

Quantitative Trait Loci for Genotype and Genotype by Environment Interaction Effects for Seed Yield Plasticity to Terminal Water-Deficit Conditions in Canola (Brassica napus L.)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Material
2.2. Experimental Design
2.3. Trait Measurements
2.4. Statistical Methods
3. Results
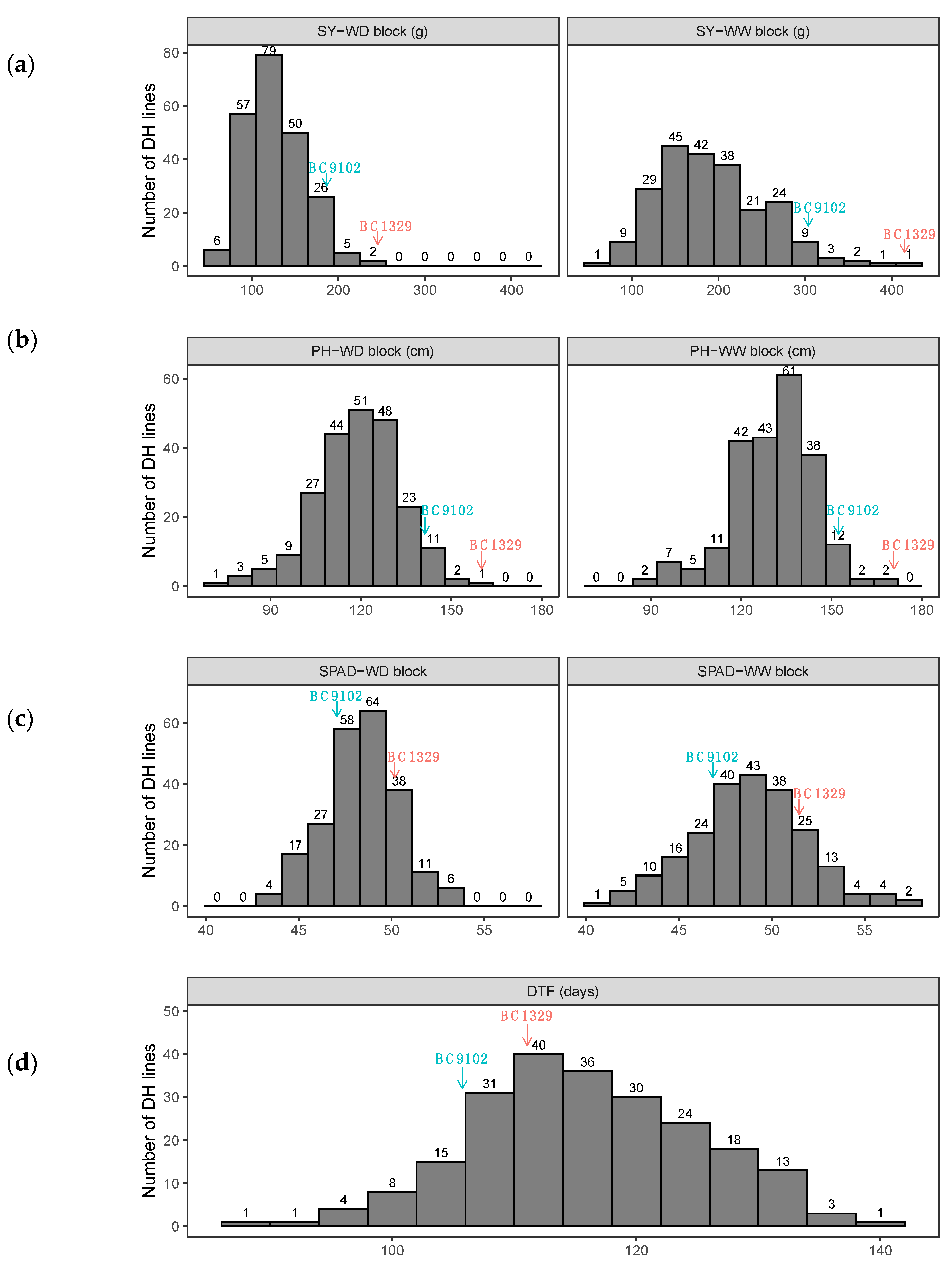
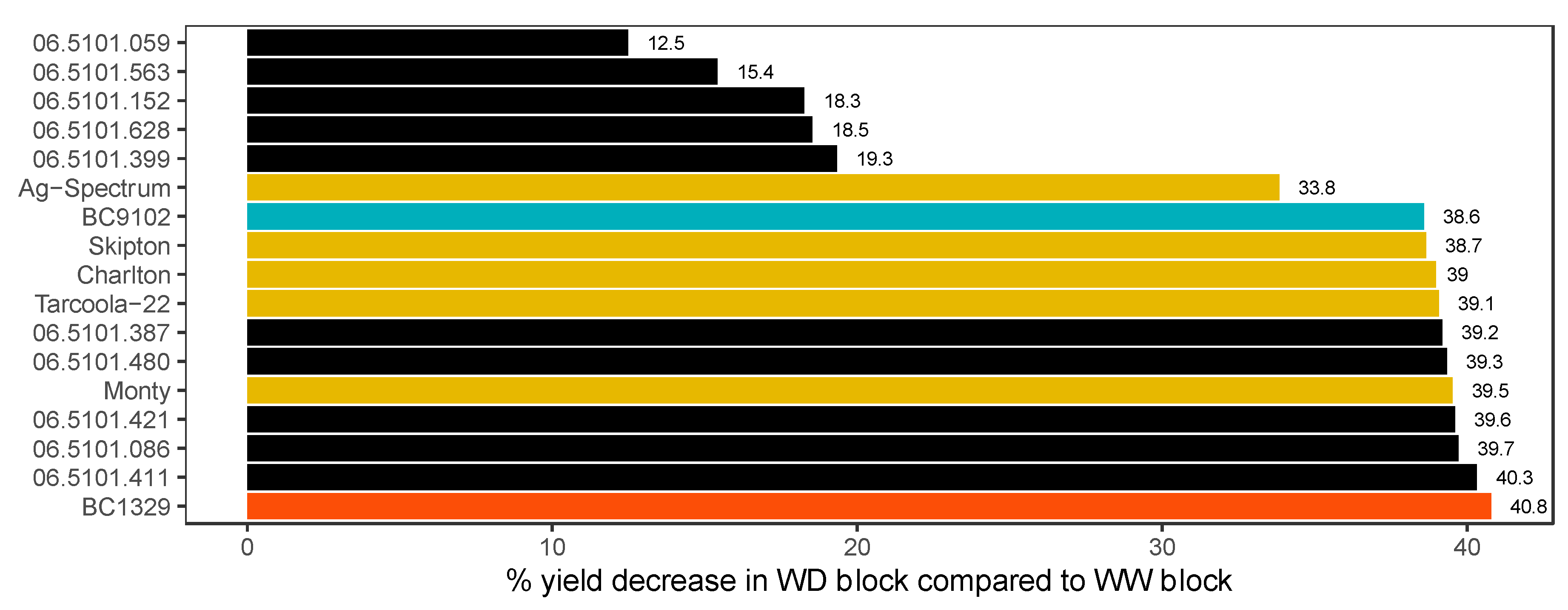
