1. Introduction
In recent years, the availability of fresh water has caused concerns, especially in semi-arid regions [
1]. Climate change that has occurred all over the world has increased atmospheric temperatures and caused droughts [
2,
3]. Thus, the climatic imbalance between evaporation and precipitation rates promotes the increase in salt concentrations in sources of water used for irrigation [
4,
5].
High salinity in water and/or soil causes salt stress, one of the environmental stresses that most compromise sustainability and agricultural production worldwide [
6]. Salt stress causes damage to agricultural production and inhibits crop growth due to reduced water availability for plants, caused by the decrease in the osmotic potential of the soil solution, leading to stomatal closure and compromising transpiration and the CO
2 assimilation rate [
7]. Salinity reduces chlorophyll synthesis and photochemical efficiency, limiting the process of photosynthesis, which directly affects crop production [
8]. According to Dias et al. [
9], irrigation with saline water also reduces pulp yield and compromises the postharvest quality of fruits, as observed by Andrade et al. [
10], who found reductions in the number of fruits and in the average weight of sour passion fruit irrigated by water with electrical conductivity (ECw) above 0.7 dS m
−1, while Lima et al. [
11] found reductions in the polar and equatorial diameter of passion fruit as a function of salt stress; in another study, Ramos et al. [
12] observed reductions in pulp yield and postharvest quality of passion fruit irrigated with ECw above 0.6 dS m
−1.
Recent studies have shown that irrigation with saline water can compromise the production components of passion fruit and reduce its yield [
13,
14,
15]. However, the severity of the effect of salt stress on plants depends on other factors, such as genotype, time of exposure to stress, edaphoclimatic conditions of the region, irrigation management, fertilization, and foliar application of elicitor substances [
3,
16].
Thus, studies that enable the use of saline waters in irrigated agriculture are important to ensure the sustainability of crops. In this context, the use of salicylic acid has emerged as a promising alternative for minimizing the deleterious effects caused by salt stress on plants [
17,
18,
19].
Salicylic acid is a natural phenolic compound, which acts as a non-enzymatic antioxidant and an endogenous signaling molecule, inducing tolerance to salt stress [
20]. Its beneficial effects are dependent on the concentration, plant species, stage of crop development, and method of application [
3,
21,
22].
Several studies have reported that foliar spraying with salicylic acid can attenuate the deleterious effects caused by salt stress in strawberry [
23], American grapevine [
24], date palm [
25], orange [
26], and soursop [
18]. However, information on its use in sour passion fruit crops irrigated with saline water is incipient in the literature.
Sour passion fruit (
Passiflora edulis Sims) is the most recognized and cultivated
Passifloraceae species in Brazil, especially in the Northeast region [
27]. Brazil stands out as the largest producer and consumer of passion fruit in the world, producing 683,993 tons in 2021 in an area of 44,827 ha, achieving an average yield of 15.26 t ha
−1, with the Northeast region accounting for 69.6% (476,006 tons) of the national production [
28].
Due to the physical-chemical quality and acceptance by consumers, its fruits are used for fresh consumption and/or through agro-industrial processing in the preparation of carbonated and mixed drinks, syrups, jellies, dairy products, ice cream and canned foods [
29,
30]. It should be noted that passion fruit is considered a salt-sensitive crop [
31], with an irrigation water salinity threshold of 1.3 dS m
−1 [
32,
33].
This study is based on the hypothesis that foliar application of salicylic acid at adequate concentrations mitigates the deleterious effects caused by irrigation with saline waters on the physiology, production components, and postharvest quality of the fruits of sour passion fruit, inducing the tolerance of plants to salt stress by increasing the biosynthesis of organic compounds and modulating activities of enzymes that detoxify reactive oxygen species. In view of the above, the aim of this study was to evaluate the effect of foliar application of salicylic acid concentrations on the induction of tolerance of sour passion fruit to salt stress in two cropping cycles.
3. Discussion
Salt stress caused by excess salts in water and/or soil limits plant growth and development [
34] and negatively affects ionic homeostasis and, consequently, nutrient absorption, causing losses in agricultural production worldwide, especially in arid and semi-arid regions [
35,
36,
37]. Fruit crops generally have a greater sensitivity to salt stress than annual crops [
38].
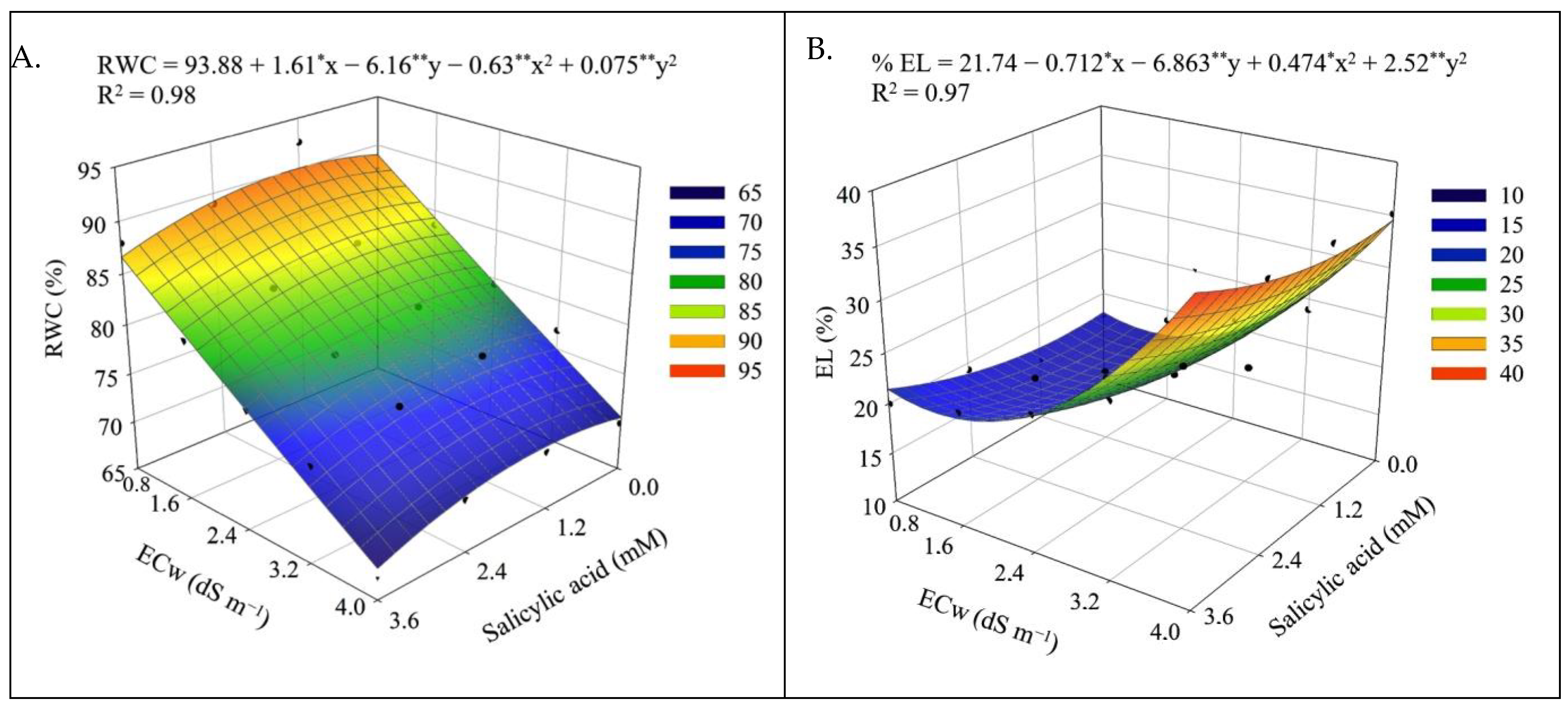
The results obtained in this study show that the salt stress caused by the increase in the electrical conductivity of irrigation water negatively affected the leaf water status, observed by the relative water content, promoted an increase in the electrolyte leakage in the leaf blade, and reduced gas exchange, synthesis of photosynthetic pigments and chlorophyll a fluorescence, negatively affecting the production components and postharvest quality of sour passion fruits.
