure, Microdialysis Biochemistry and Clinical Outcome in Patients with Traumatic Brain Injury. BMC Res. Notes 2011, 4, 540. [Google Scholar] [CrossRef] [Green Version]
- Zeiler, F.A.; Thelin, E.P.; Helmy, A.; Czosnyka, M.; Hutchinson, P.J.A.; Menon, D.K. A Systematic Review of Cerebral Microdialysis and Outcomes in TBI: Relationships to Patient Functional Outcome, Neurophysiologic Measures, and Tissue Outcome. Acta Neurochir. 2017, 159, 2245–2273. [Google Scholar] [CrossRef] [Green Version]
- Nordström, C.H. Cerebral Energy Metabolism and Microdialysis in Neurocritical Care. Child’s Nerv. Syst. 2010, 26, 465–472. [Google Scholar] [CrossRef]
- Hillered, L.; Valtysson, J.; Enblad, P.; Persson, L. Interstitial Glycerol as a Marker for Membrane Phospholipid Degradation in the Acutely Injured Human Brain. J. Neurol. Neurosurg. Psychiatry 1998, 64, 486–491. [Google Scholar] [CrossRef]
- Merenda, A.; Gugliotta, M.; Holloway, R.; Levasseur, J.E.; Alessandri, B.; Sun, D.; Bullock, M.R. Validation of Brain Extracellular Glycerol as an Indicator of Cellular Membrane Damage Due to Free Radical Activity after Traumatic Brain Injury. J. Neurotrauma 2008, 25, 527–537. [Google Scholar] [CrossRef]
- Reinstrup, P.; Ståhl, N.; Mellergård, P.; Uski, T.; Ungerstedt, U.; Nordström, C.H. Intracerebral Microdialysis in Clinical Practice: Baseline Values for Chemical Markers during Wakefulness, Anesthesia, and Neurosurgery. Neurosurgery 2000, 47, 701–710. [Google Scholar] [CrossRef]
- Öner, M.; Kocakoç, D. JMASM 49: A Compilation of Some Popular Goodness of Fit Tests for Normal Distribution: Their Algorithms and MATLAB Codes (MATLAB). J. Mod. Appl. Stat. Methods 2017, 16, 30. [Google Scholar] [CrossRef]
- Hutchinson, P.J.A.; O’Connell, M.T.; Al-Rawi, P.G.; Kett-White, R.; Gupta, A.K.; Kirkpatrick, P.J.; Pickard, J.D. Clinical Cerebral Microdialysis-Determining the True Extracellular Concentration. Acta Neurochir. Suppl. 2002, 81, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, M.S.; Ainslie, P.N.; Griesdale, D.E. Clinical Pathophysiology of Hypoxic Ischemic Brain Injury after Cardiac Arrest: A “Two-Hit” Model. Crit. Care 2017, 21, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemyn, J.; Cascalho, A.; Goodchild, R.E. The Ins and Outs of Endoplasmic Reticulum-controlled Lipid Biosynthesis. EMBO Rep. 2017, 18, 1905–1921. [Google Scholar] [CrossRef] [PubMed]
- Bonvento, G.; Bolaños, J.P. Astrocyte-Neuron Metabolic Cooperation Shapes Brain Activity. Cell Metab. 2021, 33, 1546–1564. [Google Scholar] [CrossRef]
- Salazar, J.; Coleman, R.; Griffith, S.; McNeil, J.; Young, H.; Calhoon, J.; Serrano, F.; DiGeronimo, R. Brain Preservation with Selective Cerebral Perfusion for Operations Requiring Circulatory Arrest: Protection at 25 °C Is Similar to 18 °C with Shorter Operating Times. Eur. J. Cardio-Thorac. Surg. 2009, 36, 524–531. [Google Scholar] [CrossRef] [Green Version]
- Cavus, E.; Hoffmann, G.; Bein, B.; Scheewe, J.; Meybohm, P.; Renner, J.; Scholz, J.; Boening, A. Cerebral Metabolism during Deep Hypothermic Circulatory Arrest vs Moderate Hypothermic Selective Cerebral Perfusion in a Piglet Model: A Microdialysis Study. Paediatr. Anaesth. 2009, 19, 770–778. [Google Scholar] [CrossRef]
