
New Strategies to Increase the Abiotic Stress Tolerance in Woody Ornamental Plants in Mediterranean Climate

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Methodology and Literature Research
3. Ornamental Shrubby Plants in the Urban Environment
4. Mechanism of Tolerance and/or Resistance of Ornamental Shrubs to Abiotic Stress
5. Strategies to Improve the Shrub Tolerance to Abiotic Stress
5.1. Plant Species and/or Cultivar Choice
5.2. Agronomic Tools and Management Plans
5.2.1. Biostimulants and Arbuscular Mycorrhizas
5.2.2. Mulching
5.2.3. Association among Different Species
5.2.4. Transplanting Modalities
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Götmark, F.; Götmark, E.; Jensen, A.M. Why be a shrub? A basic model and hypotheses for the adaptive values of a common growth form. Front. Plant Sci. 2016, 7, 1095. [Google Scholar] [CrossRef] [PubMed]
- Read, P.E.; Bavougian, C.M. Woody ornamentals. In Horticulture: Plants for People and Places; Dixon, G., Aldous, D., Eds.; Springer: Dordrecht, The Netherland, 2014; Volume 2, pp. 619–644. [Google Scholar]
- Van Laere, K.; Hokanson, S.C.; Contreras, R.; Van Huylenbroeck, J. Woody ornamentals of the temperate zone. In Ornamental Crops; Van Huylenbroeck, J., Ed.; Springer: Cham, Switzerland, 2018; Volume 11, pp. 803–887. [Google Scholar]
- Lists of Names of Woody Plants and Perennials. Available online: https://www.naktuinbouw.com/lists-names (accessed on 15 January 2023).
- Paz, S.; Negev, M.; Clermont, A.; Green, M.S. Health aspects of climate change in cities with Mediterranean climate, and local adaptation plans. Int. J. Environ. Res. Public Health 2016, 13, 438. [Google Scholar] [CrossRef] [PubMed]
- World Water Assessment Programme. The United Nations World Water Development Report 2014: Water and Energy; UNESCO: Paris, France, 2014; Available online: https://www.unwater.org/publications/un-world-water-development-report-2014 (accessed on 15 January 2023).
- Munné-Bosch, S.; Peñuelas, J. Drought-induced oxidative stress in strawberry tree (Arbutus unedo L.) growing in Mediterranean field conditions. Plant Sci. 2004, 166, 1105–1110. [Google Scholar] [CrossRef]
- Cassaniti, C.; Romano, D.; Hop, M.E.C.M.; Flowers, T.J. Growing floricultural crops with brackish water. Environ. Exp. Bot. 2013, 92, 165–175. [Google Scholar] [CrossRef]
- Filella, I.; Llusia, J.; Piñol, J.; Peñuelas, J. Leaf gas exchange and fluorescence of Phillyrea latifolia, Pistacia lentiscus and Quercus ilex saplings in severe drought and high temperature conditions. J. Exp. Bot. 1998, 39, 213–220. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Heywood, V.; Skoula, M. The MEDUSA Network: Conservation and sustainable use of wild plants of the Mediterranean region. In Perspectives on New Crops and New Uses; Janick, J., Ed.; ASHS Press: Alexandria, VA, USA, 1999; pp. 148–151. [Google Scholar]
- Médail, F.; Quézel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Givnish, T.J. Leaf and Canopy Adaptations in Tropical Forests. In Physiological Ecology of Plants of the Wet Tropics: Tasks for Vegetation Science; Medina, E., Mooney, H.A., Vázquez-Yánes, C., Eds.; Springer: Dordrecht, The Netherlands, 1984; Volume 12, pp. 51–84. [Google Scholar] [CrossRef]
- Francini, A.; Romano, D.; Toscano, S.; Ferrante, A. The contribution of ornamental plants to urban ecosystem services. Earth 2022, 3, 1258–1274. [Google Scholar] [CrossRef]
- Romano, D.; Scariot, V. Woody ornamentals: A review of genetic resources in the Mediterranean area. Acta Hortic. 2021, 1331, 325–334. [Google Scholar] [CrossRef]
- McDonald, A.G.; Bealey, W.J.; Fowler, D.; Dragosits, U.; Skiba, U.; Smith, R.I.; Nemitz, E. Quantifying the effect of urban tree planting on concentrations and depositions of PM10 in two UK conurbations. Atmos. Environ. 2007, 41, 8455–8467. [Google Scholar] [CrossRef]
- Massetti, L.; Petralli, M.; Napoli, M.; Brandani, G.; Orlandini, S.; Pearlmutter, D. Effects of deciduous shade trees on surface temperature and pedestrian thermal stress during summer and autumn. Int. J. Biometeorol. 2019, 63, 467–479. [Google Scholar] [CrossRef]
- Lopes, H.S.; Remoaldo, P.C.; Ribeiro, V.; Martin-Vide, J. Pathways for adapting tourism to climate change in an urban destination–Evidences based on thermal conditions for the Porto Metropolitan Area (Portugal). J. Environ. Manag. 2022, 315, 115161. [Google Scholar] [CrossRef]
- Lopes, H.S.; Remoaldo, P.C.; Ribeiro, V.; Martín-Vide, J. A comprehensive methodology for assessing outdoor thermal comfort in touristic city of Porto (Portugal). Urban Clim. 2022, 45, 101264. [Google Scholar] [CrossRef]
