Comparative Analysis of Chloroplast Genomes of Dalbergia Species for Identification and Phylogenetic Analysis

 ,
, 
Abstract
:1. Introduction
2. Results
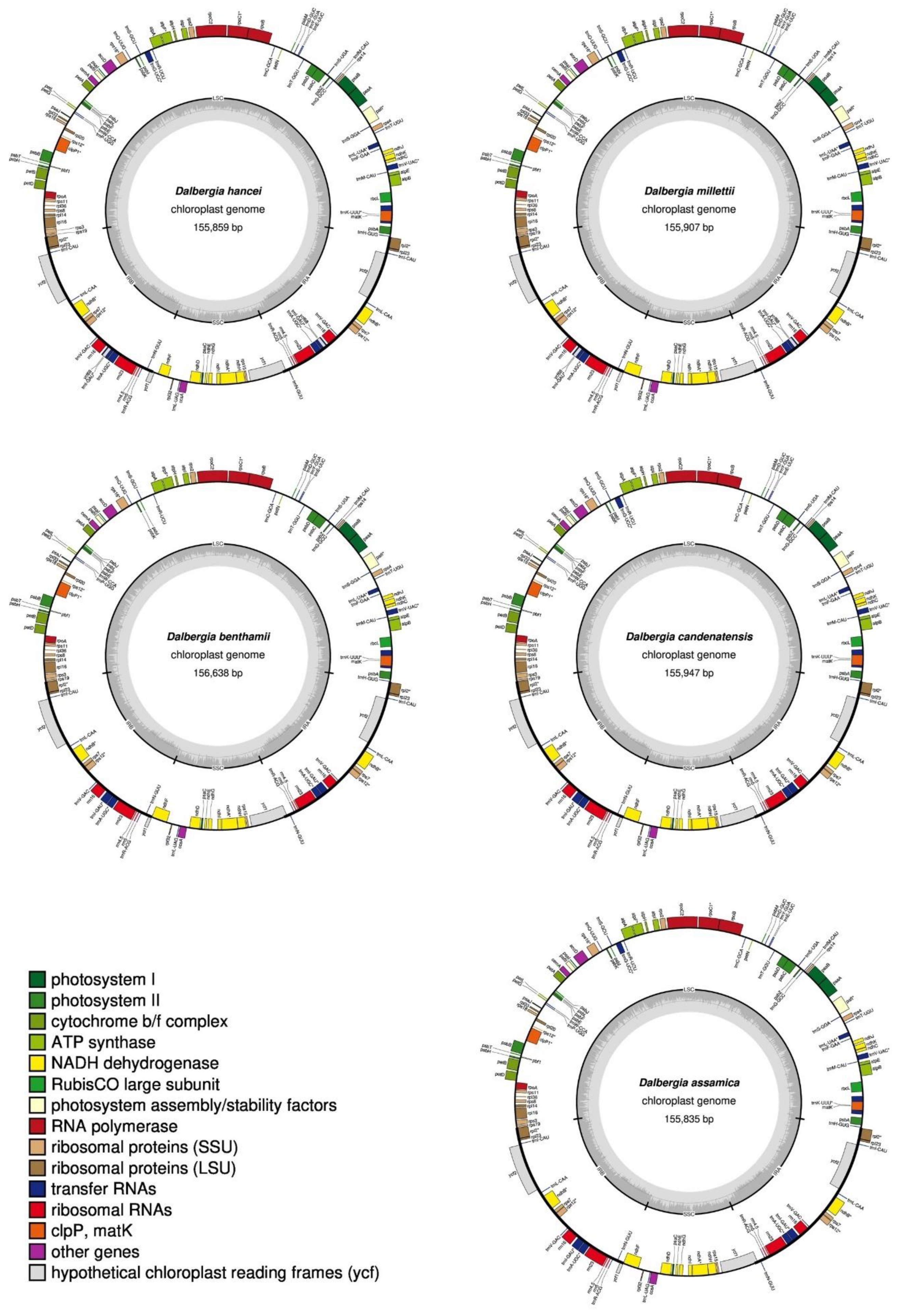
2.1. Genome Sequencing and Features of Assembled Chloroplast Genomes
2.2. Sequence Repeat Elements
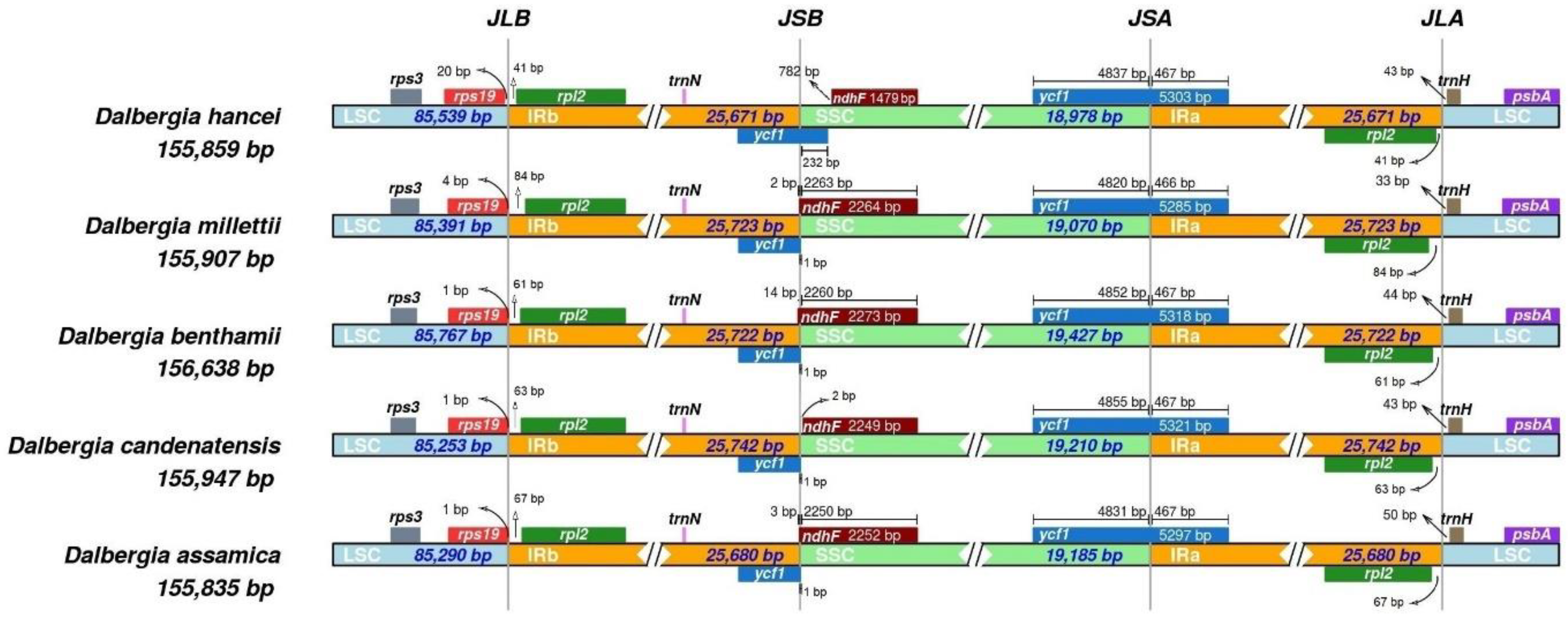
2.3. Comparative Genome Analysis
2.4. Detection of Potential MTPTs
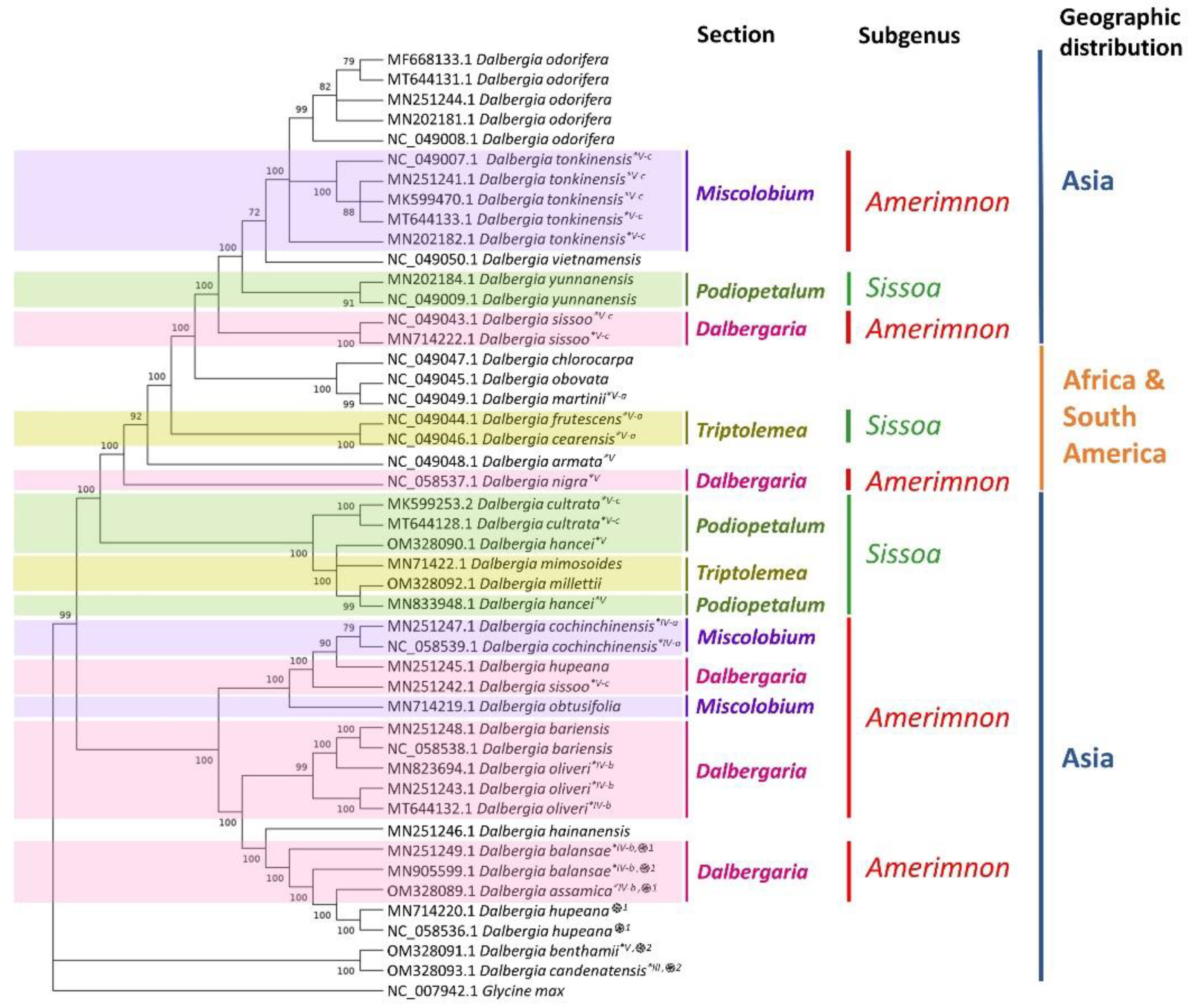
2.5. Phylogenetic Analysis
2.6. Candidate Markers for Identification of Medicinal Dalbergia Species
3. Discussion
4. Material and Methods
4.1. Ethics Statement
4.2. Plant Material and DNA Extraction
