
Targeting the Ghrelin Receptor as a Novel Therapeutic Option for Epilepsy


Abstract
:1. Introduction
2. Ghrelin and Its Receptor
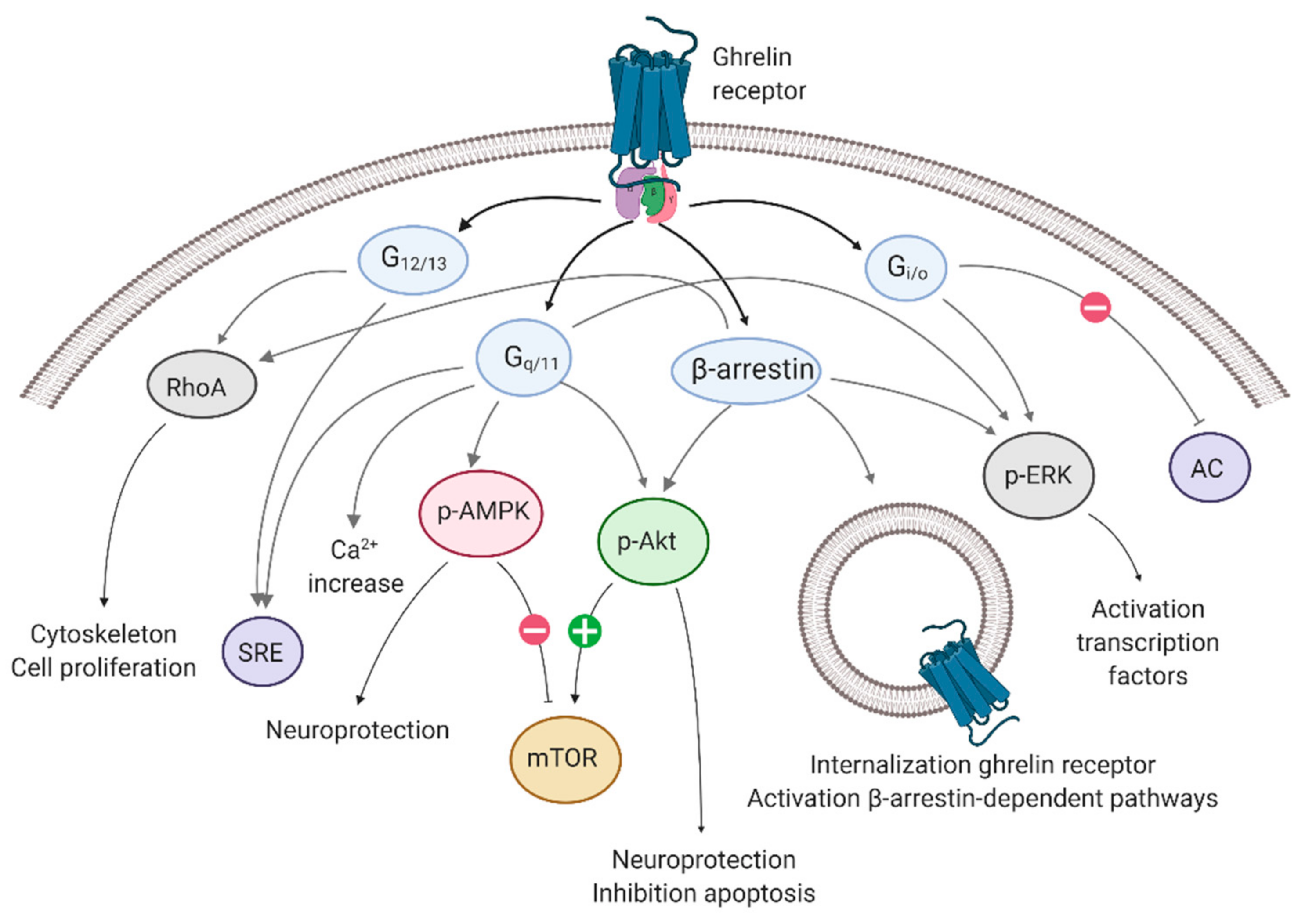
2.1. Signaling Pathways and Heteromerization Complicate Ghrelin-R Signaling
2.2. What Is Known about Ghrelin’s Central Availability?
3. Studies in Humans
3.1. Adults
3.2. Children
The Ketogenic Diet
4. Preclinical Evidence for Ghrelin as a Potential Antiseizure Drug
4.1. Ghrelin in Seizure and Status Epilepticus Rodent Models
4.2. Ghrelin Receptor Agonists
4.3. Administration of Other Ghrelin Receptor Ligands

{kind=link}
| Compound | Dose | Administration Regimen | Anticonvulsant | Animal Model | Ref |
|---|---|---|---|---|---|
| Agonists | |||||
| Macimorelin | 0.33 mg/kg | i.p 10 min prior to pilo | no | Pilocarpine i.p. rat model | [31,115] |
| Macimorelin | 5 mg/kg | i.p., 20 min prior to stimulus | yes | Acute 6- Hzmouse model | [124] |
| Macimorelin | 5 mg/kg | i.v. infusion | yes | Fully kindled 6-Hz mouse model | [124] |
| Macimorelin | 5 mg/kg | 30 min prior to SKF | yes | D1R-mediated kindling mouse model | [125] |
| Macimorelin | 5 mg/kg | 14 days, 2×/day | yes | IHKA mouse model | [126] |
| Capromorelin | 0.01–10 µM | i.h. infusion 120 min prior to pilo | yes | Pilocarpine i.h. infusion rat model | [116] |
| Hexarelin | 0.33 mg/kg | i.p. 10 min prior to pilo | yes | Pilocarpine i.p. rat model | [115] |
| Hexarelin | 0.33 mg/kg | i.p. 10 min prior to KA | no | KA i.p. rat model | [115] |
| Antagonists | |||||
| EP-80317 | 0.33 mg/kg | i.p. 10 min prior to pilo | yes/no | Pilocarpine i.p. rat model | [115] |
| EP-80317 | 0.33 mg/kg | i.p. 10 min prior to KA | no | KA i.p. rat model | [115] |
| EP-80317 | 0.33 mg/kg | i.p. 10–15 min prior to stimulus | yes | 6-Hz repeated mouse model | [131] |
| JMV-2959 | 0.33 mg/kg | i.p. 10 min prior to pilo | no | Pilocarpine i.p. rat model | [115] |
| JMV-2959 | 10 mg/kg | i.p. 20 min prior to stimulus | no | Acute 6-Hz mouse model | [124] |
| JMV-2959 | 10 mg/kg | i.v. infusion | no | 6-Hz fully kindled mice | [124] |
| JMV-2959 | 5 mg/kg | i.p. 30 min prior to SKF | no | D1R-mediated kindling mouse model | [125] |
| D-Lys-3-GHRP-6 | 1–100 µg | i.c.v. 30 min prior to stimulus | no | Amygdala kindling rat model | [133] |