Two studies have indicated that the sensitivity and tolerance of crops to salt stress may vary between species and cultivars of the same species, in addition to depending on climatic conditions, type of soil, plant development stage, irrigation method, and time of exposure to salt stress [
16,
39]. In the present study, it was observed that the deleterious effects of salinity were intensified in the second cycle, possibly due to a longer period of exposure to salt stress and the accumulation of salts in the soil.
In the first cycle of sour passion fruit, foliar application of salicylic acid between the estimated concentrations of 1.0 and 1.4 mM was able to partially mitigate the effects of salt stress on the relative water content in the leaf blade, electrolyte leakage, gas exchange, and photosynthetic pigments. In addition, salicylic acid promoted an increase in the production components and postharvest quality of the fruits of sour passion fruit. In the second cycle, there was no effect of the application of salicylic acid on any of the variables analyzed, which may be related to the lower number of applications due to the reduction in the cycle, as the first cycle lasted 258 days, while the second cycle lasted 172 days, that is, a reduction of 86 days.
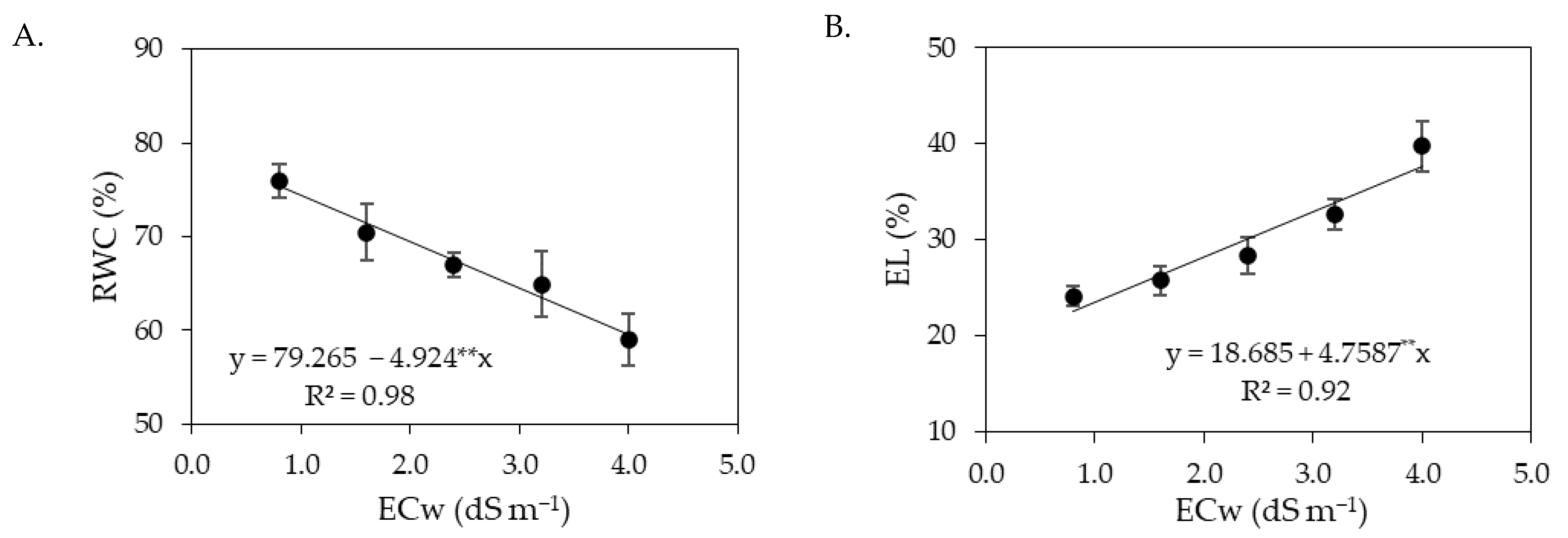
The decrease in the relative water content in the leaves (
Figure 1A and
Figure 9A) and the increase in electrolyte leakage (
Figure 1B and
Figure 9B) with the increase in ECw can be seen as a harmful effect of salinity. The reduction in RWC results from the loss of turgor of plant tissues, since salinity causes osmotic stress, which hinders the absorption and translocation of water from the soil to the plant, affecting its growth and metabolism [
40]. Salt stress also favors the production of reactive oxygen species (ROS), such as superoxide radical, hydroxyl radicals, and hydrogen peroxide [
41]. Under normal conditions, the production and elimination of ROS are balanced, but under salt stress conditions, there is an imbalance between production and elimination, which can cause photo-oxidative damage to photosystems and the cell membrane peroxidation [
42,
43], generating an increase in the percentage of electrolyte leakage, as observed in the present study (
Figure 1B and
Figure 9B).
In a study conducted by Silva Neta et al. [
44] with the sour passion fruit ‘BRS Rubi do Cerrado’ under irrigation with saline water (ECw ranging from 0.3 to 3.5 dS m
−1), a reduction in RWC was also observed with the increase in the levels of electrical conductivity of irrigation water, equal to 7.73% when comparing plants irrigated with ECw of 3.5 dS m
−1 to those cultivated with an ECw of 0.3 dS m
−1. Wanderley et al. [
45] evaluated yellow passion fruit under salt stress (ECw ranging from 0.3 to 3.1 dS m
−1) and observed an increase of 24.65% in the percentage of electrolyte leakage in plants irrigated with an ECw of 3.1 dS m
−1 compared to those cultivated with an ECw of 0.3 dS m
−1.
Despite the deleterious effects of salt stress on RWC and EL, foliar application of salicylic acid in concentrations between 1.0 and 1.2 mM mitigated the effects of irrigation water salinity in the first cropping cycle of sour passion fruit. Salicylic acid acts to protect plant cells from the toxicity caused by ion accumulation and improves antioxidant activity, nitrogen metabolism, and water absorption in plants [
46,
47], increasing leaf turgor and reducing cell membrane damage [
1].