4.3. Chloroplast Genome Sequencing, Assembly and Annotation
4.4. Repeat Sequence Analysis
4.5. Comparative Genome Analysis
4.6. Identification Potential Mitochondrial Plastid DNAs (MTPTs)
4.7. Phylogenetic Analysis
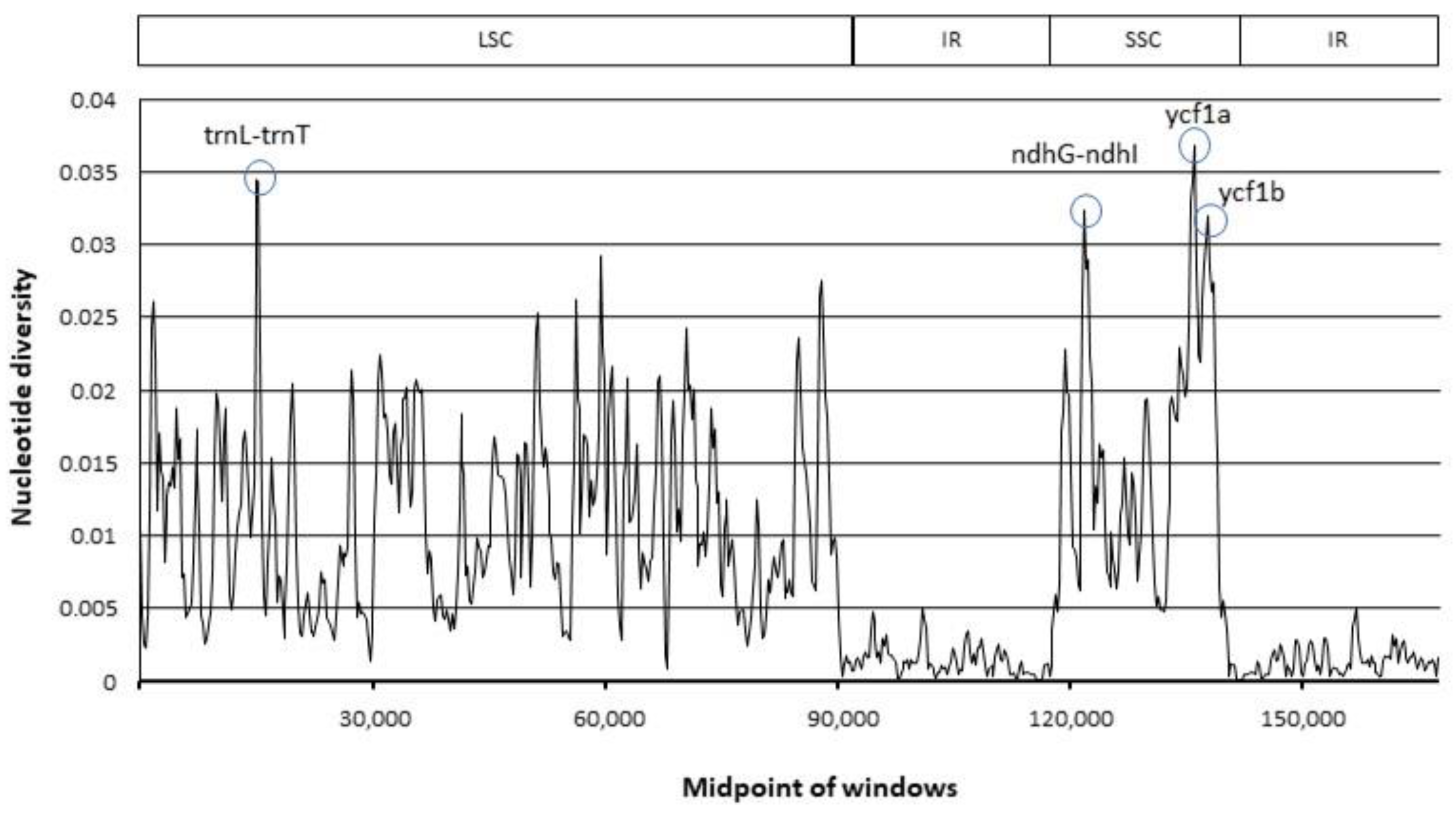
4.8. Evaluation of Divergence Hotspots and ITS2 Sequence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dalbergia L.f.|Plants of the World Online|Kew Science. Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:331542-2 (accessed on 21 March 2022).
- Legume|Dalbergia L.f. Available online: https://www.legumedata.org/taxonomy/taxon/755401 (accessed on 21 March 2022).