- Ilyas, M.; Liu, Y.Y.; Shah, S.; Ali, A.; Khan, A.H.; Zaman, F.; Zhang, Y.; Saud, S.; Adnan, M.; Wang, Y.J.; et al. Adaptation of functional traits and their plasticity of three ornamental trees growing in urban environment. Sci. Hortic. 2021, 286, 110248. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Khan, E.A. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants. Plant Physiol. Biochem. 2018, 135, 295–303. [Google Scholar] [CrossRef]
- Iqbal, M.S.; Singh, A.K.; Ansari, M.I. Effect of drought stress on crop production. In New Frontiers in Stress Management for Durable Agriculture; Rakshit, A., Singh, H.B., Singh, A.K., Singh, U.S., Fraceto, L., Eds.; Springer: Singapore, 2020; pp. 35–47. [Google Scholar]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Levitt, J. Responses of Plants to Environmental Stresses. Volume II: Water, Radiation, Salt, and Other Stresses; Academic Press: New York, NY, USA, 1980; p. 607. [Google Scholar]
- Kozlowski, T.T.; Pallardy, S.G. Acclimation and adaptive responses of woody plants to environmental stresses. Bot. Rev. 2002, 68, 270–334. [Google Scholar] [CrossRef]
- Vinod, K.K. Stress in plantation crops: Adaptation and management. In Crop Stress and Its Management: Perspectives and Strategies; Venkateswarlu, B., Shanker, A.K., Shanker, C., Maheswari, M., Eds.; Springer: Dordrecht, The Netherland, 2012; pp. 45–137. [Google Scholar]
- Chen, J.W.; Zhang, Q.; Li, X.S.; Cao, K.F. Independence of stem and leaf hydraulic traits in six Euphorbiaceae tree species with contrasting leaf phenology. Planta 2009, 230, 459–468. [Google Scholar] [CrossRef]
- Medrano, H.; Flexas, J.; Galmés, J. Variability in water use efficiency at the leaf level among Mediterranean plants with different growth forms. Plant Soil 2009, 317, 17–29. [Google Scholar] [CrossRef]
- Álvarez, S.; Rodríguez, P.; Broetto, F.; Sánchez-Blanco, M.J. Long term responses and adaptive strategies of Pistacia lentiscus under moderate and severe deficit irrigation and salinity: Osmotic and elastic adjustment, growth, ion uptake and photosynthetic activity. Agric. Water Manag. 2018, 202, 253–262. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Romano, D. Response of Mediterranean ornamental plants to drought stress. Horticulturae 2019, 5, 6. [Google Scholar] [CrossRef]
- Riaz, A.; Younis, A.; Taj, A.R.; Karim, A.; Tariq, U.; Munir, S.; Riaz, S. Effect of drought stress on growth and flowering of marigold (Tagetes erecta L.). Pak. J. Bot. 2013, 45 (Suppl. S1), 123–131. [Google Scholar]
- Galmés, J.; Medrano, H.; Flexas, J. Photosynthetic limitations in response to water stress and recovery in Mediterranean plants with different growth forms. New Phytol. 2007, 175, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Toscano, S.; Ferrante, A.; Tribulato, A.; Romano, D. Leaf physiological and anatomical responses of Lantana and Ligustrum species under different water availability. Plant Physiol. Biochem. 2018, 127, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.F. Shrub stems: Form and function. In Plant Stems Physiology and Functional Morphology; Gartner, B.L., Ed.; Academic Press: Cambridge, MA, USA, 1995; pp. 91–102. [Google Scholar] [CrossRef]
- Iqbal, A.; Fahad, S.; Iqbal, M.; Alamzeb, M.; Ahmad, A.; Anwar, S.; Khan, A.A.; Arif, M.; Saeed, M.; Song, M. Special adaptive features of plant species in response to drought. In Salt and Drought Stress Tolerance in Plants: Signaling and Communication in Plants; Hasanuzzaman, M., Tanveer, M., Eds.; Springer: Cham, Switzerland, 2020; pp. 77–118. [Google Scholar] [CrossRef]
- Vilagrosa, A.; Hernández, E.I.; Luis, V.C.; Cochard, H.; Pausas, J.G. Physiological differences explain the co-existence of different regeneration strategies in Mediterranean ecosystems. New Phytol. 2014, 201, 1277–1288. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü.; Keenan, T. Photosynthetic responses to stress in Mediterranean evergreens: Mechanisms and models. Environ. Exp. Bot. 2014, 103, 24–41. [Google Scholar] [CrossRef]
- Nadal, M.; Roig-Oliver, M.; Bota, J.; Flexas, J. Leaf age-dependent elastic adjustment and photosynthetic performance under drought stress in Arbutus unedo seedlings. Flora 2020, 271, 151662. [Google Scholar] [CrossRef]
- Flexas, J.; Diaz-Espejo, A.; Gago, J.; Gallé, A.; Galmés, J.; Gulías, J.; Medrano, H. Photosynthetic limitations in Mediterranean plants: A review. Environ. Exp. Bot. 2014, 103, 12–23. [Google Scholar] [CrossRef]
- Nadal, M.; Flexas, J. Variation in photosynthetic characteristics with growth form in a water-limited scenario: Implications for assimilation rates and water use efficiency in crops. Agric. Water Manag. 2019, 216, 457–472. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G. Morpho-anatomical traits for plant adaptation to drought. In Plant Responses to Drought Stress: From Morphological to Molecular Features; Aroca, R., Ed.; Springer: Berlin, Germany, 2012; pp. 37–61. [Google Scholar]