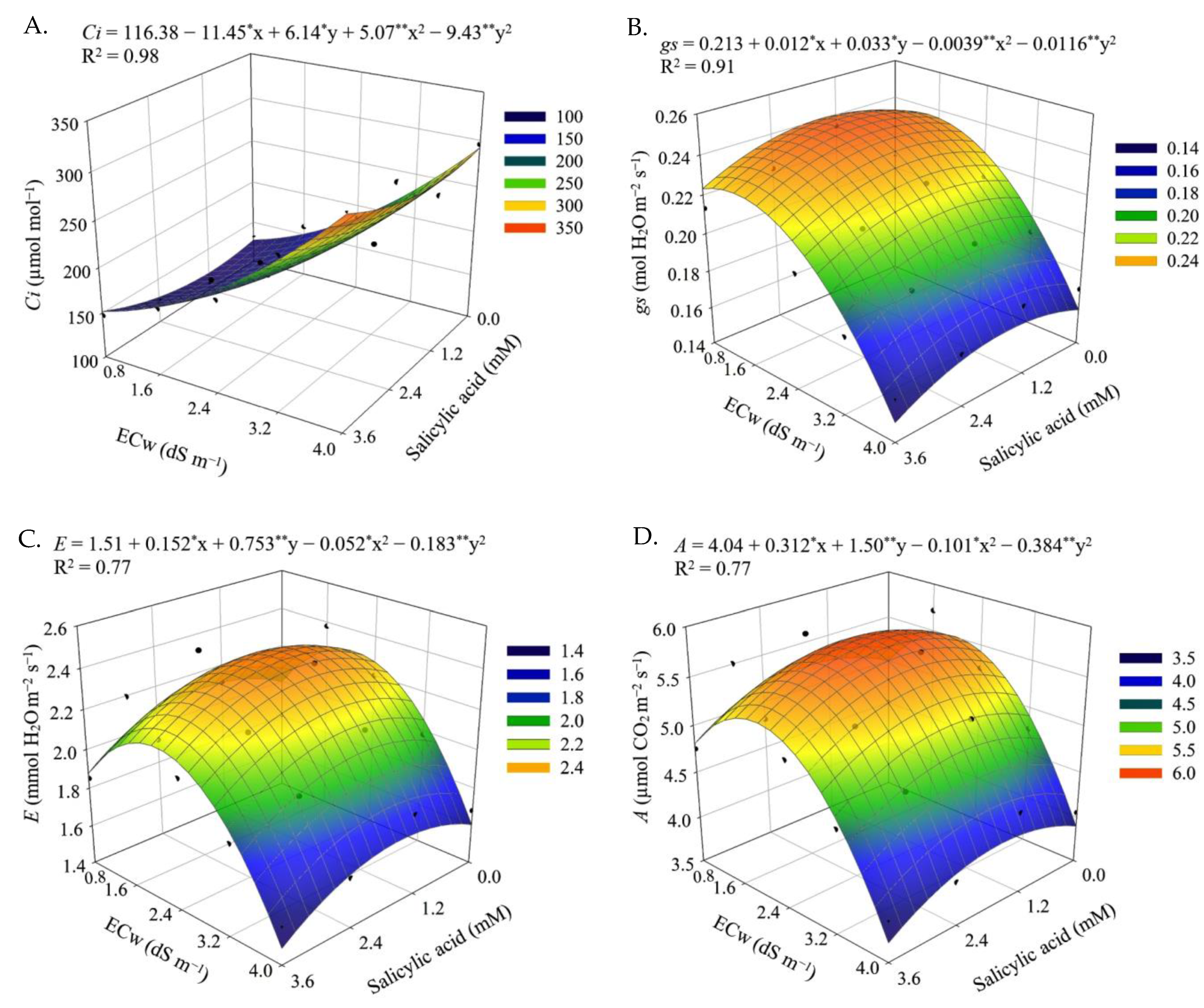
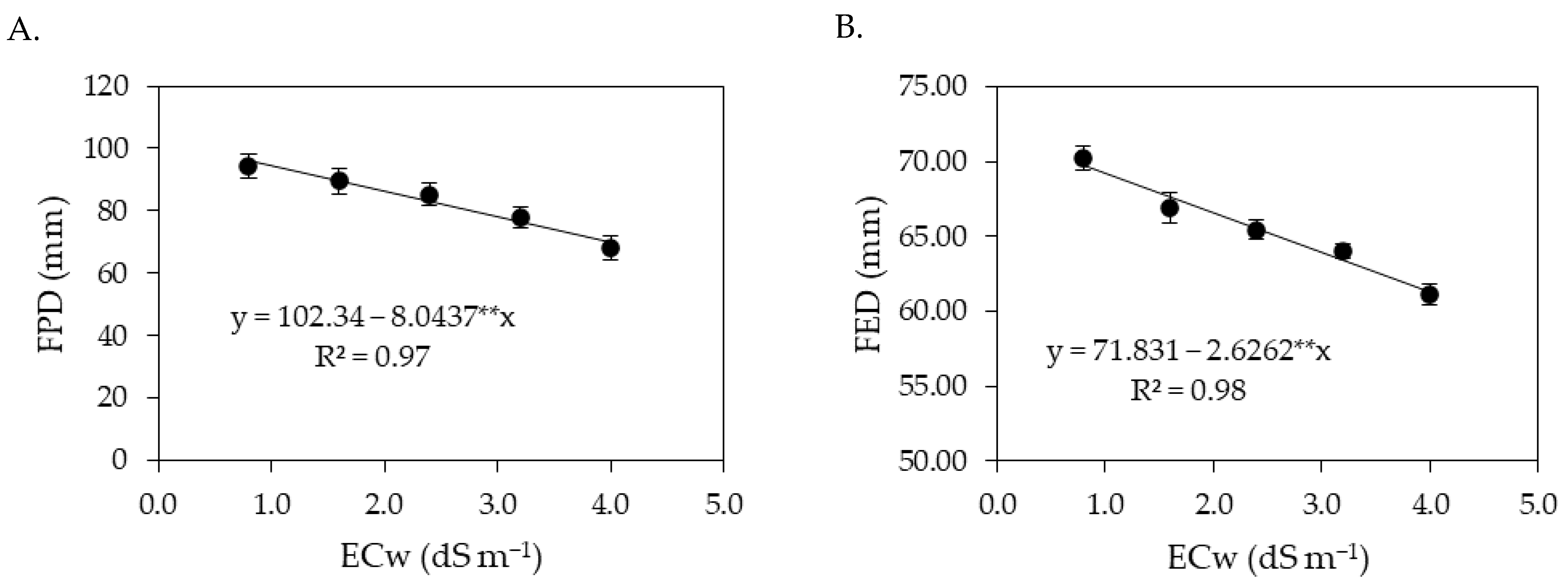
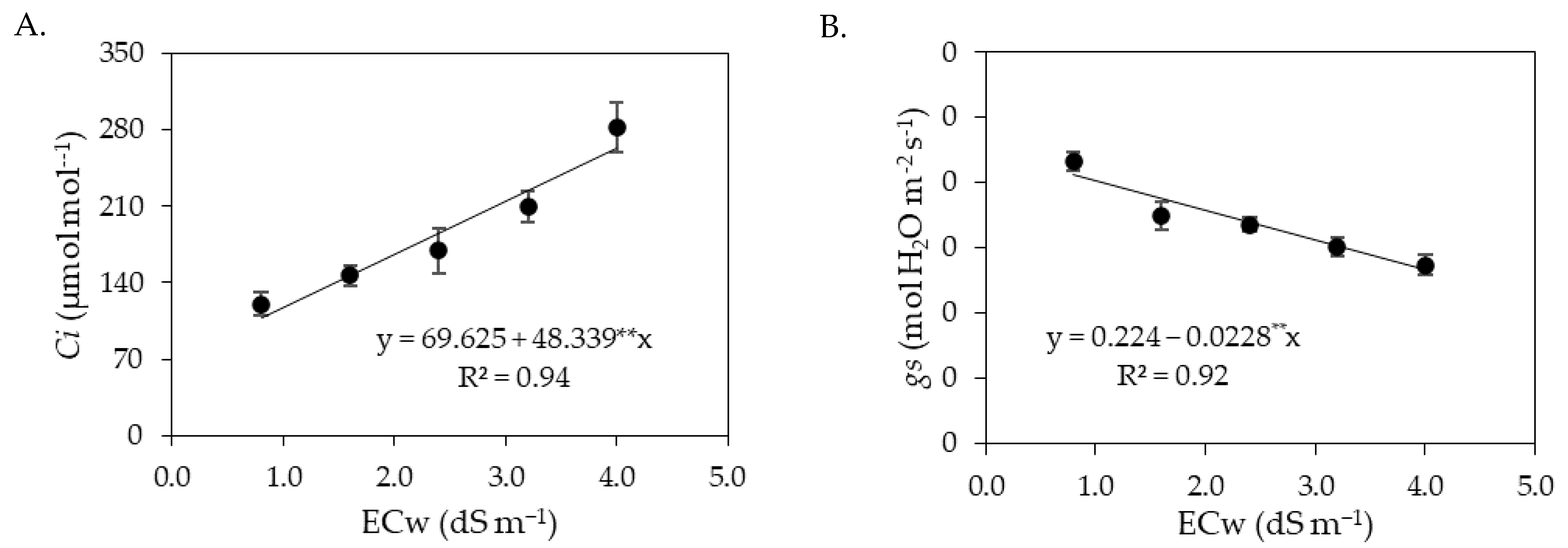
The results obtained in the present study indicate that salt stress caused by the increase in the electrical conductivity of irrigation water negatively impacted the gas exchange variables (
Ci,
gs,
E and
A) of sour passion fruit in both cycles. When subjected to salt stress, plants tend to close their stomata in order to reduce water loss to the atmosphere [
18]. In addition, the reduction in leaf turgor, as observed in this study (
Figure 1A and
Figure 9A), induces stomatal closure, as a defense mechanism of the plant against the loss of water by transpiration, affecting physiological processes, such as stomatal conductance, internal CO
2 concentration and CO
2 assimilation rate [
48]. Reductions in gas exchange in sour passion fruit plants caused by irrigation with saline water have also been reported in other studies, such as [
5,
13,
16,
30].