- Klitgaard, B.B.; Lavin, M. Tribe Dalbergieae. In Legumes of the World; Lewis, G.P., Schrire, B.D., Lock, M., Mackinder, B., Eds.; Royal Botanic Gardens, Kew: London, UK, 2005; pp. 307–335. [Google Scholar]
- Cardoso, D.; de Queiroz, L.P.; Pennington, R.T.; de Lima, H.C.; Fonty, E.; Wojciechowski, M.F.; Lavin, M. Revisiting the phylogeny of papilionoid legumes: New insights from comprehensively sampled early-branching lineages. Am. J. Bot. 2012, 99, 1991–2013. [Google Scholar] [CrossRef] [PubMed]
- National Standard of Hongmu (GB/T 18107-2017). Available online: http://openstd.samr.gov.cn/bzgk/gb/newGbInfo?hcno=6E961C6DB78254EF883B5053D08BFA3B (accessed on 21 March 2022).
- Royal Botanic Gardens, Kew: Medicinal Plant Names Services. Available online: https://mpns.science.kew.org/mpns-portal/ (accessed on 21 March 2022).
- Monograph of Dalbergiae Oderiferae Lignum. In Pharmacopoeia of the People’s Republic of China, 2020 ed.; China Medical Science and Technology Press: Beijing, China, 2020; Volume I, p. 240.
- Zhao, X.; Wang, C.; Meng, H.; Yu, Z.; Yang, M.; Wei, J. Dalbergia odorifera: A review of its traditional uses, phytochemistry, pharmacology, and quality control. J. Ethnopharmacol. 2020, 248, 112328. [Google Scholar] [CrossRef] [PubMed]
- Zhonghuabencao; Shanghai Science and Technology Press: Shanghai, China, 1997; Volume 4, pp. 432–441.