- Scoffoni, C.; Vuong, C.; Diep, S.; Cochard, H.; Sack, L. Leaf shrinkage with dehydration: Coordination with hydraulic vulnerability and drought tolerance. Plant Physiol. 2014, 164, 1772–1788. [Google Scholar] [CrossRef]
- Jafari, S.; Hashemi Garmdareh, S.E.; Azadegan, B. Effects of drought stress on morphological, physiological, and biochemical characteristics of stock plant (Matthiola incana L.). Sci. Hortic. 2019, 253, 128–133. [Google Scholar] [CrossRef]
- Geng, D.; Chen, P.; Shen, X.; Zhang, Y.; Li, X.; Jiang, L.; Xie, Y.; Niu, C.; Zhang, J.; Huang, X.; et al. MdMYB88 and MdMYB124 enhance drought tolerance by modulating root vessels and cell walls in apple. Plant Physiol. 2018, 178, 1296–1309. [Google Scholar] [CrossRef]
- Aspelmeier, S.; Leuschner, C. Genotypic variation in drought response of silver birch (Betula pendula Roth): Leaf and root morphology and carbon partitioning. Trees 2006, 20, 42–52. [Google Scholar] [CrossRef]
- Cai, X.; Starman, T.; Niu, G.; Hall, C.; Lombardini, L. Response of selected garden roses to drought stress. HortScience 2012, 47, 1050–1055. [Google Scholar] [CrossRef]
- Toscano, S.; Branca, F.; Romano, D.; Ferrante, A. An evaluation of different parameters to screen ornamental shrubs for salt spray tolerance. Biology 2020, 9, 250. [Google Scholar] [CrossRef]
- USEPA. Manual: Guidelines for Water Reuse; USEPA, Rep. 625/R-92/004; USEPA: Washington, DC, USA, 1992. Available online: https://www.epa.gov/sites/default/files/2019-08/documents/2004-guidelines-water-reuse.pdf (accessed on 10 February 2023).
- Cassaniti, C.; Romano, D.; Flowers, T.J. The response of ornamental plants to saline irrigation water. In Irrigation: Water Management, Pollution and Alternative Strategies; Garcia-Garizabal, I., Abrahao, R., Eds.; IntechOpen Limited: London, UK, 2012; pp. 131–158. [Google Scholar]
- Percival, G.C. Identification of foliar salt tolerance of woody perennials using chlorophyll fluorescence. HortScience 2005, 40, 1892–1897. [Google Scholar] [CrossRef]
- Appleton, B.; Huff, R.R.; French, S.C. Evaluating trees for saltwater spray tolerance for oceanfront sites. J. Arboric. 1999, 25, 205–210. [Google Scholar] [CrossRef]
- Tribulato, A.; Toscano, S.; Di Lorenzo, V.; Romano, D. Effects of water stress on gas exchange, water relations and leaf structure in two ornamental shrubs in the Mediterranean area. Agronomy 2019, 9, 381. [Google Scholar] [CrossRef]
- Azza Mazher, A.M.; Fatma El-Quesni, E.M.; Farahat, M.M. Responses of ornamental plants and woody trees to salinity. World J. Agric. Sci. 2007, 3, 386–395. [Google Scholar]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Cameron, R.W.F.; Harrison-Murray, R.S.; Scott, M.A. The use of controlled water stress to manipulate growth of container-grown Rhododendron cv. Hoppy. J. Hortic. Sci. Biotechnol. 1999, 74, 161–169. [Google Scholar] [CrossRef]
- Devitt, D.A.; Morris, R.L.; Fenstermaker, L.K. Foliar damage, spectral reflectance, and tissue ion concentrations of trees sprinkle irrigated with waters of similar salinity but different chemical composition. HortScience 2005, 40, 819–826. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Colmer, T.D.; Munns, R.; Flowers, T.J. Improving salt tolerance of wheat and barley: Future prospects. Aust. J. Exp. Agric. 2005, 45, 1425–1443. [Google Scholar] [CrossRef]
- Boursier, P.; Läuchli, A. Growth responses and mineral nutrient relations of salt stressed sorghum. Crop Sci. 1990, 30, 1226–1233. [Google Scholar] [CrossRef]
- Cassaniti, C.; Leonardi, C.; Flowers, T.J. The effect of sodium chloride on ornamental shrubs. Sci. Hortic. 2009, 122, 586–593. [Google Scholar] [CrossRef]
- Ferguson, L.; Grattan, S.R. How salinity damages citrus: Osmotic effects and specific ion toxicities. HortTechnology 2005, 15, 95–99. [Google Scholar] [CrossRef]
- Álvarez, S.; Sánchez-Blanco, M.J. Comparison of individual and combined effects of salinity and deficit irrigation on physiological, nutritional and ornamental aspects of tolerance in Callistemon laevis plants. J. Plant Physiol. 2015, 185, 65–74. [Google Scholar] [CrossRef]
- Sánchez-Blanco, M.J.; Álvarez, S.; Ortuño, M.F.; Ruiz-Sánchez, M.C. Root System Response to Drought and Salinity: Root Distribution and Water Transport. In Root Engineering: Soil Biology; Morte, A., Varma, A., Eds.; Springer: Berlin, Germany, 2014; Volume 40, pp. 325–352. [Google Scholar] [CrossRef]
- Álvarez, S.; Gómez-Bellot, M.J.; Castillo, M.; Banon, S.; Sánchez-Blanco, M.J. Osmotic and saline effect on growth, water relations, and ion uptake and translocation in Phlomis purpurea plants. Environ. Exp. Bot. 2012, 78, 138–145. [Google Scholar] [CrossRef]
- Fornes, F.; Belda, R.M.; Carrión, C.; Noguera, V.; García-Agustín, P.; Abad, M. Pre-conditioning ornamental plants to drought by means of saline water irrigation as related to salinity tolerance. Sci. Hortic. 2007, 113, 52–59. [Google Scholar] [CrossRef]