Stomata are the structures responsible for regulating the gas exchange of plants [
49]. In this study, the first cycle of sour passion fruit foliar application of salicylic acid at a concentration of 1.2 mM increased stomatal conductance, resulting in improvements in the internal CO
2 concentration, transpiration, and CO
2 assimilation rate (
Figure 2). Previous studies have shown that salicylic acid can reduce lipid peroxidation and interact with other plant hormones, increasing the tolerance of plants to salt stress [
50,
51]. In addition, the beneficial effects of salicylic acid observed in gas exchanges may be related to the accumulation of osmoprotectants, improving the turgor of plant cells under stress and the activation of antioxidant enzymes, resulting in better photosynthetic activity [
52].
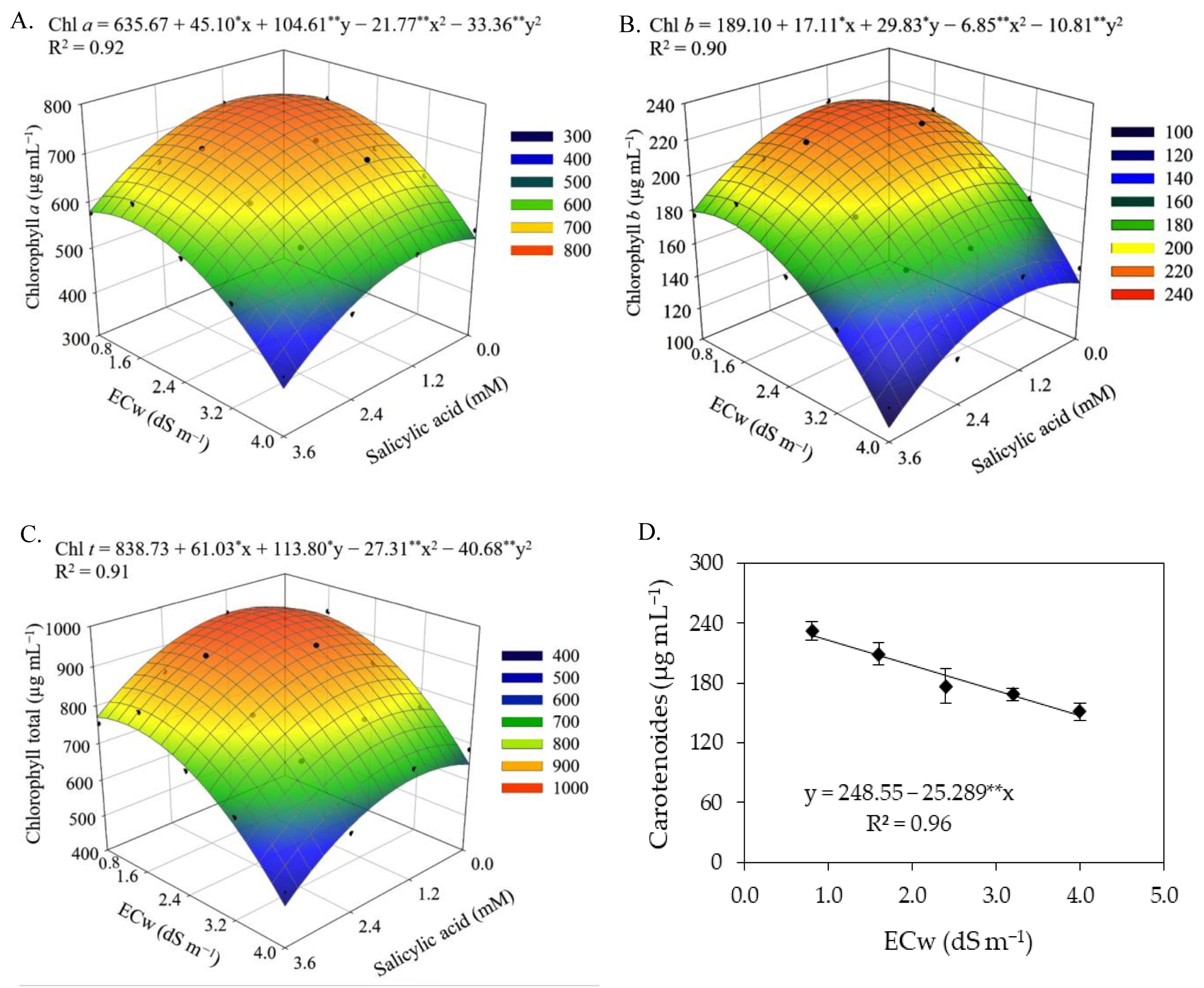
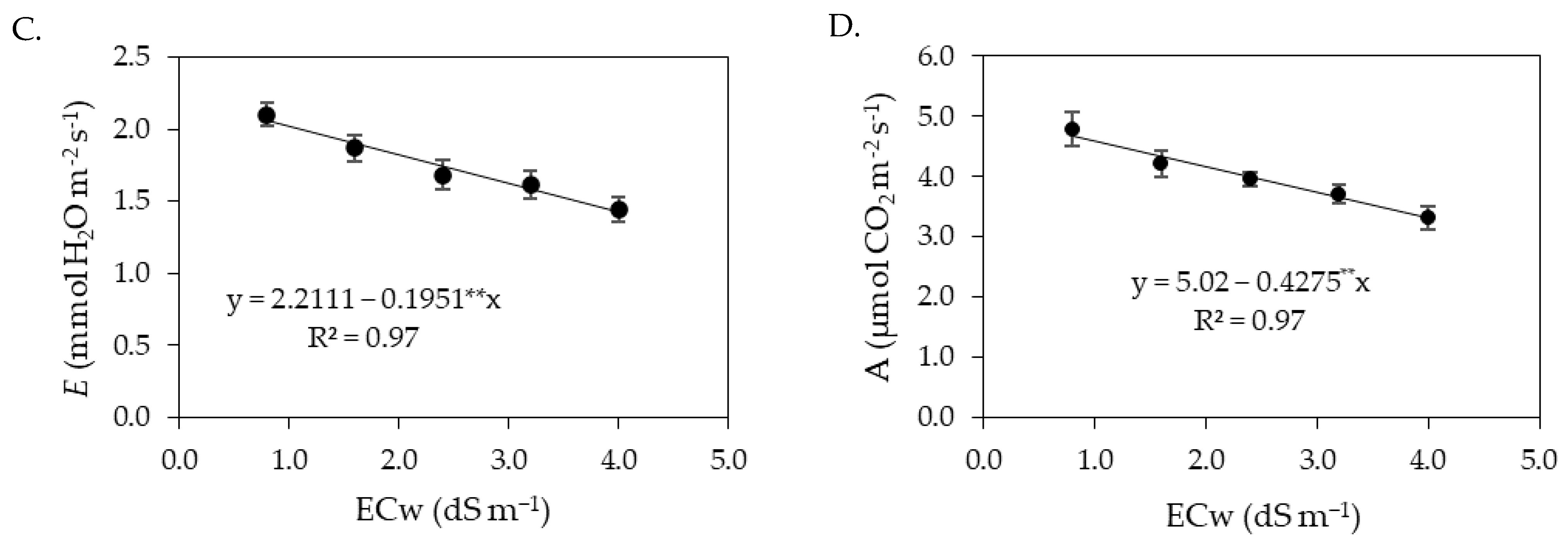
Photosynthetic pigments are considered determinant elements for plant growth and development [
53]. The results of this study showed that the increase in electrical conductivity of irrigation water negatively affected the photosynthetic pigments of sour passion fruit, especially in the second cycle. The excess salts present in the irrigation water inhibits the activity of 5-aminolevulinic acid, which is a precursor of chlorophyll, in addition to increasing the activity of the chlorophyllase enzyme, which acts by degrading the molecules of photosynthetic pigments, causing damage to chloroplasts and limiting the activity of pigmentation proteins [
54,
55]. Reductions in photosynthetic pigments in sour passion fruit plants as a function of irrigation with saline water have also been reported in studies conducted by [
14,
56,
57].