- Wheeler, E.A. InsideWood—A web resource for hardwood anatomy. IAWA J. 2011, 32, 199–211. [Google Scholar] [CrossRef]
- Koch, G.; Richter, H.; Schmitt, U. Design and application of CITESwoodID computer-aided identification and description of CITES-protected timbers. IAWA J. 2011, 32, 213–220. [Google Scholar] [CrossRef]
- de Palacios, P.; Esteban, L.G.; Gasson, P.; García-Fernández, F.; de Marco, A.; García-Iruela, A.; García-Esteban, L.; González-de-Vega, D. Using lenses attached to a smartphone as a macroscopic early warning tool in the illegal timber trade, in particular for CITES-listed species. Forests 2020, 11, 1147. [Google Scholar] [CrossRef]
- Rout, G.R.; Bhatacharya, D.; Nanda, R.M.; Nayak, S.; Das, P. Evaluation of genetic relationships in Dalbergia species using RAPD markers. Biodivers Conserv. 2003, 12, 197–206. [Google Scholar] [CrossRef]
- de Oliveira Buzatti, R.S.; Chicata, F.S.L.; Lovato, M.B. Transferability of microsatellite markers across six Dalbergia (Fabaceae) species and their characterization for Dalbergia miscolobium. Biochem. Syst. Ecol. 2016, 69, 161–165. [Google Scholar] [CrossRef]
- Vatanparast, M.; Klitgård, B.B.; Adema, F.A.C.B.; Pennington, R.T.; Yahara, T.; Kajita, T. First molecular phylogeny of the pantropical genus Dalbergia: Implications for infrageneric circumscription and biogeography. S. Afr. J. Bot. 2013, 89, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Hartvig, I.; Czako, M.; Kjær, E.D.; Nielsen, L.R.; Theilade, I. The use of DNA barcoding in identification and conservation of rosewood (Dalbergia spp.). PLoS ONE 2015, 10, e0138231. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wu, J.; Wang, Y.; Lian, X.; Wu, F.; Zhou, L.; Huan, Z.; Zhu, S. The phylogenetic analysis of Dalbergia (Fabaceae: Papilionaceae) based on different DNA barcodes. Holzforschung 2017, 71, 939–949. [Google Scholar] [CrossRef]
- Yu, M.; Jiao, L.; Guo, J.; Wiedenhoeft, A.C.; He, T.; Jiang, X.; Yin, Y. DNA barcoding of vouchered xylarium wood specimens of nine endangered Dalbergia species. Planta 2017, 246, 1165–1176. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Jiao, L.; Yu, G.J.; Jiang, X.; Yin, Y. DNA barcoding authentication for the wood of eight endangered Dalbergia timber species using machine learning approaches. Holzforschung 2018, 73, 277–285. [Google Scholar] [CrossRef]
- Hassold, S.; Lowry, P.P., II; Bauert, M.R.; Razafintsalama, A.; Ramamonjisoa, L.; Widmer, A. DNA Barcoding of Malagasy Rosewoods: Towards a Molecular Identification of CITES-Listed Dalbergia Species. PLoS ONE 2016, 11, e0157881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bentham, G. A Synopsis of the Dalbergieae, a tribe of the Leguminosae. J. Proc. Linn. Soc. Bot. 1860, 4, 1–134. [Google Scholar] [CrossRef]
- Prain, D. The species of Dalbergia of southeastern Asia. Ann. R. Bot. Gard. 1904, 10, 1–114. [Google Scholar]
- de Carvalho, A.M. A Synopsis of the Genus Dalbergia (Fabaceae: Dalbergieae) in Brazil. Brittonia 1997, 49, 87–109. [Google Scholar] [CrossRef]
- Thomas, W.W.; Amorim, A.M. André Maurício Vieira de Carvalho (1951–2002). Taxon 2003, 52, 147–148. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Henry, R.J.; Rossetto, M.; Wang, Y.; Chen, S. Plant DNA barcoding: From gene to genome. Biol. Rev. Camb. Philos. Soc. 2015, 90, 157–166. [Google Scholar] [CrossRef]
- Krawczyk, K.; Nobis, M.; Myszczyński, K.; Klichowska, E.; Sawicki, J. Plastid super-barcodes as a tool for species discrimination in feather grasses (Poaceae: Stipa). Sci. Rep. 2018, 8, 1924. [Google Scholar] [CrossRef] [Green Version]
- Kong, B.L.H.; Park, H.S.; Lau, T.W.D.; Lin, Z.; Yang, T.J.; Shaw, P.C. Comparative analysis and phylogenetic investigation of Hong Kong Ilex chloroplast genomes. Sci. Rep. 2021, 11, 5153. [Google Scholar] [CrossRef]
- Wu, L.; Wu, M.; Cui, N.; Xiang, L.; Li, X.; Chen, S. Plant super-barcode: A case study on genome-based identification for closely related species of Fritillaria. Chin. Med. 2021, 16, 52. [Google Scholar] [CrossRef] [PubMed]
- Yik, M.H.-Y.; Kong, B.L.-H.; Siu, T.-Y.; Lau, D.T.-W.; Cao, H.; Shaw, P.-C. Differentiation of Hedyotis diffusa and Common Adulterants Based on Chloroplast Genome Sequencing and DNA Barcoding Markers. Plants 2021, 10, 161. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, Y.; Xu, J.; Li, W.; Li, M. Characterization of the complete chloroplast genome sequence of Dalbergia species and its phylogenetic implications. Sci. Rep. 2019, 9, 20401. [Google Scholar] [CrossRef] [PubMed]
- Saski, C.; Lee, S.B.; Daniell, H.; Wood, T.C.; Tomkins, J.; Kim, H.G.; Jansen, R.K. Complete chloroplast genome sequence of Gycine max and comparative analyses with other legume genomes. Plant Mol. Biol. 2005, 59, 309–322. [Google Scholar] [CrossRef]