3.1. Genetic Variation in DH Lines
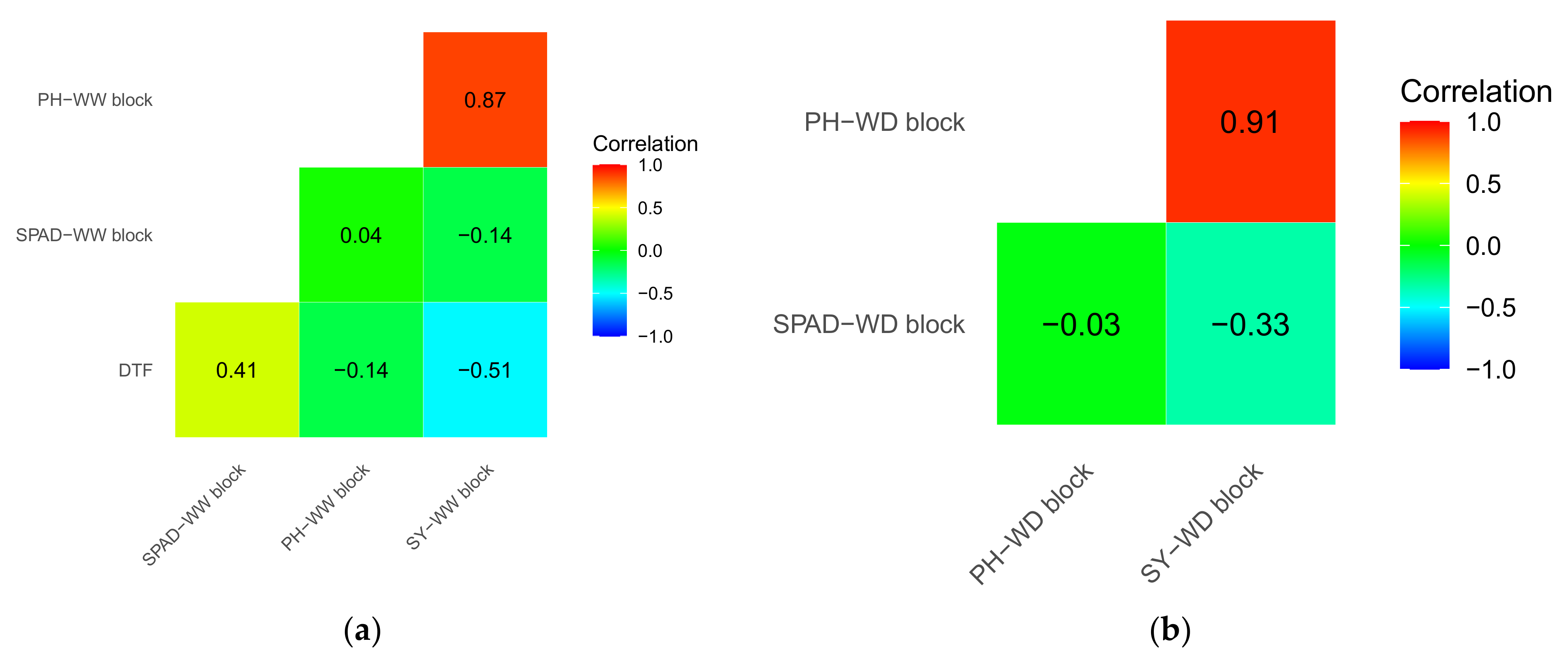
3.2. Genetic Correlations between Traits for the Two Blocks
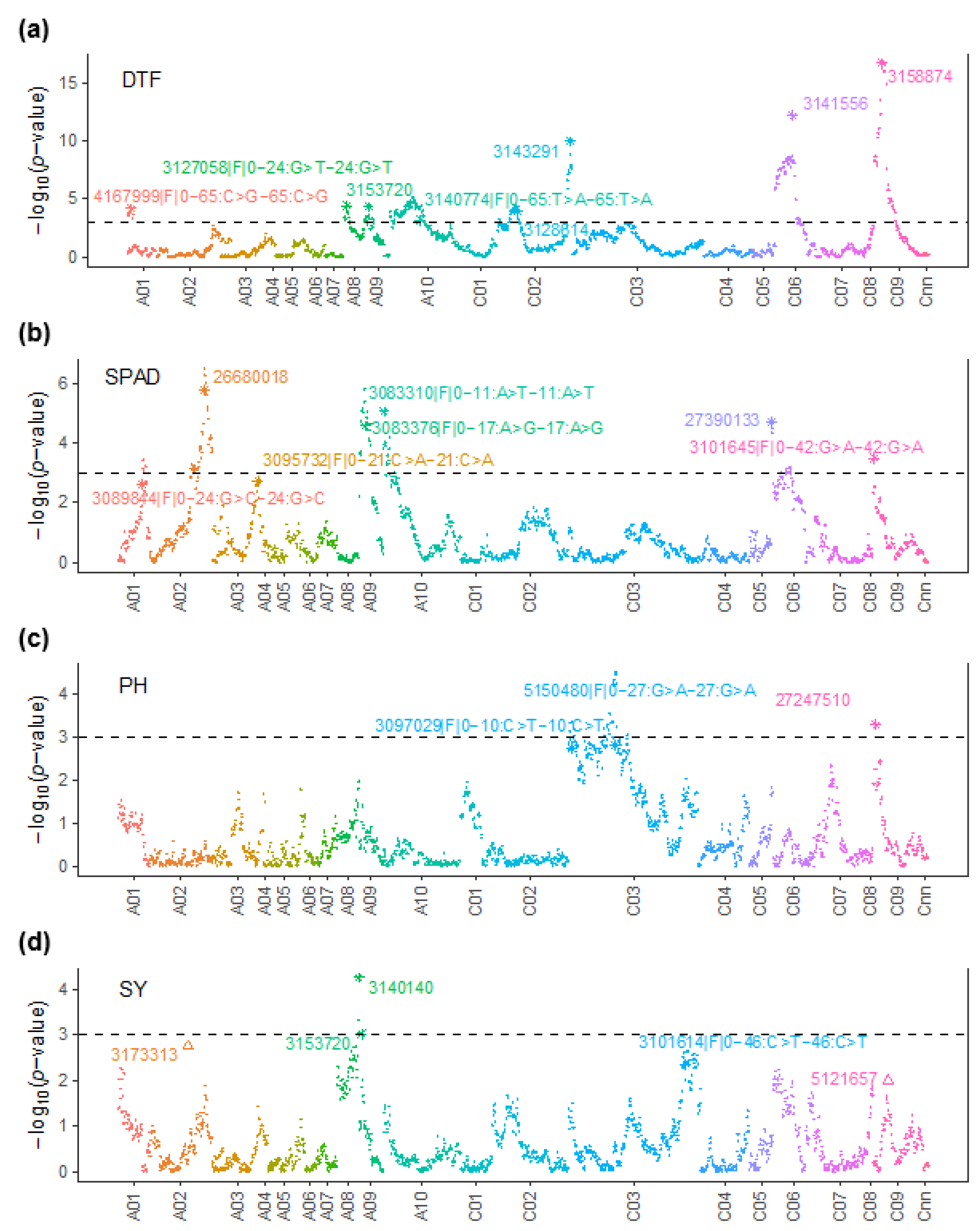
3.3. QTL Analysis Identifies QTL for Main Effects and QE Interaction Effects for Productivity Traits in Contrasting Water Regimes
3.4. Comparison of QTL across Environments
4. Discussion
4.1. Phenotypic Plasticity to Water-Deficit Conditions
4.2. Potential Proxy Traits for Selection of Genotypes with Improved Water Productivity
4.3. Identification of Stable QTL for Seed Yield and Related Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Ördög, V. Water and Nutrients in Plants. In Plant Physiology; 2011; p. 15. Available online: www.esalq.usp.br/lepse/imgs/conteudo/Plant-Physiology-by-Vince-Ordog.pdf (accessed on 30 January 2023).