The foliar application of salicylic acid at the estimated concentration of 1.0 mM mitigated the effects of salt stress on the synthesis of chlorophyll
a, chlorophyll
b, and total chlorophyll in sour passion fruits in the first cropping cycle. According to Hundare et al. [
58], salicylic acid can stimulate the biosynthesis of chlorophyll and/or reduce its degradation, improving plant growth and development. In agreement with the present study, Silva et al. [
34] evaluated the effect of foliar application of salicylic acid on photosynthetic pigments in soursop under salt stress and found that foliar application of salicylic acid at a concentration of 1.4 mM reduced the effects of irrigation water salinity, promoting an increase in the contents of photosynthetic pigments (Chl
a, Chl
b, Chl
t, and Car).
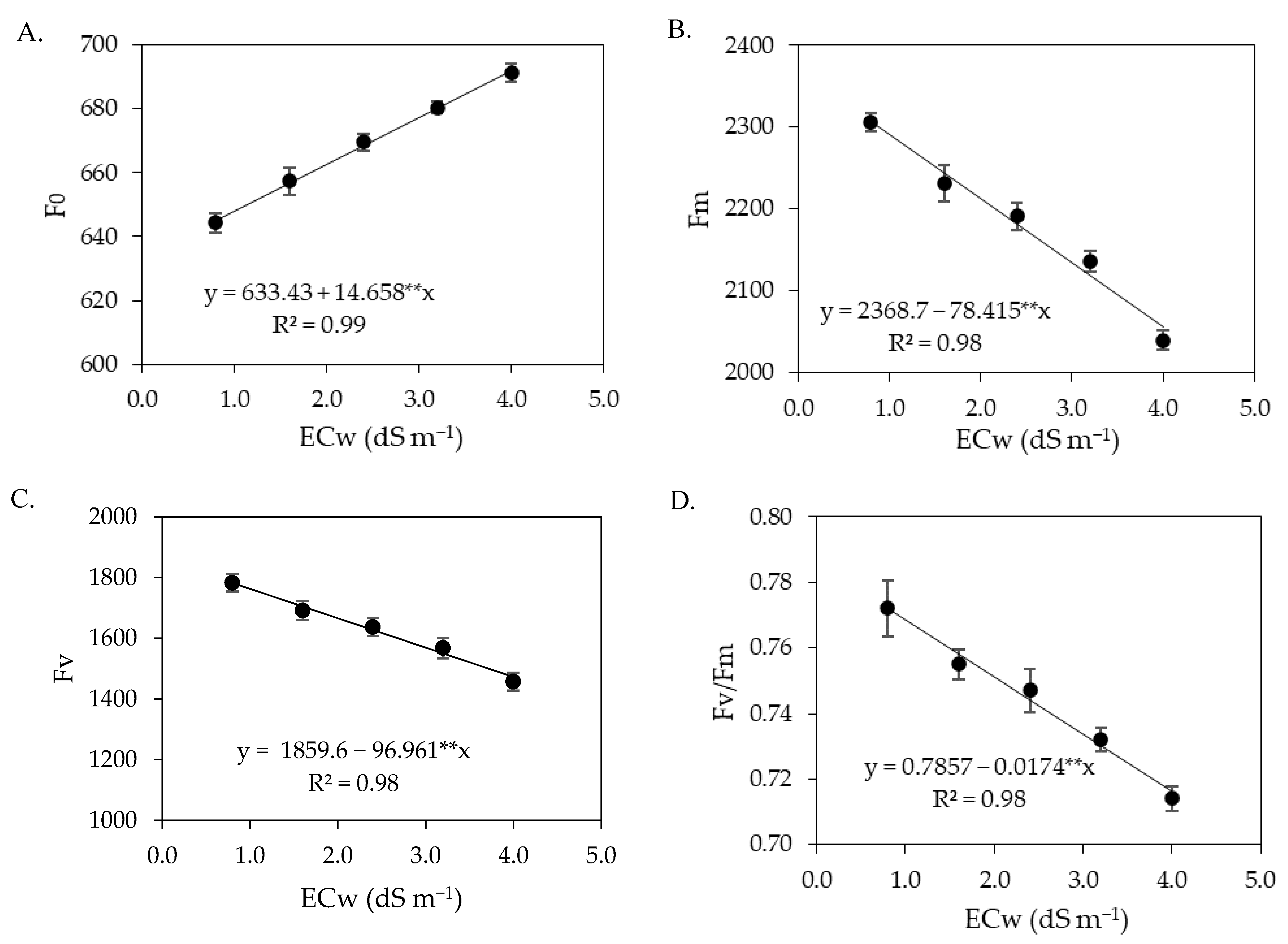
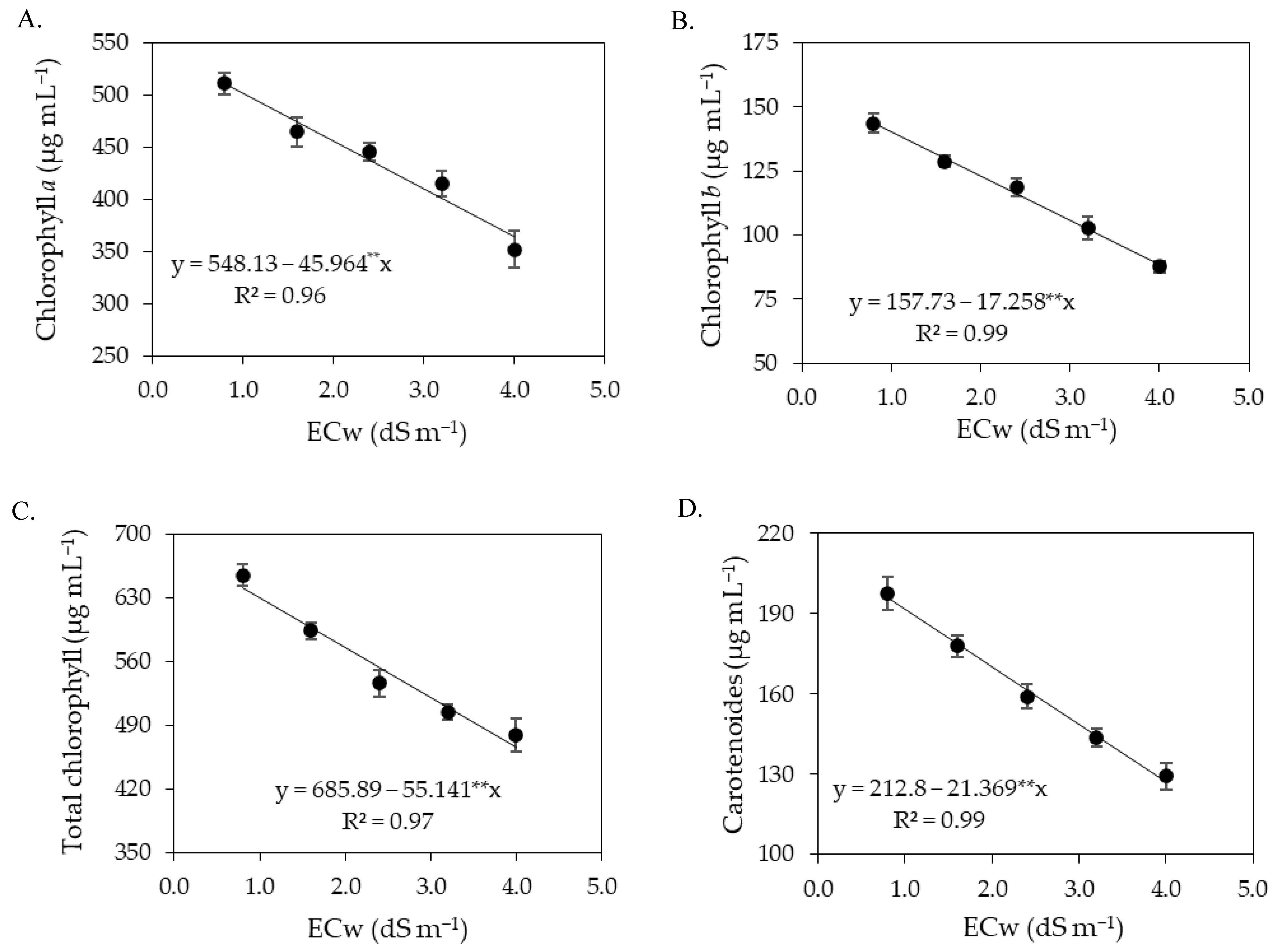
Chlorophyll fluorescence variables are widely used to explain chlorophyll energy dissipation in thylakoid membranes [
59,
60]. In the present study, it was observed that the concentrations of salicylic acid were not able to attenuate the effects of salt stress on chlorophyll fluorescence in any of the cropping cycles. The increase in the electrical conductivity of irrigation water promoted an increase in initial fluorescence (
Figure 4 and
Figure 12), indicating damage to the light-harvesting complex of the photosystem II of sour passion fruit. According to Kalaji et al. [
61], the increase in initial fluorescence results in the utilization of less photochemical energy in the reaction centers of photosystem II, thus serving as an indicator of salt stress effects.