- Yang, J.L.; Zhang, G.L. Formation, characteristics and eco-environmental implications of urban soils—A review. Soil Sci. Plant Nutr. 2015, 61, 30–46. [Google Scholar] [CrossRef]
- Jackson, M.B. Hormones from roots as signals for the shoots of stressed plants. Trends Plant Sci. 1997, 2, 22–28. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Responses of woody plants to flooding and salinity. Tree Physiol. 1997, 17, 490. [Google Scholar] [CrossRef]
- Pezeshki, S.R. Wetland plant responses to soil flooding. Environ. Exp. Bot. 2001, 46, 299–312. [Google Scholar] [CrossRef]
- Folzer, H.; Dat, J.F.; Capelli, N.; Rieffel, D.; Badot, P.M. Response to flooding of sessile oak: An integrative study. Tree Physiol. 2006, 26, 759–766. [Google Scholar] [CrossRef]
- Else, M.A.; Coupland, D.; Dutton, L.; Jackson, M.B. Decreased root hydraulic conductivity reduces leaf water potential, initiates stomatal closure and slows leaf expansion in flooded plants of castor oil (Ricinus communis) despite diminished delivery of ABA from roots to shoots in xylem sap. Physiol. Plant. 2001, 111, 46–54. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Rennenberg, H. Molecular and physiological responses of trees to waterlogging stress. Plant Cell Environ. 2014, 37, 2245–2259. [Google Scholar] [CrossRef]
- Yamamoto, F.; Sakata, T.; Terazawa, K. Physiological, morphological and anatomical responses of Fraxinus mandshurica seedlings to flooding. Tree Physiol. 1995, 15, 713–719. [Google Scholar] [CrossRef]
- Parent, C.; Capelli, N.; Berger, A.; Crèvecoeur, M.; Dat, J.F. An overview of plant responses to soil waterlogging. Plant Stress 2008, 2, 20–27. [Google Scholar]
- Sauter, M. Root responses to flooding. Curr. Opin. Plant Biol. 2013, 16, 282–286. [Google Scholar] [CrossRef]
- Voesenek, L.A.; Bailey-Serres, J. Flood adaptive traits and processes: An overview. New Phytol. 2015, 206, 57–73. [Google Scholar] [CrossRef]
- Cameron, R.W.; King, C.M.; Robinson, S. Ornamental Mediterranean plants in the UK: Root adaptations to hypoxia and anoxia. Acta Hortic. 2010, 881, 469–474. [Google Scholar] [CrossRef]
- Yang, W.C.; Lin, K.H.; Wu, C.W.; Chang, Y.J.; Chang, Y.S. Effects of waterlogging with different water resources on plant growth and tolerance capacity of four herbaceous flowers in a bioretention basin. Water 2020, 12, 1619. [Google Scholar] [CrossRef]
- Asgari Lajayer, B.; Khadem Moghadam, N.; Maghsoodi, M.R.; Ghorbanpour, M.; Kariman, K. Phytoextraction of heavy metals from contaminated soil, water and atmosphere using ornamental plants: Mechanisms and efficiency improvement strategies. Environ. Sci. Pollut. Res. 2019, 26, 8468–8484. [Google Scholar] [CrossRef]
- Khan, A.H.A.; Kiyani, A.; Mirza, C.R.; Butt, T.A.; Barros, R.; Ali, B.; Iqbal, M.; Yousaf, S. Ornamental plants for the phytoremediation of heavy metals: Present knowledge and future perspectives. Environ. Res. 2021, 195, 110780. [Google Scholar] [CrossRef] [PubMed]
- Kneer, R.; Zenk, M.H. Phytochelatins protect plant enzymes from heavy metal poisoning. Phytochemistry 1992, 31, 2663–2667. [Google Scholar] [CrossRef]
- Cobbett, C.S. Phytochelatins and their roles in heavy metal detoxification. Plant Physiol. 2000, 123, 825–832. [Google Scholar] [CrossRef]
- Nazir, F.; Fariduddin, Q.; Khan, T.A. Hydrogen peroxide as a signalling molecule in plants and its crosstalk with other plant growth regulators under heavy metal stress. Chemosphere 2020, 252, 126486. [Google Scholar] [CrossRef]
- Middleton, L. Shade-tolerant flowering plants: Adaptations and horticultural implications. Acta Hortic. 2001, 552, 95–102. [Google Scholar] [CrossRef]
- Fonteno, W.C.; McWilliams, E.L. Light compensation points and acclimatization of four tropical foliage plants. J. Am. Soc. Hortic. Sci. 1978, 103, 52–56. [Google Scholar] [CrossRef]
- Zhao, D.; Hao, Z.; Tao, J. Effects of shade on plant growth and flower quality in the herbaceous peony (Paeonia lactiflora Pall.). Plant Physiol. Biochem. 2012, 61, 187–196. [Google Scholar] [CrossRef]
- Lugassi-Ben-Hamo, M.; Kitron, M.; Bustan, A.; Zaccai, M. Effect of shade regime on flower development, yield and quality in lisianthus. Sci. Hortic. 2010, 124, 248–253. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Ferrante, A. Molecular responses of vegetable, ornamental crops, and model plants to salinity stress. Int. J. Mol. Sci. 2023, 24, 3190. [Google Scholar] [CrossRef]
- Franco, J.A.; Martínez-Sánchez, J.J.; Fernández, J.A.; Bañón, S. Selection and nursery production of ornamental plants for landscaping and xerogardening in semi-arid environments. J. Hortic. Sci. Biotechnol. 2006, 81, 3–17. [Google Scholar] [CrossRef]
- Francini, A.; Toscano, S.; Romano, D.; Ferrini, F.; Ferrante, A. Biological contribution of ornamental plants for improving slope stability along urban and suburban areas. Horticulturae 2021, 7, 310. [Google Scholar] [CrossRef]