- Guo, X.; Castillo-Ramírez, S.; González, V.; Bustos, P.; Fernández-Vázquez, J.L.; Santamaría, R.I.; Arellano, J.; Cevallos, M.A.; Dávila, G. Rapid evolutionary change of common bean (Phaseolus vulgaris L) plastome, and the genomic diversification of legume chloroplasts. BMC Genom. 2007, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.E.; Rousseau-Gueutin, M.; Cordonnier, S.; Lima, O.; Michon-Coudouel, S.; Naquin, D.; de Carvalho, J.F.; Aïnouche, M.; Salmon, A.; Aïnouche, A. The first complete chloroplast genome of the Genistoid legume Lupinus luteus: Evidence for a novel major lineage-specific rearrangement and new insights regarding plastome evolution in the legume family. Ann. Bot. 2014, 113, 1197–1210. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J. Promiscuous DNA—Chloroplast genes inside plant mitochondria. Nature 1982, 299, 678–679. [Google Scholar] [CrossRef]
- Stern, D.B.; Lonsdale, D.M. Mitochondrial and chloroplast genomes of maize have a 12-kilobase DNA sequence in common. Nature 1982, 299, 698–702. [Google Scholar] [CrossRef]
- Smith, D.R. Extending the limited transfer window hypothesis to inter-organelle DNA migration. Genome Biol. Evol. 2011, 3, 743–748. [Google Scholar] [CrossRef] [Green Version]
- Sloan, D.B.; Wu, Z. History of plastid DNA insertions reveals weak deletion and at mutation biases in angiosperm mitochondrial genomes. Genome Biol. Evol. 2014, 6, 3210–3221. [Google Scholar] [CrossRef]
- Wang, X.-C.; Chen, H.; Yang, D.; Liu, C. Diversity of mitochondrial plastid DNAs (MTPTs) in seed plants. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2018, 29, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Jayakodi, M.; Lee, S.H.; Jeon, J.H.; Lee, H.O.; Park, J.Y.; Moon, B.C.; Kim, C.K.; Wing, R.A.; Newmaster, S.G.; et al. Mitochondrial plastid DNA can cause DNA barcoding paradox in plants. Sci. Rep. 2020, 10, 6112. [Google Scholar] [CrossRef] [Green Version]
- Keller, J.; Rousseau-Gueutin, M.; Martin, G.E.; Morice, J.; Boutte, J.; Coissac, E.; Ourari, M.; Aïnouche, M.; Salmon, A.; Cabello-Hurtado, F.; et al. The evolutionary fate of the chloroplast and nuclear rps16 genes as revealed through the sequencing and comparative analyses of four novel legume chloroplast genomes from Lupinus. DNA Res. 2017, 24, 343–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, J.J.; Doyle, J.L.; Ballenger, J.A.; Palmer, J.D. The distribution and phylogenetic significance of a 50-kb chloroplast DNA inversion in the flowering plant family Leguminosae. Mol. Phylogenet. Evol. 1996, 5, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, E.N.; Ruhlman, T.A.; Sabir, J.S.M.; Hajrah, N.H.; Alharbi, N.S.; Al-Malki, A.L.; Bailey, C.D.; Jansen, R.K. Plastid genome sequences of legumes reveal parallel inversions and multiple losses of rps16 in papilionoids. J. Syst. Evol. 2015, 53, 458–468. [Google Scholar] [CrossRef]
- Magee, A.M.; Aspinall, S.; Rice, D.W.; Cusack, B.P.; Semon, M.; Perry, A.S.; Stefanović, S.; Milbourne, D.; Barth, S.; Palmer, J.D.; et al. Localized hypermutation and associated gene losses in legume chloroplast genomes. Genome Res. 2010, 20, 1700–1710. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.; Doyle, J.L.; Palmer, J.D. Multiple independent losses of two genes and one intron from legume chloroplast genome. Syst. Bot. 1995, 20, 272–294. [Google Scholar] [CrossRef]
- Hiratsuka, J.; Shimada, H.; Whittier, R.; Ishibashi, T.; Sakamoto, M.; Mori, M.; Kondo, C.; Honji, Y.; Sun, C.R.; Meng, B.Y.; et al. The complete sequence of the rice (Oryza sativa) chloroplast genome: Intermolecular recombination between distinct tRNA genes accounts for a major plastid DNA inversion during the evolution of the cereals. Mol. Gen. Genet. 1989, 217, 185–194. [Google Scholar] [CrossRef]
- Stoebe, B.; Martin, W.; Kowallik, K.V. Distribution and nomenclature of protein-coding genes in 12 sequenced chloroplast genomes. Plant Mol. Biol. Report 1998, 16, 243–255. [Google Scholar] [CrossRef]
- Raubeson, L.A.; Peery, R.; Chumley, T.W.; Dziubek, C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Comparative chloroplast genomics: Analyses including new sequences from the angiosperms Nuphar advena and Ranunculus macranthus. BMC Genom. 2017, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- GCMTI RD-5:2020 Generating DNA Barcodes for Plant-Derived Chinese Materia Medica (CMM). Available online: https://www.cmro.gov.hk/html/eng/useful_information/gcmti/research/testing_methods/plant_derived.html# (accessed on 21 March 2022).