In turn, maximum fluorescence and variable fluorescence were reduced by the increase in the electrical conductivity of irrigation water. These results may be related to low efficiency in quinone photoreduction and electron flow between photosystems, which results in low PSII activity in the thylakoid membrane, directly influencing the flow of electrons between photosystems [
62,
63]. In addition, it may indicate that the photosynthetic apparatus was damaged by salt stress, compromising photosystem II, with negative effects on the photosynthetic process [
1], as observed in this study.
The reductions in maximum (Fm) and variable (Fv) fluorescence caused by the increase in the electrical conductivity of irrigation water contributed negatively to reducing the quantum efficiency of photosystem II (Fv/Fm). Several authors consider Fv/Fm values between 0.75 and 0.85 as normal in unstressed plants [
18,
64,
65]. Thus, the results indicate that irrigation with an ECw above 2.0 dS m
−1 in the first cycle and above 1.4 dS m
−1 in the second cycle negatively affected Fv/Fm as the values were lower than 0.75.
The reduction in the quantum efficiency of photosystem II of sour passion fruits shows lower activity of P
680 under salt stress conditions, which may be associated with the activity of the chlorophyllase enzyme, which reduces the chlorophyll contents and, consequently, affects the electron capture and transport between the reaction centers to free plastoquinone [
66]. Reduction in Fv/Fm in sour passion fruit plants due to salt stress was also verified by Andrade et al. [
56], who found a decrease of 6.36% per unit of ECw increase, highlighting it as indicative of the occurrence of a photoinhibition effect caused by salt stress.
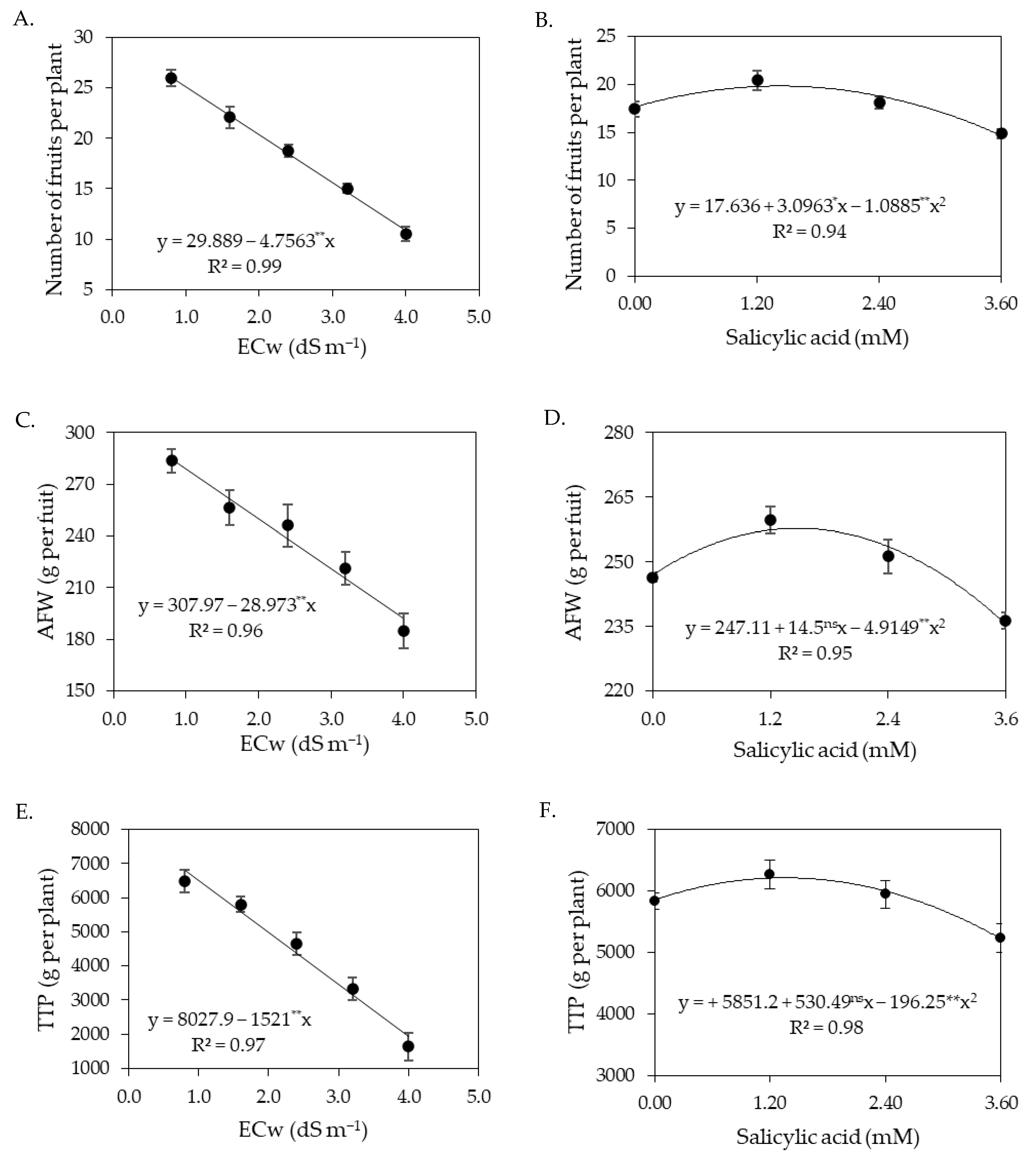
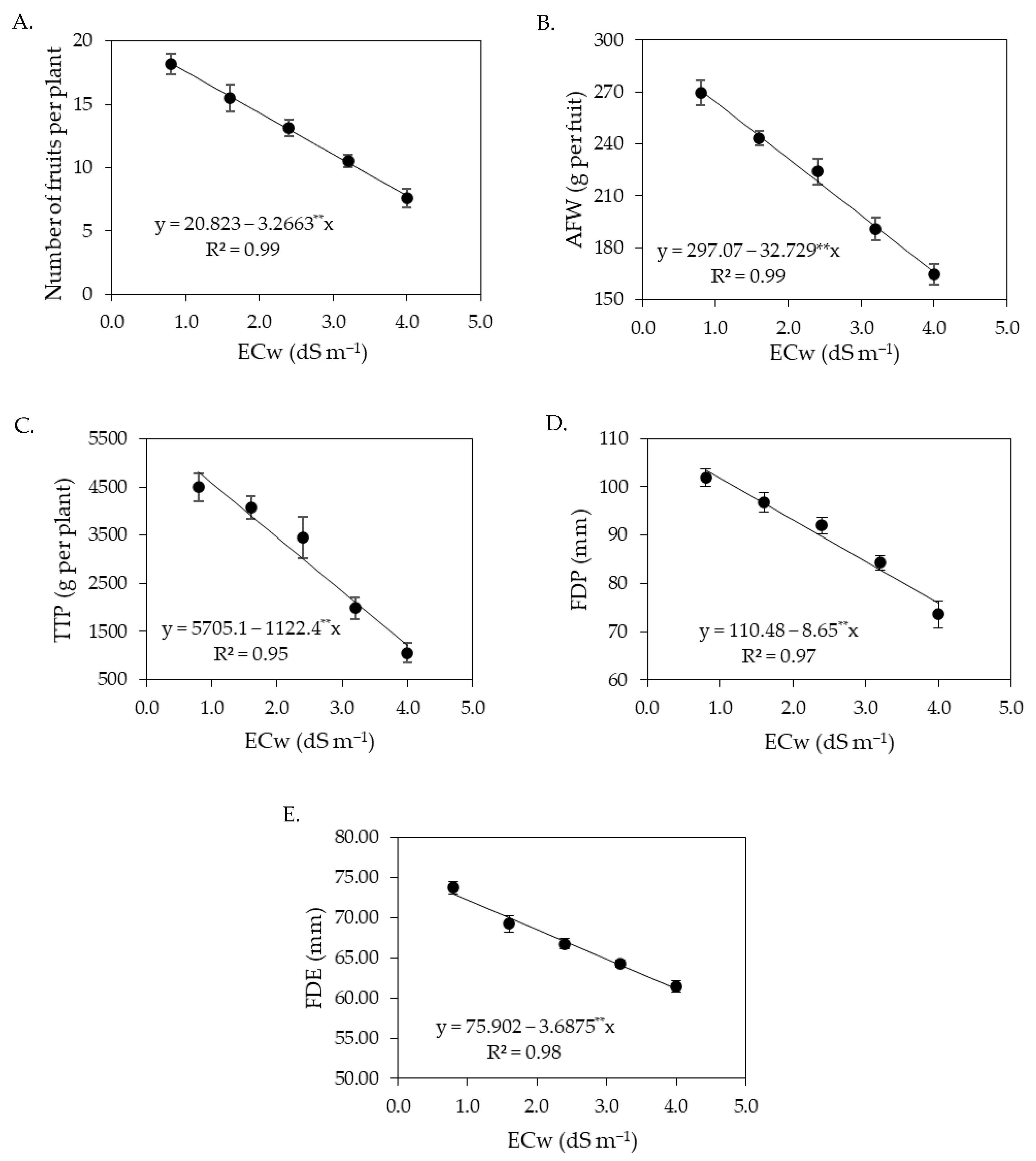
The increase in the electrical conductivity of irrigation water negatively affected the production components of sour passion fruits, observed by the number of fruits per plant, average fruit weight, total production per plant, and polar and equatorial diameter of fruits, with the most intense reductions in the second cycle. Under salt stress, plants suffer from water and nutrient deficiencies due to the osmotic effect that hinders the absorption of water and nutrients [
67]. The excess of salts in irrigation water also induces the inhibition of physiological and metabolic processes, negatively affecting the production components [