- Vaid, T.M.; Runkle, E.S. Developing flowering rate models in response to mean temperature for common annual ornamental crops. Sci. Hortic. 2013, 161, 15–23. [Google Scholar] [CrossRef]
- Khodorova, N.V.; Boitel-Conti, M. The role of temperature in the growth and flowering of geophytes. Plants 2013, 2, 699–711. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Vogt, J.; Hauer, R.J.; Fischer, B.C. The costs of maintaining and not maintaining the urban forest: A review of the urban forestry and arboriculture literature. Arboric. Urban For. 2015, 41, 293–323. [Google Scholar] [CrossRef]
- Núñez-Florez, R.; Pérez-Gómez, U.; Fernández-Méndez, F. Functional diversity criteria for selecting urban trees. Urban For. Urban Green. 2019, 38, 251–266. [Google Scholar] [CrossRef]
- Farrell, C.; Livesley, S.; Arndt, S.; Beaumont, L.; Burley, H.; Ellsworth, D.; Esperon-Rodriguez, M.; Fletcher, T.; Gallagher, R.; Ossola, A.; et al. Can we integrate ecological approaches to improve plant selection for green infrastructure? Urban For. Urban Green. 2022, 76, 127732. [Google Scholar] [CrossRef]
- Capotorti, G.; Bonacquisti, S.; Abis, L.; Aloisi, I.; Attorre, F.; Bacaro, G.; Balletto, G.; Banfi, E.; Barni, E.; Bartoli, F.; et al. More nature in the city. Plant Biosyst. 2020, 154, 1003–1006. [Google Scholar] [CrossRef]
- McPherson, E.G.; Berry, A.M.; van Doorn, N.S. Performance testing to identify climate-ready trees. Urban For. Urban Green. 2018, 29, 28–39. [Google Scholar] [CrossRef]
- Ferrante, A.; Toscano, S.; Romano, D.; Vagge, I. Physiological and morpho-anatomical traits used as markers for the selection of drought tolerance of ornamental plants. Acta Hortic. 2021, 1331, 253–260. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Romano, D. Can pedoclimatic needs be a driver for ornamental plant selection in Mediterranean area? Acta Hortic. 2022, 1345, 451–458. [Google Scholar] [CrossRef]
- Asgarzadeh, M.; Vahdati, K.; Lotfi, M.; Arab, M.; Babaei, A.; Naderi, F.; Soufi, M.P.; Rouhani, G. Plant selection method for urban landscapes of semi-arid cities (a case study of Tehran). Urban For. Urban Green. 2014, 13, 450–458. [Google Scholar] [CrossRef]
- Ghafari, S.; Kaviani, B.; Sedaghathoor, S.; Allahyari, M.S. Ecological potentials of trees, shrubs and hedge species for urban green spaces by multi criteria decision making. Urban For. Urban Green. 2020, 55, 126824. [Google Scholar] [CrossRef]
- Van der Veken, S.; Hermy, M.; Vellend, M.; Knapen, A.; Verheyen, K. Garden plants get a head start on climate change. Front. Ecol. Environ. 2008, 6, 212–216. [Google Scholar] [CrossRef]
- Miller, R.W. Urban Forestry: Planning and Managing Urban Greenspaces, 2nd ed.; Prentice-Hall: Hoboken, NJ, USA, 1997; p. 560. [Google Scholar]
- Roloff, A.; Korn, S.; Gillner, S. The Climate-Species-Matrix to select tree species for urban habitats considering climate change. Urban For. Urban Green. 2009, 8, 295–308. [Google Scholar] [CrossRef]
- Toscano, S.; Ferrante, A.; Romano, D.; Tribulato, A. Interactive effects of drought and saline aerosol stress on morphological and physiological characteristics of two ornamental shrub species. Horticulturae 2021, 7, 517. [Google Scholar] [CrossRef]
- Dale, A.G.; Frank, S.D. Urban warming trumps natural enemy regulation of herbivorous pests. Ecol. Appl. 2014, 24, 1596–1607. [Google Scholar] [CrossRef]
- Farieri, E.; Toscano, S.; Ferrante, A.; Romano, D. Identification of ornamental shrubs tolerant to saline aerosol for coastal urban and peri-urban greening. Urban For. Urban Green. 2016, 18, 9–18. [Google Scholar] [CrossRef]
- Santamour, F.S. Trees for urban planting: Diversity, uniformity, and common sense. In Proceedings of the Seventh Conference of the Metropolitan Tree Improvement Alliance (METRIA), The Morton Arboretum, Lisle, IL, USA, 11–12 June 1990; pp. 57–65. [Google Scholar]
- Kendal, D.; Dobbs, C.; Lohr, V.I. Global patterns of diversity in the urban forest: Is there evidence to support the 10/20/30 rule? Urban For. Urban Green. 2014, 13, 411–417. [Google Scholar] [CrossRef]
- Aronson, M.F.; Lepczyk, C.A.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S.; Vargo, T. Biodiversity in the city: Key challenges for urban green space management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systems. Italus Hortus 2018, 25, 27–36. [Google Scholar] [CrossRef]
- Koller, G.L. Transplanting stress—A view from the plant’s perspective. Arnoldia 1977, 37, 230–241. [Google Scholar]
- EC. REGULATION (EU) 2019/1009 of the European Parliament and of the Council of 5 June 2019 Laying down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003. 2019. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex%3A32019R1009 (accessed on 24 February 2023).