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The Manifold Applications of Repeat Analysis on a Genomic Scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Brudno, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Glocal Alignment: Finding Rear-rangements During Alignment. Bioinformatics 2003, 19, i54–i62. [Google Scholar] [CrossRef] [Green Version]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Qin, M.; Zhu, C.-J.; Yang, J.-B.; Vatanparast, M.; Schley, R.; Lai, Q.; Zhang, D.-Y.; Tu, T.-Y.; Klitgard, B.B.; Li, S.-J.; et al. Comparative analysis of complete plastid genome reveals powerful barcode regions for identifying wood of Dalbergia odorifera and D. tonkinensis (Leguminosae). J. Syst. Evol. 2020, 60, 73–84. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GenBank Accession | Genome Size (bp) | LSC (bp) | IR (bp) | SSC (bp) | Total Gene Number | Protein Coding | Hypothetical Protein | tRNA | rRNA | GC% | A% | C% | G% | T% |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dalbergia hancei | OM328090 | 155,859 | 85,539 | 25,671 | 18,978 | 114 | 75 | 3 | 32 | 4 | 36.18 | 31.90 | 17.95 | 18.23 | 31.92 |
| Dalbergia millettii | OM328092 | 155,907 | 85,391 | 25,723 | 19,070 | 112 | 75 | 3 | 30 | 4 | 36.14 | 31.94 | 17.93 | 18.21 | 31.92 |
| Dalbergia benthamii | OM328091 | 156,638 | 85,767 | 25,722 | 19,427 | 112 | 75 | 2 | 31 | 4 | 36.02 | 31.99 | 17.88 | 18.14 | 31.99 |
| Dalbergia candenatensis | OM328093 | 155,947 | 85,253 | 25,742 | 19,210 | 111 | 75 | 2 | 30 | 4 | 36.07 | 31.96 | 17.89 | 18.17 | 31.97 |
| Dalbergia assamica | OM328089 | 155,835 | 85,290 | 25,680 | 19,185 | 111 | 75 | 2 | 30 | 4 | 36.19 | 31.92 | 17.96 | 18.23 | 31.89 |
| Gene Category | Gene Function | Gene Name |
|---|---|---|
| Photosynthesis-related genes | Rubisco | rbcL |
| Photosystem I | psaA, psaB, psaC, psaI, psaJ | |
| Assembly/stability of photosystem I | pafI **, pafII, pbf1 | |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbH, psbI, psbJ, psbK, psbL, psbM, psbT, psbZ | |
| ATP synthase | atpA, atpB, atpE, atpF *, atpH, atpI | |
| Cytochrome b/f complex | petA, petB *, petD, petG, petL, petN | |
| Cytochrome c synthesis | ccsA | |
| NADPH dehydrogenase | ndhA *, ndhB * (×2), ndhC, ndhD, ndhE, ndhF a, ndhG, ndhH, ndhI, ndhJ, ndhK | |
| Transcription- and translation-related genes | Transcription | rpoA, rpoB, rpoC1 *, rpoC2 |
| Ribosomal protein | rpl2 * (×2), rpl14, rpl16 *, rpl20, rpl23 (×2), rpl32, rpl33, rpl36, rps2, rps3, rps4, rps7 (×2), rps8, rps11, rps12 ** (×2, tran-spliced), rps14, rps15, rps16 *, rps18, rps19 | |
| RNA genes | Ribosomal RNA | rrn4.5 (×2), rrn5 (×2), rrn16 (×2), rrn23 (×2) |
| Transfer RNA | trnA-UGC * (×2), trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC, trnG-UCC *b, trnH-GUG, trnI-CAU (×2), trnI-GAU * (×2), trnK-UUU *, trnL-CAA (×2), trnL-UAA *, trnL-UAG, trnM-CAU, trnN-GUU (×2), trnP-UGG, trnQ-UUG, trnR-ACG (×2), trnR-UCU, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC (×2), trnV-UAC *, trnW-CCA, trnY-GUA | |