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef]
- Ludlow, M.M. Strategies in Response to Water Stress. In Structural and Functional Response to Environmental Stresses: Water Shortage; Kreeb, H.K., Richter, H., Hinkley, T., Eds.; SPB Academic Press: Amsterdam, The Netherlands, 1989; pp. 269–281. [Google Scholar]
- Levitt, J. Responses of Plants to Environmental Stresses. In Water, Radiation, Salt, and Other Stresses; Academic Press: New York, NY, USA, 1980; Volume 2. [Google Scholar]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- McKay, J.K.; Richards, J.H.; Mitchell-Olds, T. Genetics of drought adaptation in Arabidopsis thaliana: I. Pleiotropy contributes to genetic correlations among ecological traits. Mol. Ecol. 2003, 12, 1137–1151. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.J.; Kirkegaard, J.A. Water-use efficiency of dryland canola in an equi-seasonal rainfall environment. Aust. J. Agric. Res. 2005, 56, 1373–1386. [Google Scholar] [CrossRef]
- Cocks, B.; Robertson, M.J.; Cawley, S. Water extraction and water use efficiency of canola in the north. In Proceedings of the Australian Research Assembly on Brassicas, Geelong, Australia, 2–5 October 2001. [Google Scholar]
- Raman, R.; Diffey, S.; Carling, J.; Cowley, R.; Kilian, A.; Luckett, D.; Raman, H. Quantitative genetic analysis of yield in an Australian Brassica napus doubled haploid population. Crop Pasture Sci. 2016, 67, 298–307. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; Mathews, K.; Diffey, S.; Salisbury, P. QTL mapping reveals genomic regions for yield based on an incremental tolerance index to drought stress and related agronomic traits in canola. Crop Pasture Sci. 2020, 71, 562–577. [Google Scholar] [CrossRef]
- McKay, J.K.; Richards, J.H.; Nemali, K.S.; Sen, S.; Mitchell-Olds, T.; Boles, S.; Juenger, T.E. Genetics of drought adaptation in Arabidopsis thaliana II. QTL analysis of a new mapping population, Kas-1 x Tsu-1. Evolution 2008, 62, 3014–3026. [Google Scholar] [CrossRef]
- Fletcher, R.S.; Mullen, J.L.; Heiliger, A.; McKay, J.K. QTL analysis of root morphology, flowering time, and yield reveals trade-offs in response to drought in Brassica napus. J. Exp. Bot. 2015, 66, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Mekonnen, M.D.; Mullen, J.L.; Arathi, H.S.; Assefa, Y.; McKay, J.K.; Byrne, P.F. Quantitative trait locus mapping for carbon isotope ratio and root pulling force in canola. Agrosyst. Geosci. Environ. 2020, 3, e20095. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; Pirathiban, R.; McVittie, B.; Sharma, N.; Liu, S.; Qiu, Y.; Zhu, A.; Killian, A.; Cullis, B.; et al. Multi-environment QTL analysis delineates a major locus associated with homoeologous exchanges for water-use efficiency and seed yield in allopolyploid Brassica napus. Plant Cell Environ. 2022, 45, 12019–12036. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, R.S.; Herrmann, D.; Mullen, J.L.; Li, Q.; Schrider, D.R.; Price, N.; Lin, J.; Grogan, K.; Kern, A.; McKay, J.K. Identification of polymorphisms associated with drought adaptation QTL in Brassica napus by resequencing. G3 Genes Genomes Genet. 2016, 6, 793–803. [Google Scholar] [CrossRef]
- Lambrides, C.J.; Chapman, S.C.; Shorter, R. Genetic variation for carbon isotope discrimination in sunflower: Association with transpiration efficiency and evidence for cytoplasmic inheritance. Crop Sci. 2004, 44, 1642–1653. [Google Scholar] [CrossRef]