- La Torre, A.; Battaglia, V.; Caradonia, F. An overview of the current plant biostimulant legislations in different European Member States. J. Sci. Food Agric. 2016, 96, 727–734. [Google Scholar] [CrossRef]
- Newman, S.E.; Davies, F.T., Jr. High soil temperature and water relations of endomycorrhizal nursery crops. J. Environ. Hort. 1987, 5, 93–96. [Google Scholar] [CrossRef]
- de Vasconcelos, A.C.F.; Chaves, L.H.G. Biostimulants and their role in improving plant growth under abiotic stresses. In Biostimulants in Plant Science; Mirmajlessi, S.M., Radhakrishnan, R., Eds.; IntechOpen Limited: London, UK, 2020; pp. 3–16. [Google Scholar] [CrossRef]
- Massa, D.; Lenzi, A.; Montoneri, E.; Ginepro, M.; Prisa, D.; Burchi, G. Plant response to biowaste soluble hydrolysates in hibiscus grown under limiting nutrient availability. J. Plant Nutr. 2018, 41, 396–409. [Google Scholar] [CrossRef]
- Massa, D.; Prisa, D.; Montoneri, E.; Battaglini, D.; Ginepro, M.; Negre, M.; Burchi, G. Application of municipal biowaste derived products in Hibiscus cultivation: Effect on leaf gaseous exchange activity, and plant biomass accumulation and quality. Sci. Hortic. 2016, 205, 59–69. [Google Scholar] [CrossRef]
- Ferrante, A.; Trivellini, A.; Vernieri, P.; Piaggesi, A. Application of Actiwave® for improving the rooting of Camellia cuttings. Acta Hortic. 2013, 1009, 213–218. [Google Scholar] [CrossRef]
- Costa, G.; Labrousse, P.; Bodin, C.; Lhernould, S.; Carlué, M.; Krausz, P.; Authier, F. Effects of humic substances on the rooting and development of woody plant cuttings. Acta Hortic. 2008, 779, 255–262. [Google Scholar] [CrossRef]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Aalipour, H.; Nikbakht, A.; Ghasemi, M.; Amiri, R. Morpho-physiological and biochemical responses of two turfgrass species to arbuscular mycorrhizal fungi and humic acid under water stress condition. J. Soil Sci. Plant Nutr. 2020, 20, 566–576. [Google Scholar] [CrossRef]
- Papafotiou, M.; Kokotsakis, C.; Kavadia, A.; Ehaliotis, C. The effect of inoculation with arbuscular mycorrhizal fungi (AMF) and substrate type on growth and flowering of Gardenia jasminoides. Acta Hortic. 2021, 1327, 509–514. [Google Scholar] [CrossRef]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The combined effects of arbuscular mycorrhizal fungi (AMF) and lead (Pb) stress on Pb accumulation, plant growth parameters, photosynthesis, and antioxidant enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef]
- Larraburu, E.E.; Carletti, S.M.; Rodríguez Cáceres, E.A.; Llorente, B.E. Micropropagation of photinia employing rhizobacteria to promote root development. Plant Cell Rep. 2007, 26, 711–717. [Google Scholar] [CrossRef]
- Loconsole, D.; Sdao, A.E.; Cristiano, G.; De Lucia, B. Different responses to adventitious rhizogenesis under indole-3-butyric acid and seaweed extracts in ornamental’s cuttings: First results in Photinia x fraseri ‘Red Robin’. Agriculture 2023, 13, 513. [Google Scholar] [CrossRef]
- Rathore, S.S.; Chaudhary, D.R.; Boricha, G.N.; Ghosh, A.; Bhatt, B.P.; Zodape, S.T.; Patolia, J.S. Effect of seaweed extract on the growth, yield and nutrient uptake of soybean (Glycine max) under rainfed conditions. S. Afr. J. Bot. 2009, 75, 351–355. [Google Scholar] [CrossRef]