| Miscellaneous group | Maturase | matK |
| Inner membrane protein | cemA | |
| ATP-dependent protease | clpP1 ** | |
| Acetyl-CoA carboxylase | accD | |
| Unknown functions | ycf1 (×2) c, ycf2 (×2), ycf68 (×2) d |
| Species | Number of MTPTs (Range of Length of MTPTs) | Total Length & Percentage of MTPTs | ||||
|---|---|---|---|---|---|---|
| Total | Protein-Coding | Transfer RNA | Ribosomal RNA | Non-Coding | ||
| Dalbergia hancei |
45 (50–2493 bp) | 12 (50–2493 bp) | 9 (54–276 bp) | 22 (51-1022 bp) | 2 (76–202 bp) | 16,376 bp (10.51%) |
| Dalbergia millettii |
44 (50–2493 bp) | 12 (50–2493 bp) | 8 (54–86 bp) | 22 (51–1022 bp) | 2 (76–278 bp) | 16,176 bp (10.38%) |
| Dalbergia benthamii |
44 (50–2493 bp) | 11 (86–2493 bp) | 6 (54–81 bp) | 22 (51–1020 bp) | 5 (50–279 bp) | 15,636 bp (9.98%) |
| Dalbergia candenatensis |
44 (50–2493 bp) | 11 (86–2493 bp) | 6 (54–86 bp) | 22 (51–1021 bp) | 5 (50–281 bp) | 15,640 bp (10.03%) |
| Dalbergia assamica |
44 (50–2493 bp) | 12 (50–2493 bp) | 8 (54–86 bp) | 22 (51–1022 bp | 2 (76–293 bp) | 15,654 bp (10.05%) |
| Dalbergia odorifera (NC_049008.1) |
44 (50–2493 bp) | 10 (50–2493 bp) | 11 (55–193 bp) | 21 (51–1022 bp) | 2 (109–193 bp) | 15,670 bp (10.04%) |
| Species | Specimen Voucher No. | Collector No. | Inventory No. |
|---|---|---|---|
| Dalbergia hancei | D.T.W. Lau 240 | D.T.W. Lau 240 | CUSLSH2130 |
| Dalbergia candenatensis | R & E 016 | Rare and endangered plants 016 | HK0051279 |
| Dalbergia assamica | R & E 020 | Rare and endangered plants 020 | HK0051283 |
| Dalbergia millettii | T.Y. Siu 580 | T.Y. Siu 580 | CUSLSH2682 |
| Dalbergia benthamii | S.K. Tsang 070 | S.K. Tsang 070 | CUSLSH1909 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.-Y.; Wong, K.-H.; Kong, B.L.-H.; Siu, T.-Y.; But, G.W.-C.; Tsang, S.S.-K.; Lau, D.T.-W.; Shaw, P.-C. Comparative Analysis of Chloroplast Genomes of Dalbergia Species for Identification and Phylogenetic Analysis. Plants 2022, 11, 1109. https://doi.org/10.3390/plants11091109
Wu H-Y, Wong K-H, Kong BL-H, Siu T-Y, But GW-C, Tsang SS-K, Lau DT-W, Shaw P-C. Comparative Analysis of Chloroplast Genomes of Dalbergia Species for Identification and Phylogenetic Analysis. Plants. 2022; 11(9):1109. https://doi.org/10.3390/plants11091109
Chicago/Turabian StyleWu, Hoi-Yan, Kwan-Ho Wong, Bobby Lim-Ho Kong, Tin-Yan Siu, Grace Wing-Chiu But, Stacey Shun-Kei Tsang, David Tai-Wai Lau, and Pang-Chui Shaw. 2022. "Comparative Analysis of Chloroplast Genomes of Dalbergia Species for Identification and Phylogenetic Analysis" Plants 11, no. 9: 1109. https://doi.org/10.3390/plants11091109
APA StyleWu, H. -Y., Wong, K. -H., Kong, B. L. -H., Siu, T. -Y., But, G. W. -C., Tsang, S. S. -K., Lau, D. T. -W., & Shaw, P. -C. (2022). Comparative Analysis of Chloroplast Genomes of Dalbergia Species for Identification and Phylogenetic Analysis. Plants, 11(9), 1109. https://doi.org/10.3390/plants11091109