- Jonsson, O.; Myrdal, G.; Zemgulis, V.; Valtysson, J.; Hillered, L.; Thelin, S. Selective Antegrade Cerebral Perfusion at Two Different Temperatures Compared to Hypothermic Circulatory Arrest—An Experimental Study in the Pig with Microdialysis. Interact. Cardiovasc. Thorac. Surg. 2009, 8, 647–653. [Google Scholar] [CrossRef] [Green Version]
- Liang, M.Y.; Tang, Z.X.; Chen, G.X.; Rong, J.; Yao, J.P.; Chen, Z.; Wu, Z.K. Is Selective Antegrade Cerebral Perfusion Superior to Retrograde Cerebral Perfusion for Brain Protection during Deep Hypothermic Circulatory Arrest? Metabolic Evidence from Microdialysis. Crit. Care Med. 2014, 42, e319–e328. [Google Scholar] [CrossRef]
- Ungerstedt, U.; Rostami, E. Microdialysis in Neurointensive Care. Curr. Pharm. Des. 2005, 10, 2145–2152. [Google Scholar] [CrossRef]
- Syková, E.; Nicholson, C. Diffusion in Brain Extracellular Space. Physiol. Rev. 2008, 88, 1277–1340. [Google Scholar] [CrossRef] [Green Version]
- Sumbria, R.K.; Klein, J.; Bickel, U. Acute Depression of Energy Metabolism after Microdialysis Probe Implantation Is Distinct from Ischemia-Induced Changes in Mouse Brain. Neurochem. Res. 2011, 36, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Ten, V.S.; Starkov, A. Hypoxic-Ischemic Injury in the Developing Brain: The Role of Reactive Oxygen Species Originating in Mitochondria. Neurol. Res. Int. 2012, 2012, 542976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henriksen, O.M.; Vestergaard, M.B.; Lindberg, U.; Aachmann-Andersen, N.J.; Lisbjerg, K.; Christensen, S.J.; Rasmussen, P.; Olsen, N.V.; Forman, J.L.; Larsson, H.B.W.; et al. Interindividual and Regional Relationship between Cerebral Blood Flow and Glucose Metabolism in the Resting Brain. J. Appl. Physiol. 2018, 125, 1080–1089. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Timepoint | Parameter | DHCA (n = 7) | DHCBP (n = 7) | SCP (n = 6) |
|---|---|---|---|---|
| Normothermic Bypass | ICT, °C | 36.5 [35.6, 36.6] * | 34.2 [30.3, 35.2] * | 35.8 [34.6, 36.4] |
| StO2, % | 54.7 [51.9, 58.4] | 56.3 [52.7, 57.7] | 61.4 [59.5, 63.2] | |
| rTHC, % Baseline | 95.5 [89.5, 96.1] † | 98.5 [85.7, 100.5] | 102.9 [99.2, 106.5] † | |
| rBFI, % Baseline | 106.6 [102.8, 133.9] | 115.2 [105.0, 160.1] | 104.1 [88.8, 125.1] | |
| LPR | 21.2 [10.0, 41.8] | 13.8 [0.7, 22.0] | 11.9 [10.3, 15.3] | |
| Glycerol, µmol/L | 32.3 [18.6, 47.6] † | 21.8 [17.1, 27.9] | 13.3 [11.4, 20.3] † | |
| Cooling | ICT, °C | 26.4 [24.2, 29.4] | 29.2 [25.8, 31.4] | 22.6 [21.5, 25.3] |
| StO2, % | 66.1 [63.7, 69.9] | 59.5 [57.8, 63.6] | 66.8 [61.8, 66.8] | |
| rTHC, % Baseline | 101.1 [99.1, 105.5] | 101.6 [97.2, 106.7] | 105.1 [101.5, 111.5] | |
| rBFI, % Baseline | 63.3 [57.2, 66.2] | 89.5 [65.6, 96.6] | 41.0 [17.9, 70.3] | |
| LPR | 25.3 [21.5, 34.1] | 17.5 [5.5, 34.2] | 17.5 [14.0, 25.4] | |
| Glycerol, µmol/L | 25.8 [13.8, 45.0] † | 13.8 [12.3, 15.8] | 11.7 [10.9, 13.3] † | |
| Deep Hypothermia | ICT, °C | 19.9 [19.0, 20.8] | 19.4 [18.9, 21.1] | 20.4 [19.7, 23.3] |