- Abd-Elkader, D.Y.; Mohamed, A.A.; Feleafel, M.N.; Al-Huqail, A.A.; Salem, M.Z.; Ali, H.M.; Hassan, H.S. Photosynthetic pigments and biochemical response of zucchini (Cucurbita pepo L.) to plant-derived extracts, microbial, and potassium silicate as biostimulants under greenhouse conditions. Front. Plant Sci. 2022, 13, 879545. [Google Scholar] [CrossRef]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Abd-Allah, E.F.; Egamberdieva, D. Arbuscular mycorrhizal fungi enhance basil tolerance to salt stress through improved physiological and nutritional status. Pak. J. Bot. 2016, 48, 37–45. [Google Scholar]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Kazemi, F.; Safari, N. Effect of mulches on some characteristics of a drought tolerant flowering plant for urban landscaping. Desert 2018, 23, 75–84. [Google Scholar] [CrossRef]
- Ferrini, F.; Fini, A.; Frangi, P.; Amoroso, G. Mulching of ornamental trees: Effects on growth and physiology. Arboric. Urban For. 2008, 34, 157. [Google Scholar] [CrossRef]
- Qu, B.; Liu, Y.; Sun, X.; Li, S.; Wang, X.; Xiong, K.; Yun, B.; Zhang, H. Effect of various mulches on soil physico—Chemical properties and tree growth (Sophora japonica) in urban tree pits. PLoS ONE 2019, 14, e0210777. [Google Scholar] [CrossRef]
- Ni, X.; Song, W.; Zhang, H.; Yang, X.; Wang, L. Effects of mulching on soil properties and growth of tea olive (Osmanthus fragrans). PLoS ONE 2016, 11, e0158228. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Kurniadie, D. Allelopathy of Lantana camara as an invasive plant. Plants 2021, 10, 1028. [Google Scholar] [CrossRef]
- Hussain, F.; Ghulam, S.; Sher, Z.; Ahmad, B. Allelopathy by Lantana camara. Pak. J. Bot. 2011, 43, 2373–2378. [Google Scholar]
- Chick, T.A.; Kielbaso, J.J. Allelopathy as an inhibition factor in ornamental tree growth: Implications from the literature. J. Arboric. 1998, 24, 274–279. [Google Scholar] [CrossRef]
- Vallejo, V.R.; Serrasolses, I.; Cortina, J.; Seva, J.P.; Valdecantos, A.; Vilagrosa, A. Restoration strategies and actions in Mediterranean degraded lands. In Desertification in Europe: Mitigation Strategies and Land-Use Planning; Enne, G., Zanolla, C., Peter, D., Eds.; European Commission: Luxembourg, 2000; pp. 221–233. [Google Scholar]
- Caravaca, F.; Barea, J.M.; Palenzuela, J.; Figueroa, D.; Alguacil, M.M.; Roldán, A. Establishment of shrub species in a degraded semiarid site after inoculation with native or allochthonous arbuscular mycorrhizal fungi. Appl. Soil Ecol. 2003, 22, 103–111. [Google Scholar] [CrossRef]
- Green, J.J.; Baddeley, J.A.; Cortina, J.; Watson, C.A. Root development in the Mediterranean shrub Pistacia lentiscus as affected by nursery treatments. J. Arid Environ. 2005, 61, 1–12. [Google Scholar] [CrossRef]
- Gilman, E.F.; Wiese, C.L.; Paz, M.; Shober, A.L.; Scheiber, S.M.; Moore, K.A.; Brennan, M. Effects of irrigation volume and frequency on shrub establishment in Florida. J. Environ. Hortic. 2009, 27, 149–154. [Google Scholar] [CrossRef]
- Maestre, F.T.; Bautista, S.; Cortina, J.; Bellot, J. Potential for using facilitation by grasses to establish shrubs on a semiarid degraded steppe. Ecol. Appl. 2001, 11, 1641–1655. [Google Scholar] [CrossRef]
- Reddy, P.P. Micro-Catchment Rainwater Harvesting. In Sustainable Intensification of Crop Production; Springer: Singapore, 2016. [Google Scholar] [CrossRef]
- Ali, A.; Yazar, A. Effect of micro-catchment water harvesting on soil-water storage and shrub establishment in the arid environment. Int. J. Agric. Biol. 2007, 9, 2007. [Google Scholar]
- Montague, T.; McKenney, C.; Maurer, M.; Winn, B. Influence of irrigation volume and mulch on establishment of select shrub species. Arboric. Urban For. 2007, 33, 202. [Google Scholar] [CrossRef]



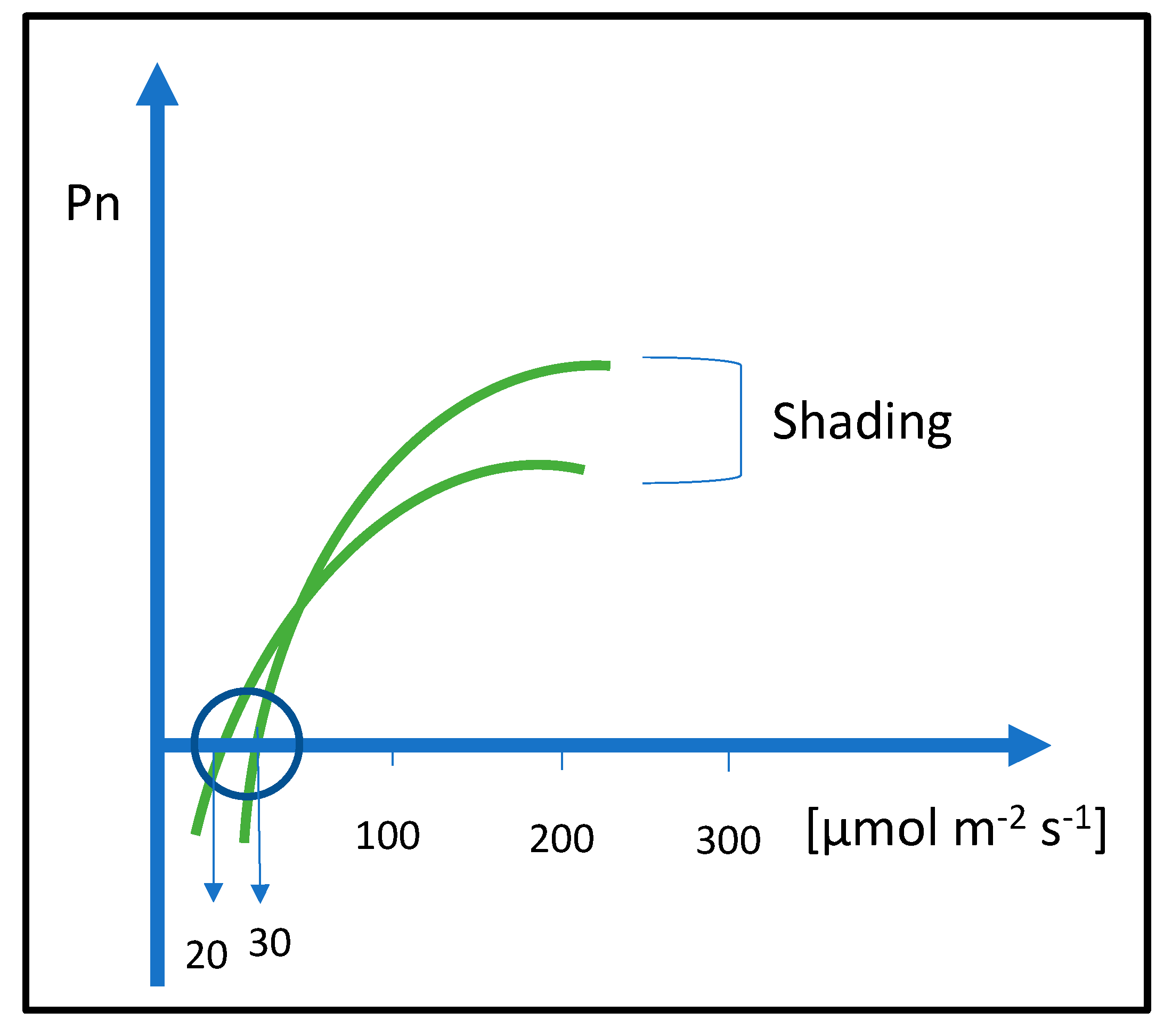{kind=link}
{kind=link}