- Sheshshayee, M.S.; Bindumadhava, H.; Rachaputi, N.R.; Prasad, T.G.; Udayakumar, M.; Wright, G.C.; Nigam, S.N. Leaf chlorophyll concentration relates to transpiration efficiency in peanut. Ann. Appl. Biol. 2006, 148, 7–15. [Google Scholar] [CrossRef]
- Pradhan, A.; Aher, L.; Hegde, V.; Jangid, K.K.; Rane, J. Cooler canopy leverages sorghum adaptation to drought and heat stress. Sci. Rep. 2022, 12, 4603. [Google Scholar] [CrossRef]
- Hossain, S.M.; Masle, J.; Easton, A.; Hunter, M.N.; Godwin, I.D.; Farquhar, G.D.; Lambrides, C.J. Genetic variation for leaf carbon isotope discrimination and its association with transpiration efficiency in canola (Brassica napus). Funct. Plant Biol. 2020, 47, 355–367. [Google Scholar] [CrossRef]
- Smith, A.; Lim, P.; Cullis, B. The design and analysis of multi-phase plant breeding experiments. J. Agric. Sci. 2006, 144, 393–409. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; McVittie, B.; Borg, L.; Diffey, S.; Singh Yadav, A.; Balasubramanian, S.; Farquhar, G. Genetic and physiological bases for variation in water use efficiency in canola. Food Energy Secur. 2020, 9, e237. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Richards, R.A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust. J. Plant Physiol. 1984, 11, 539–552. [Google Scholar] [CrossRef]
- Bailey, R.A.; Greenwood, J.J.D. Effects of neonicotinoids on Bees: An invalid experiment. Ecotoxicology 2018, 27, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bailey, R. Design of Comparative Experiments (Cambridge Series in Statistical and Probabilistic Mathematics); Cambridge University Press: Cambridge, UK, 2008. [Google Scholar] [CrossRef]
- Kadkol, G.; Smith, A.; Cullis, B.; Chenu, K. Variation in Australian durum wheat germplasm for productivity traits under irrigated and rainfed conditions: Genotype performance for agronomic traits and benchmarking. J. Agric. Sci. 2020, 158, 479–495. [Google Scholar] [CrossRef]
- Verbyla, A.P.; Cullis, B.R. Multivariate whole genome average interval mapping: QTL analysis for multiple traits and/or environments. Theor. Appl. Genet. 2012, 125, 933–953. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Ganesalingam, A.; Kuchel, H.; Cullis, B. Factor analytic mixed models for the provision of grower information from national crop variety testing programmes. Theor. Appl. Genet. 2015, 128, 55–72. [Google Scholar] [CrossRef]
- Butler, D.G.; Cullis, B.R.; Gilmour, A.R.; Gogel, B.J.; Thompson, R. ASReml-R Reference Manual Version 4; Technical Report; VSN International Ltd.: Hemel Hempstead, UK, 2018. [Google Scholar]
- Cullis, B.R.; Smith, A.B.; Coombes, N.E. On the design of early generation variety trials with correlated data. J. Agric. Biol. Environ. Stat. 2006, 11, 381–393. [Google Scholar] [CrossRef]
- Genesio, L.; Bassi, R.; Miglietta, F. Plants with less chlorophyll: A global change perspective. Glob. Chang. Biol. 2021, 27, 959–967. [Google Scholar] [CrossRef]
- Gu, J.; Zhou, Z.; Li, Z.; Chen, Y.; Wang, Z.; Zhang, H. Rice (Oryza sativa L.) with reduced chlorophyll content exhibit higher photosynthetic rate and efficiency, improved canopy light distribution, and greater yields than normally pigmented plants. Field Crops Res. 2017, 200, 58–70. [Google Scholar] [CrossRef]