| Inverse Agonists | |||||
| A778193 | 0.01–10 µM | i.h. infusion 120 min prior to pilo | yes | Pilocarpine i.h. infusion rat model | [116] |
| SP | 0.01–10 µM | i.h. infusion 120 min prior to pilo | yes | Pilocarpine i.h. infusion rat model | [116] |
| Biased Agonists | |||||
| YIL781 | 5 mg/kg | i.p. 30 min prior to SKF | no | D1R-mediated kindling mouse model | [125] |
5. Molecular Mechanisms-of-Action
5.1. Mechanisms of Ghrelin’s Anticonvulsant Action
5.2. Mechanisms of Neuroprotection
5.3. Inflammation
5.4. Oxidative Stress
6. Functional Implications of Diminished Ghrelin-R Signaling in the Context of Excitability
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Fisher, R.S.; Cross, J.H.; French, J.A.; Higurashi, N.; Hirsch, E.; Jansen, F.E.; Lagae, L.; Moshé, S.L.; Peltola, J.; Roulet Perez, E.; et al. Operational classification of seizure types by the International League Against Epilepsy: Position Paper of the ILAE Commission for Classification and Terminology. Epilepsia 2017, 58, 522–530. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Epilepsy: A Public Health Imperative. Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Nayak, C.S.; Bandyopadhyay, S. Mesial Temporal Lobe Epilepsy; Updated 5 June 2020; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Téllez-Zenteno, J.F.; Hernández-Ronquillo, L. A review of the epidemiology of temporal lobe epilepsy. Epilepsy Res. Treat. 2012, 2012, 630853. [Google Scholar] [CrossRef]
- Date, Y.; Kojima, M.; Hosoda, H.; Sawaguchi, A.; Mondal, M.S.; Suganuma, T.; Matsukura, S.; Kangawa, K.; Nakazato, M. Ghrelin, a novel growth hormone-releasing acylated peptide, is synthesized in a distinct endocrine cell type in the gastrointestinal tracts of rats and humans. Endocrinology 2000, 141, 4255–4261. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Hayashida, Y.; Nakao, N.; Nakai, N.; Nakashima, K. Testis-specific and developmentally induced expression of a ghrelin gene-derived transcript that encodes a novel polypeptide in the mouse. Biochim. Biophys. Acta 2001, 1522, 62–65. [Google Scholar] [CrossRef]