68]. Reductions in the production components of sour passion fruits as a function of irrigation water salinity have also been reported in studies conducted by [
13,
15,
39].
Despite the reduction in the production components of sour passion fruits, it was observed in this study that the foliar application of salicylic acid at concentrations between 1.4 and 1.5 mM promoted an increase in the number of fruits per plant, average fruit weight, and total production per plant (
Figure 5) in the first cropping cycle. In this study, it was observed that salicylic acid regulated stomatal conductance (
Figure 2B) and increased photosynthetic activity (
Figure 2B) and the synthesis of chlorophyll (
Figure 3), contributing directly to an increase in the production components. The beneficial effect of salicylic acid on the production components of sour passion fruit may be related to its role in reducing the absorption of Na
+ and increasing that of N, P, K, Ca, and Mg by plants [
69].
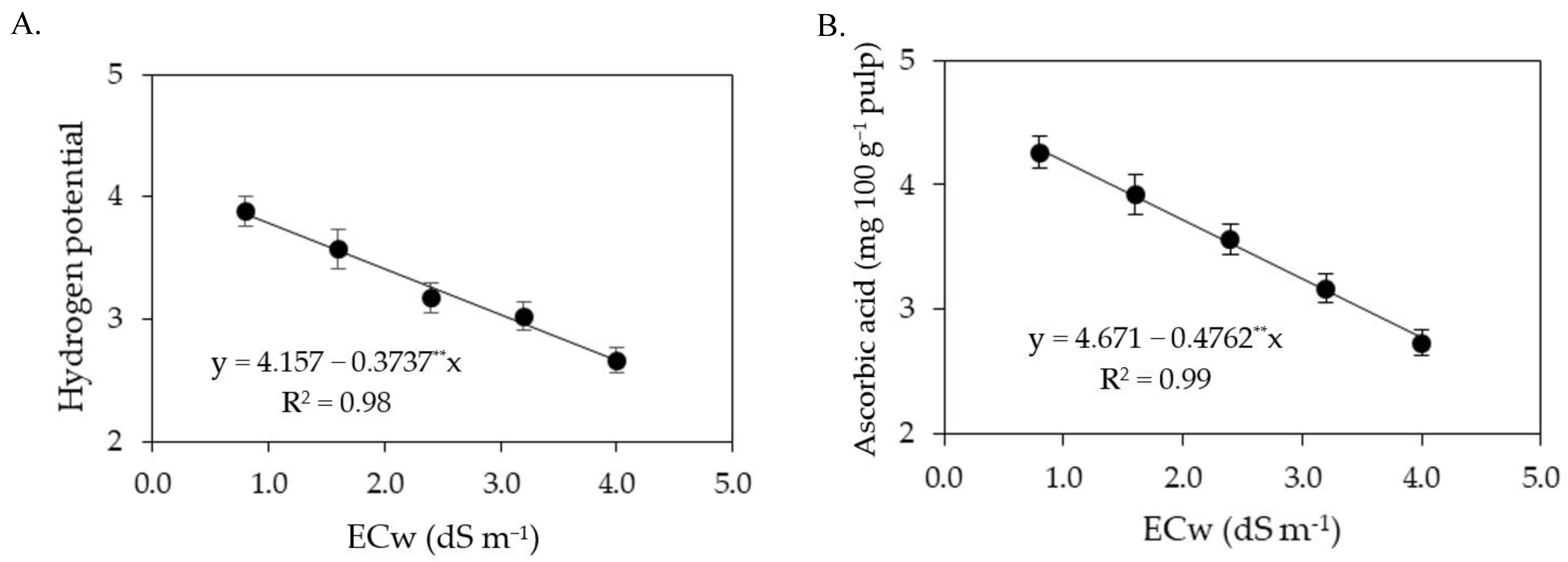
The increase in the electrical conductivity of irrigation water reduced the pH of sour passion fruit pulp both in the first and in the second cycle. According to Lacerda et al. [
19], pH is an important variable of postharvest quality, as low values can ensure the conservation of the pulp without the need for high heat treatment, thus avoiding nutritional losses. In the first cropping cycle, foliar application of salicylic acid at the estimated concentration of 1.6 mM increased the ascorbic acid contents of the sour passion fruit pulp (
Figure 7B). In the second cycle, the ascorbic acid contents were reduced by irrigation water salinity from 0.8 dS m
−1.
Soluble solids content is a parameter that has been used as an indicator of fruit quality [
70]. The increase in the electrical conductivity of irrigation water increased the soluble solids content of sour passion fruits, whose values were within the ideal range from 13 to 15 °Brix, as described by Aguiar et al. [
71]. Unlike the results obtained in this study, Ramos et al. [
12] evaluated the production and postharvest quality of passion fruit irrigated with saline water (ECw ranging from 0.6 to 3.0 dS m
−1) and found reductions in soluble solids contents with the increase in the electrical conductivity of irrigation water.