- The International Brachypodium Initiative. Genome sequencing and analysis of the model grass Brachypodium distachyon. Nature 2010, 463, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Sakowska, K.; Alberti, G.; Genesio, L.; Peressotti, A.; Vedove, G.D.; Gianelle, D.; Colombo, R.; Rodeghiero, M.; Panigada, C.; Juszczak, R.; et al. Leaf and canopy photosynthesis of a chlorophyll deficient soybean mutant. Plant Cell Environ. 2018, 41, 1427–1437. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; Eckermann, P.; Coombes, N.; Manoli, S.; Zou, X.; Edwards, D.; Meng, J.; Prangnell, R.; Stiller, J.; et al. Genetic and physical mapping of flowering time loci in canola (Brassica napus L.). Theor. Appl. Genet. 2013, 126, 119–132. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; Qiu, Y.; Yadav, A.S.; Sureshkumar, S.; Borg, L.; Rohan, M.; Wheeler, D.; Owen, O.; Menz, I.; et al. GWAS hints at pleiotropic roles for FLOWERING LOCUS T in flowering time and yield-related traits in canola. BMC Genom. 2019, 20, 636. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Fu, D.; Mason, A.S.; Zeng, Y.; Zhao, C.; Huang, Y. In silico integration of quantitative trait loci for seed yield and yield-related traits in Brassica napus. Mol. Breed. 2014, 33, 881–894. [Google Scholar] [CrossRef]
- Raboanatahiry, N.; Chao, H.; Dalin, H.; Pu, S.; Yan, W.; Yu, L.; Wang, B.; Li, M. QTL Alignment for Seed Yield and Yield Related Traits in Brassica napus. Front. Plant Sci. 2018, 9, 1127. [Google Scholar] [CrossRef]
- Ye, J.; Liu, H.; Zhao, Z.; Xu, L.; Li, K.; Du, D. Fine mapping of the QTL cqSPDA2 for chlorophyll content in Brassica napus L. BMC Plant Biol. 2020, 20, 511. [Google Scholar] [CrossRef]
- Wang, Y.; He, Y.; Yang, M.; He, J.; Xu, P.; Shao, M.; Chu, P.; Guan, R. Fine mapping of a dominant gene conferring chlorophyll-deficiency in Brassica napus. Sci. Rep. 2016, 6, 31419. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.; Long, Y.; Liu, L.; Zhao, Y.; Tian, J.; Zhao, W.; Li, B.; Chen, L.; Chao, H.; et al. Dynamic and comparative QTL analysis for plant height in different developmental stages of Brassica napus L. Theor. Appl. Genet. 2015, 128, 1175–1192. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Alam, M.K.; Xie, M.; Yang, L.; Liu, J.; Helal, M.M.U.; Huang, J.; Cheng, X.; Liu, Y.; Tong, C.; et al. Mapping of a major QTL controlling plant height using a high-density genetic map and QTL-seq methods based on whole-genome resequencing in Brassica napus. G3 Genes Genomes Genet. 2021, 11, jkab118. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Chen, B.; Xu, K.; Gao, G.; Yan, G.; Qiao, J.; Li, J.; Li, H.; Li, L.; Xiao, X.; et al. A genome-wide association study of plant height and primary branch number in rapeseed (Brassica napus). Plant Sci. 2016, 242, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Xu, E.; Xiang, Y.; Li, Z.; Yang, Y.; Ge, X. Novel and major QTL for branch angle detected by using DH population from an exotic introgression in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2018, 131, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Tuberosa, R.; Salvi, S.; Sanguineti, M.C.; Maccaferri, M.; Giuliani, S.; Landi, P. Searching for quantitative trait loci controlling root traits in maize: A critical appraisal. Plant Soil 2003, 255, 35–54. [Google Scholar] [CrossRef]
- Lanceras, J.C.; Pantuwan, G.; Jongdee, B.; Toojinda, T. Quantitative trait loci associated with drought tolerance at reproductive stage in rice. Plant Physiol. 2004, 135, 384–399. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S. Identification of drought tolerant genotypes using physiological traits in soybean. Physiol. Mol. Biol. Plants 2019, 25, 697–711. [Google Scholar] [CrossRef] [PubMed]
- Menendez, Y.C.; Sanchez, D.H.; Snowdon, R.J.; Rondanini, D.P.; Botto, J.F. Unraveling the impact on agronomic traits of the genetic architecture underlying plant-density responses in canola. J. Exp. Bot. 2021, 72, 5426–5441. [Google Scholar] [CrossRef]