| Target Organs | Stress Effects | Tolerance or Adaptation Response | References |
|---|---|---|---|
| Roots | Increase of root biomass | Increase the functional roots and architectures | [30] |
| Stem | Decrease the growth, elongation, diameter and biomass | Increase the lignification process (chi lo dice?) | [30,43,44] |
| Leaves | Reduction of size and leaf number | Increase the wax or thickness, and trichome number | [27,45] |
| Flowers | Reduction of flower production and longevity | Increase the flower longevity and turnover | [46] |
| Target Organs | Stress Effects | Tolerance or Adaptation Response | Reference |
|---|---|---|---|
| Roots | Increase of roots biomass | Increase the water uptake and exclusion of some toxic ions such as Na+ or Cl− | [63] |
| Stem | Decrease the growth and biomass | Increase the extrusion or storage | [64] |
| Leaves | Reduction of size, necrosis, or abscission | Increase the storage of ions in vacuole | [49] |
| Flowers | Reduction of flowers production and longevity | Increase the flower longevity and turnover | [65] |
| Target Organs | Stress Effects | Tolerance or Adaptation Response | References |
|---|---|---|---|
| Roots | Increase the ethylene production | Cell death and in some herbaceous plants the aerenchyma formation | [77,78] |
| Stem | Decrease the growth and biomass | Increase the ethylene biosynthesis | [77] |
| Leaves | Leaf yellowing, abscission | Increase the storage of ions in vacuole | [77] |
| Flowers | Reduction of flower production and longevity | Increase the flower longevity | [78] |
| Target Organs | Stress Effects | Tolerance or Adaptation Response | References |
|---|---|---|---|
| Roots | Decrease of roots biomass | - | [86] |
| Stem | Increase of stem elongation | Increase the firmness of cell walls | [86] |
| Leaves | Reduction of thickness and leaf number | Increase the chlorophyll concentration and lowering the light compensation point | [86,87] |
| Flowers | Reduction of flower number | Increase the flower longevity and turnover | [87] |
| Target Organs | Stress Effects | Tolerance or Adaptation Response | References |
|---|---|---|---|
| Roots | Decrease of roots functionality | Increase the roots uptake of ions | [90,91] |
| Stem | Increase or decrease of stem growth | Increase the firmness of cell walls | [92] |
| Leaves | Increase of thickness, leaf necrosis | reduction the chlorophyll concentration | [92] |
| Flowers | Reduction of flower numbers | - | [93] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leotta, L.; Toscano, S.; Ferrante, A.; Romano, D.; Francini, A. New Strategies to Increase the Abiotic Stress Tolerance in Woody Ornamental Plants in Mediterranean Climate. Plants 2023, 12, 2022. https://doi.org/10.3390/plants12102022
Leotta L, Toscano S, Ferrante A, Romano D, Francini A. New Strategies to Increase the Abiotic Stress Tolerance in Woody Ornamental Plants in Mediterranean Climate. Plants. 2023; 12(10):2022. https://doi.org/10.3390/plants12102022
Chicago/Turabian StyleLeotta, Luca, Stefania Toscano, Antonio Ferrante, Daniela Romano, and Alessandra Francini. 2023. "New Strategies to Increase the Abiotic Stress Tolerance in Woody Ornamental Plants in Mediterranean Climate" Plants 12, no. 10: 2022. https://doi.org/10.3390/plants12102022
APA StyleLeotta, L., Toscano, S., Ferrante, A., Romano, D., & Francini, A. (2023). New Strategies to Increase the Abiotic Stress Tolerance in Woody Ornamental Plants in Mediterranean Climate. Plants, 12(10), 2022. https://doi.org/10.3390/plants12102022