| StO2, % | 36.3 [26.9, 45.2] *,† | 66.7 [54.2, 74.3] * | 65.5 [58.2, 67.0] † | |
| rTHC, % Baseline | 84.2 [72.6, 86.3] *,† | 106.8 [87.9, 117.1] * | 99.7 [94.7, 109.0] † | |
| rBFI, % Baseline | 1.8 [1.3, 8.2] *,† | 43.8 [22.6, 70.3] * | 23.4 [17.3, 35.4] † | |
| LPR | 108.9 [52.7, 172.3] * | 7.2 [1.3, 13.5] * | 13.9 [8.2, 27.7] | |
| Glycerol, µmol/L | 27.4 [18.2, 40.2] | 11.3 [8.3, 19.9] | 13.8 [11.9, 18.0] | |
| Rewarming | ICT, °C | 27.9 [25.7, 29.8] | 26.8 [22.8, 31.6] | 30.0 [27.0, 31.8] |
| StO2, % | 54.3 [48.3, 54.6] *,† | 60.7 [56.5, 62.6] * | 59.4 [58.3, 61.8] † | |
| rTHC, % Baseline | 101.0 [93.5, 103.6] | 112.5 [101.4, 120.2] | 107.8 [98.8, 118.3] | |
| rBFI, % Baseline | 58.0 [43.9, 78.8] | 63.8 [40.3, 71.9] | 55.2 [45.8, 61.3] | |
| LPR | 38.3 [24.0, 59.1] | 7.1 [4.7, 22.6] | 14.7 [5.2, 21.1] | |
| Glycerol, µmol/L | 61.3 [45.8, 78.4] * | 17.6 [11.6, 26.2] * | 24.1 [22.3, 26.9] |
| Parameter | Odds Ratio [95% CI] | p-Value | P(x) = 0.5 Threshold |
|---|---|---|---|
| StO2 (%) | 0.86 [0.76, 0.97] | 0.010 | 48.2 |
| rTHC (% Baseline) | 0.89 [0.81, 0.98] | 0.010 | 91.0 |
| rCBF (% Baseline) | 0.94 [0.90, 0.98] | 0.003 | 39.0 |
| Parameter | Odds Ratio [95% CI] | p-Value | P(x) = 0.5 Threshold |
|---|---|---|---|
| StO2 (%) | 0.91 [0.86, 0.96] | <0.001 | 47.8 |
| rTHC (% Baseline) | 0.90 [0.85, 0.95] | <0.001 | 88.0 |
| rCBF (% Baseline) | 0.98 [0.97, 1.00] | 0.007 | 9.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, T.S.; Mavroudis, C.D.; Benson, E.J.; Forti, R.M.; Melchior, R.W.; Boorady, T.W.; Morano, V.C.; Mensah-Brown, K.; Lin, Y.; Aronowitz, D.; et al. Correlation of Cerebral Microdialysis with Non-Invasive Diffuse Optical Cerebral Hemodynamic Monitoring during Deep Hypothermic Cardiopulmonary Bypass. Metabolites 2022, 12, 737. https://doi.org/10.3390/metabo12080737
Ko TS, Mavroudis CD, Benson EJ, Forti RM, Melchior RW, Boorady TW, Morano VC, Mensah-Brown K, Lin Y, Aronowitz D, et al. Correlation of Cerebral Microdialysis with Non-Invasive Diffuse Optical Cerebral Hemodynamic Monitoring during Deep Hypothermic Cardiopulmonary Bypass. Metabolites. 2022; 12(8):737. https://doi.org/10.3390/metabo12080737
Chicago/Turabian StyleKo, Tiffany S., Constantine D. Mavroudis, Emilie J. Benson, Rodrigo M. Forti, Richard W. Melchior, Timothy W. Boorady, Vincent C. Morano, Kobina Mensah-Brown, Yuxi Lin, Danielle Aronowitz, and et al. 2022. "Correlation of Cerebral Microdialysis with Non-Invasive Diffuse Optical Cerebral Hemodynamic Monitoring during Deep Hypothermic Cardiopulmonary Bypass" Metabolites 12, no. 8: 737. https://doi.org/10.3390/metabo12080737
APA StyleKo, T. S., Mavroudis, C. D., Benson, E. J., Forti, R. M., Melchior, R. W., Boorady, T. W., Morano, V. C., Mensah-Brown, K., Lin, Y., Aronowitz, D., Starr, J. P., Rosenthal, T. M., Shade, B. C., Schiavo, K. L., White, B. R., Lynch, J. M., Gaynor, J. W., Licht, D. J., Yodh, A. G., ... Kilbaugh, T. J. (2022). Correlation of Cerebral Microdialysis with Non-Invasive Diffuse Optical Cerebral Hemodynamic Monitoring during Deep Hypothermic Cardiopulmonary Bypass. Metabolites, 12(8), 737. https://doi.org/10.3390/metabo12080737