- Shi, J.; Li, R.; Zou, J.; Long, Y.; Meng, J. A dynamic and complex network regulates the heterosis of yield-correlated traits in rapeseed (Brassica napus L.). PLoS ONE 2011, 6, e21645. [Google Scholar] [CrossRef]
- Butler, D. Pedicure: Pedigree Tools. R Package Version 2.0.1. 2021. Available online: https://mmade.org/pedicure/ (accessed on 30 January 2023).
- Cullis, B.R.; Smith, A.B. The Analysis of QTL and QTL By Treatment Experiments Using Spatial Models for Marker Effects; Technical Report; University of Wollongong: Wollongong, Australia, 2016. [Google Scholar]
- Oakey, H.; Verbyla, A.; Cullis, B.R.; Wei, X.; Pitchford, W. Joint modelling of additive and non-additive (genetic line) effects in multi-environment trials. Theor. Appl. Genet. 2007, 114, 1319–1332. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 30 January 2023).
- Taylor, J.D.; Butler, D. R Package ASMap: Efficient genetic linkage map construction and diagnosis. J. Stat. Softw. 2017, 79, 1–29. [Google Scholar] [CrossRef]
- Tolhurst, D.J.; Mathews, K.L.; Smith, A.B.; Cullis, B.R. Genomic selection in multi-environment plant breeding trials using a factor analytic linear mixed model. J. Anim. Breed. Genet. 2019, 136, 279–300. [Google Scholar] [CrossRef]
- Troyanskaya, O.; Cantor, M.; Sherlock, G.; Brown, P.; Hastie, T.; Tibshirani, R.; Botstein, D.; Altman, R.B. Missing value estimation methods for DNA microarrays. Bioinformatics 2001, 17, 520–525. [Google Scholar] [CrossRef]
- VanRaden, P.M. Efficient methods to compute genomic predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef]
- Wilkinson, G.N.; Rogers, C.E. Symbolic Description of Factorial Models for Analysis of Variance. J. R. Stat. Soc. Ser. C Appl. Stat. 1973, 22, 392. [Google Scholar] [CrossRef]
- Wu, Y.; Bhat, P.R.; Close, T.J.; Lonardi, S. Efficient and accurate construction of genetic linkage maps from the minimum spanning tree of a graph. PLoS Genet. 2008, 4, e1000212. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Environment | Additive | Non-Additive | Total | VAFm (%) | |||
|---|---|---|---|---|---|---|---|---|
| (M1, %) | (M2, %) | (M1, %) | (M2, %) | (M1) | (M2) | |||
| SY | WD | 19.70 | 6.99 | 80.30 | 93.45 | 1558.57 | 1242.56 | 8.77 |
| WW | 18.33 | 14.55 | 81.67 | 86.35 | 4304.96 | 4106.61 | ||
| PH | WD | 14.58 | 1.10 | 85.42 | 98.94 | 233.00 | 208.55 | 15.83 |
| WW | 21.88 | 0.05 | 78.12 | 99.95 | 214.68 | 168.28 | ||
| SPAD | WD | 68.61 | 75.04 | 31.39 | 29.02 | 9.40 | 3.05 | 60.27 |
| WW | 57.16 | 30.72 | 42.84 | 70.94 | 15.69 | 6.92 | ||
| DTF | - | 75.44 | 35.61 | 24.56 | 64.39 | 118.77 | 35.63 | 70.00 |
| Trait | Effects | Correlation |
|---|---|---|
| SY | Additive | 0.70 |
| Total | 0.94 | |
| PH | Additive | >0.99 |
| Total | >0.99 | |
| SPAD | Additive | >0.99 |
| Total | 0.88 |
| Trait | Environment | Heritability | Minimum | Mean | Maximum | BC1329 | BC9102 |
|---|---|---|---|---|---|---|---|
| SY (g/row plot) | WD | 0.58 | 63.71 | 127.21 | 245.77 | 245.77 | 186.59 |
| WW | 0.59 | 72.80 | 192.20 | 415.02 | 415.02 | 303.78 | |
| PH (cm) | WD | 0.66 | 75.34 | 119.08 | 159.98 | 159.98 | 141.21 |
| WW | 0.69 | 87.80 | 130.60 | 170.70 | 170.70 | 152.32 | |
| SPAD (unit) | WD | 0.35 | 43.16 | 48.40 | 53.73 | 50.19 | 47.06 |
| WW | 0.49 | 41.04 | 48.83 | 56.74 | 51.48 | 46.83 | |
| DTF (days) | - | 0.81 | 89.78 | 115.99 | 138.16 | 111.06 | 105.74 |
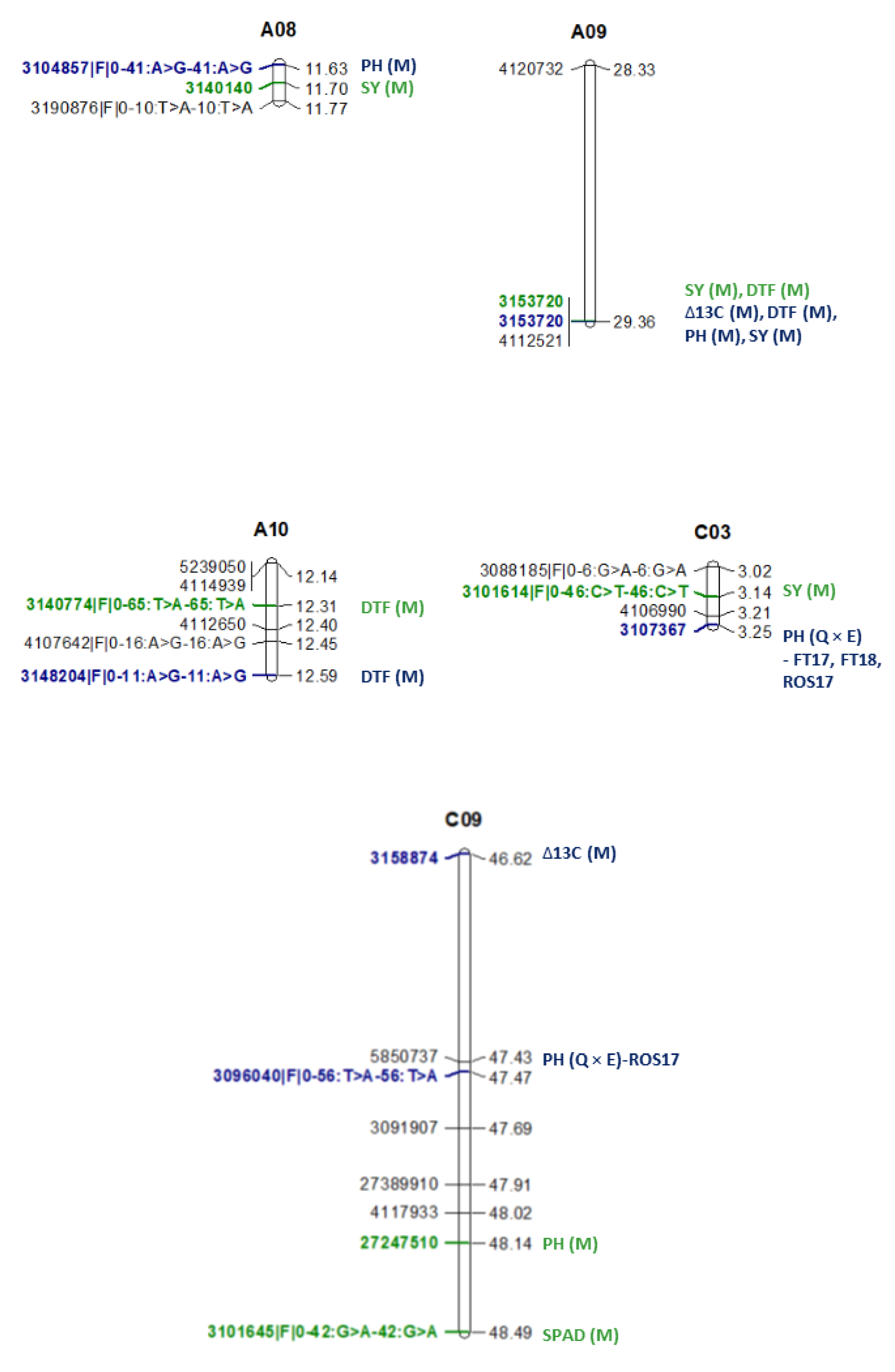
| Trait | Environment | Marker | Chromosome | Physical Map Position of ‘Top’ Marker on Darmor-bzh Genome Version 4.1 | LOD | R2 | Allelic Effect | Parental Allele |