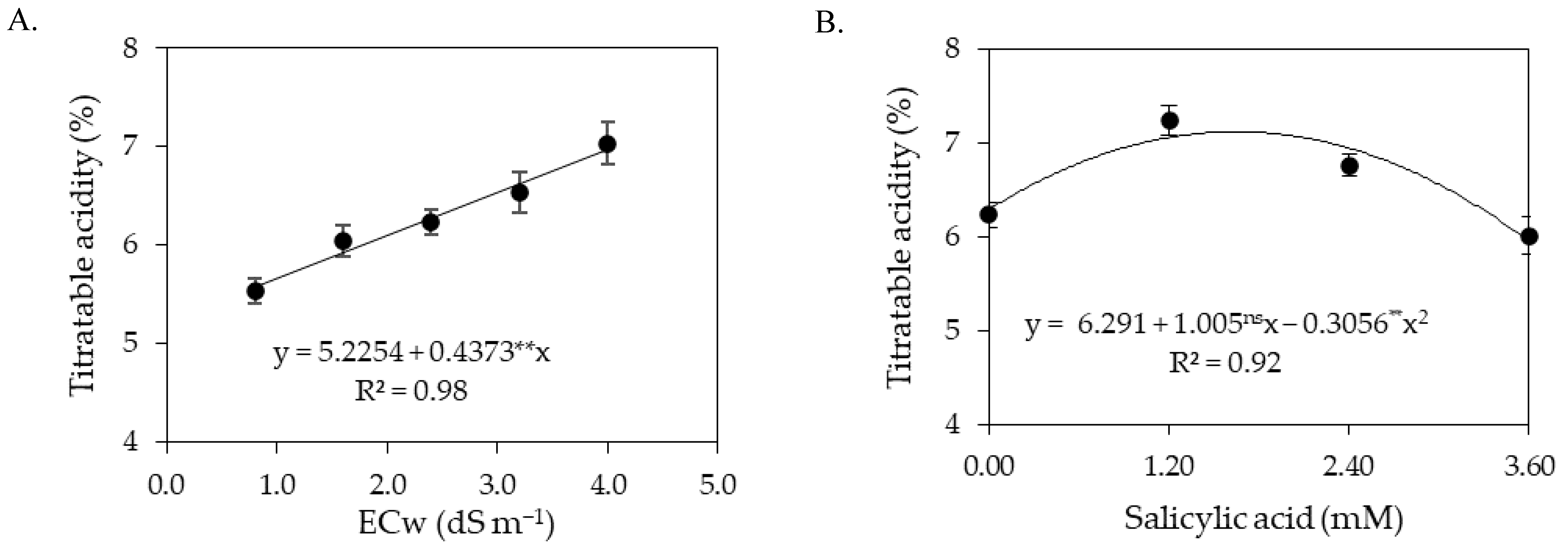
Titratable acidity in the pulp of sour passion fruit increased with the increase in the electrical conductivity of irrigation water. Titratable acidity is an important chemical attribute for the preservation of food products for both the consumer and the industry, as it makes the food more resistant to deterioration by microorganisms and allows greater flexibility in the addition of sugar, which is of particular importance in the preparation of ready-to-drink beverages [
72,
73].
In general, the results obtained indicate that irrigation with an ECw of up to 3.0 dS m−1 does not reduce the postharvest quality of the fruits of sour passion fruit, as they had values of pH, soluble solids, and titratable acidity within the technical norms for identity and quality standards for passion fruit pulp issued by the Brazilian Ministry of Agriculture, which establishes pH between 2.70 and 3.80, soluble solids greater than 11 °Brix, and titratable acidity above 2.50%.
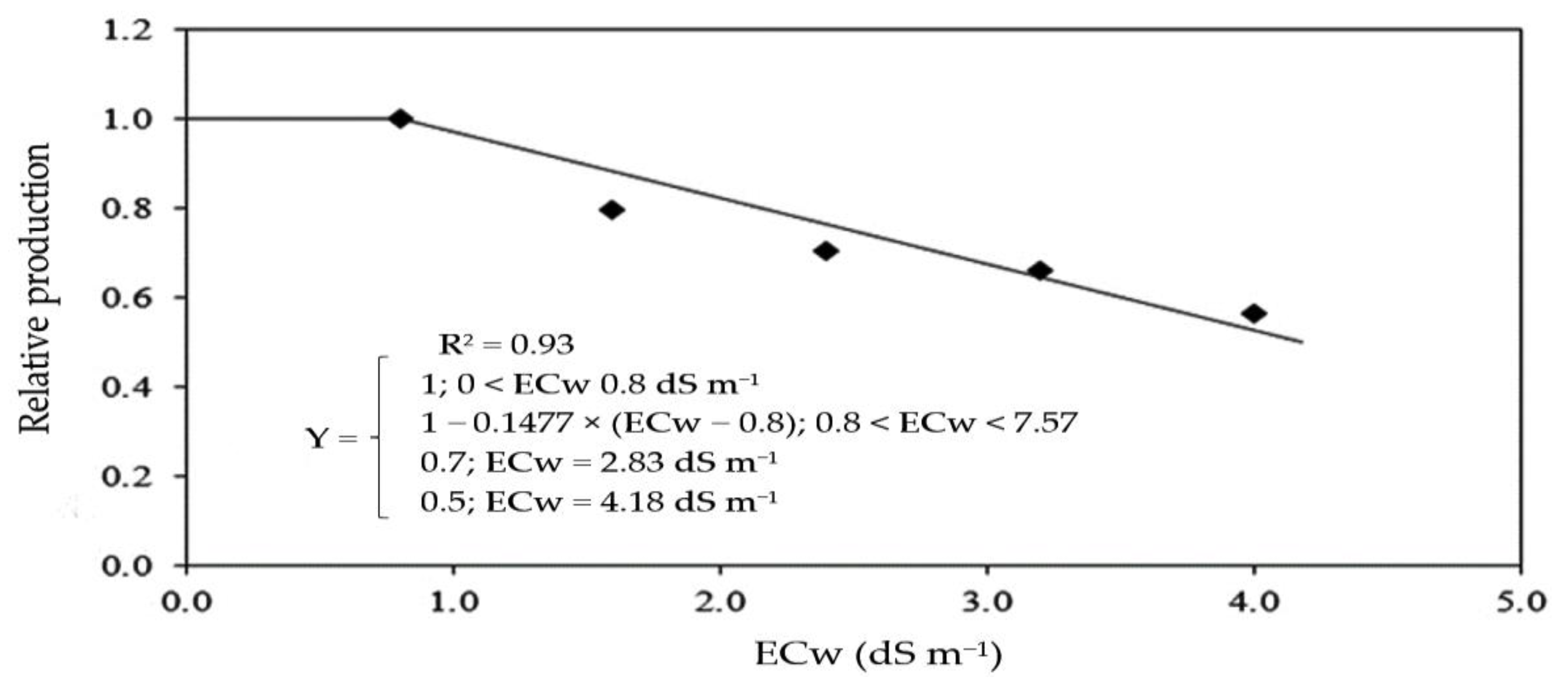
Based on the data of production per plant obtained in the first and second cycle, the tolerance of sour passion fruit to irrigation water salinity was determined through the relative production, obtained by the Plateau model followed by linear decay (
Figure 15), obtaining a salinity threshold of 0.8 dS m
−1, with a reduction of 14.78% per unit increment of ECw above this level. It is possible to obtain a relative production of 70% with an ECw of 2.83 dS m
−1 and a relative production of 50% with an ECw of 4.18 dS m
−1. The use of an electrical conductivity of irrigation water equal to or greater than 7.57 dS m
−1 will result in a relative production of 0%.
According to the criteria of degrees of tolerance based on relative production, cited by Fageria et al. [
75], considering the percentage of production loss obtained at an ECw of 4.0 dS m
−1 compared to plants irrigated with an ECw of 0.8 dS m
−1, sour passion fruit is classified as sensitive to irrigation water salinity, with a reduction of 72.81%.
In general, it was found in this study that irrigation with an ECw above 0.8 dS m−1 reduced the relative water content in the leaf blade and increased electrolyte leakage, a fact that directly reflected on gas exchange and on the biosynthesis of photosynthetic pigments; chlorophyll fluorescence was also negatively impacted, which compromised production and affected the postharvest quality of the passion fruit, especially in the second cycle. However, it was verified that the use of saline water in the cultivation of passion fruit in a semi-arid region can be made possible by the foliar application of salicylic acid in adequate concentrations. Furthermore, it was observed that SA concentrations greater than 1.4 mM can intensify the deleterious effects of saline stress on plants under semi-arid conditions.



 ,
, 























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}