|---|---|---|---|---|---|---|---|---|
| DTF | M | 4167999|F|0–65:C>G-65:C>G | A01 | 4,038,480 | 4.15 | 9.11 | 2.41 | BC1329 |
| DTF | M | 3127058|F|0–24:G>T-24:G>T | A08 | NA | 4.41 | 14.26 | −2.24 | BC9102 |
| *DTF | M | 3153720 | A09 | 29,356,333 | 4.31 | 17.81 | 2.19 | BC1329 |
| DTF | M | 3140774|F|0–65:T>A-65:T>A | A10 | NA | 3.15 | 17.58 | −2.44 | BC9102 |
| DTF | M | 3128614 | C02 | 9,287,096 | 3.88 | 13.04 | −2.25 | BC9102 |
| DTF | M | 3143291 | C02 | 45,636,489 | 9.95 | 26.7 | −3.31 | BC9102 |
| DTF | M | 3141556 | C06 | 27,740,738 | 12.17 | 39.12 | −4.13 | BC9102 |
| DTF | M | 3158874 | C09 | 46,623,311 | 16.66 | 48.55 | 5.06 | BC1329 |
| PH | M | 5150480|F|0–27:G>A-27:G>A | C03 | 23,396,698 | 2.82 | 3.36 | 3.46 | BC1329 |
| PH | M | ≠3097029|F|0–10:C>T-10:C>T/ 5034370|F|0–47:G>C-47:G>C | C03 | NA 57,776,378 | 2.75 | 4.68 | 3.34 | BC1329 |
| PH | M | 27247510/≠3088657 | C09 | NA/48,143,335 | 3.3 | 6 | 3.76 | BC1329 |
| SPAD | M | 3089844|F|0–24:G>C-24:G>C | A01 | 7,826,453 | 2.61 | 11.05 | 0.77 | BC1329 |
| SPAD | M | 26680018 | A02 | 23,433,061 | 5.79 | 27.15 | −1.3 | BC9102 |
| SPAD | M | 3095732|F|0–21:C>A-21:C>A | A03 | 1,882,135 | 2.72 | 9.44 | −0.78 | BC9102 |
| SPAD | M | 3083376|F|0–17:A>G-17:A>G | A09 | 26,477,098 | 4.58 | 16.46 | 1.01 | BC1329 |
| SPAD | M | 3083310|F|0–11:A>T-11:A>T | A10 | 16,253,199 | 5.06 | 24.39 | −1.14 | BC9102 |
| SPAD | M | 27390133 | C05 | 539,869 | 4.72 | 19.51 | −0.98 | BC9102 |
| SPAD | M | 3101645|F|0–42:G>A-42:G>A/ ≠3156841 | C09 | NA 48,490,657 | 3.48 | 14.94 | 0.87 | BC1329 |
| SY | WD | 3173313 | A02 | 19,738,340 | 2.76 | 1.13 | 3.91 | BC1329 |
| SY | WW | 3173313 | A02 | 19,738,340 | 2.76 | 1.13 | −12.86 | BC9102 |
| SY | M | 3140140 | A08 | 11,695,725 | 4.26 | 1.87 | 14.38 | BC1329 |
| *SY | M | 3153720 | A09 | 29,356,333 | 3.02 | 4.23 | −11.92 | BC9102 |
| SY | M | 3101614|F|0–46:C>T-46:C>T | C03 | 3,138,929 | 2.35 | 4.61 | 9.94 | BC1329 |
| SY | WD | 5121657/≠3079649 | C09 | NA/41,790,279 | 2 | 0.74 | −3.71 | BC9102 |
| SY | WW | 5121657/≠3079649 | C09 | NA/41,790,279 | 2 | 0.74 | 9.99 | BC1329 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raman, H.; Shamaya, N.; Pirathiban, R.; McVittie, B.; Raman, R.; Cullis, B.; Easton, A. Quantitative Trait Loci for Genotype and Genotype by Environment Interaction Effects for Seed Yield Plasticity to Terminal Water-Deficit Conditions in Canola (Brassica napus L.). Plants 2023, 12, 720. https://doi.org/10.3390/plants12040720
Raman H, Shamaya N, Pirathiban R, McVittie B, Raman R, Cullis B, Easton A. Quantitative Trait Loci for Genotype and Genotype by Environment Interaction Effects for Seed Yield Plasticity to Terminal Water-Deficit Conditions in Canola (Brassica napus L.). Plants. 2023; 12(4):720. https://doi.org/10.3390/plants12040720
Chicago/Turabian StyleRaman, Harsh, Nawar Shamaya, Ramethaa Pirathiban, Brett McVittie, Rosy Raman, Brian Cullis, and Andrew Easton. 2023. "Quantitative Trait Loci for Genotype and Genotype by Environment Interaction Effects for Seed Yield Plasticity to Terminal Water-Deficit Conditions in Canola (Brassica napus L.)" Plants 12, no. 4: 720. https://doi.org/10.3390/plants12040720
APA StyleRaman, H., Shamaya, N., Pirathiban, R., McVittie, B., Raman, R., Cullis, B., & Easton, A. (2023). Quantitative Trait Loci for Genotype and Genotype by Environment Interaction Effects for Seed Yield Plasticity to Terminal Water-Deficit Conditions in Canola (Brassica napus L.). Plants, 12(4), 720. https://doi.org/10.3390/plants12040720