- Mori, K.; Yoshimoto, A.; Takaya, K.; Hosoda, K.; Ariyasu, H.; Yahata, K.; Mukoyama, M.; Sugawara, A.; Hosoda, H.; Kojima, M.; et al. Kidney produces a novel acylated peptide, ghrelin. FEBS Lett. 2000, 486, 213–216. [Google Scholar] [CrossRef] [Green Version]
- Volante, M.; Allìa, E.; Gugliotta, P.; Funaro, A.; Broglio, F.; Deghenghi, R.; Muccioli, G.; Ghigo, E.; Papotti, M. Expression of ghrelin and of the GH secretagogue receptor by pancreatic islet cells and related endocrine tumors. J. Clin. Endocrinol. Metab. 2002, 87, 1300–1308. [Google Scholar] [CrossRef] [PubMed]
- Cowley, M.A.; Smith, R.G.; Diano, S.; Tschöp, M.; Pronchuk, N.; Grove, K.L.; Strasburger, C.J.; Bidlingmaier, M.; Esterman, M.; Heiman, M.L.; et al. The distribution and mechanism of action of ghrelin in the CNS demonstrates a novel hypothalamic circuit regulating energy homeostasis. Neuron 2003, 37, 649–661. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Guan, J.L.; Wang, Q.P.; Uehara, K.; Yamada, S.; Goto, N.; Date, Y.; Nakazato, M.; Kojima, M.; Kangawa, K.; et al. Immunocytochemical observation of ghrelin-containing neurons in the rat arcuate nucleus. Neurosci. Lett. 2002, 321, 157–160. [Google Scholar] [CrossRef]
- Masuda, Y.; Tanaka, T.; Inomata, N.; Ohnuma, N.; Tanaka, S.; Itoh, Z.; Hosoda, H.; Kojima, M.; Kangawa, K. Ghrelin stimulates gastric acid secretion and motility in rats. Biochem. Biophys. Res. Commun. 2000, 276, 905–908. [Google Scholar] [CrossRef]
- Dornonville de la Cour, C.; Lindström, E.; Norlén, P.; Håkanson, R. Ghrelin stimulates gastric emptying but is without effect on acid secretion and gastric endocrine cells. Regul. Pept. 2004, 120, 23–32. [Google Scholar] [CrossRef]
- Colldén, G.; Tschöp, M.H.; Müller, T.D. Therapeutic Potential of Targeting the Ghrelin Pathway. Int. J. Mol. Sci. 2017, 18, 798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reimer, M.K.; Pacini, G.; Ahrén, B. Dose-dependent inhibition by ghrelin of insulin secretion in the mouse. Endocrinology 2003, 144, 916–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soeki, T.; Kishimoto, I.; Schwenke, D.O.; Tokudome, T.; Horio, T.; Yoshida, M.; Hosoda, H.; Kangawa, K. Ghrelin suppresses cardiac sympathetic activity and prevents early left ventricular remodeling in rats with myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H426–H432. [Google Scholar] [CrossRef] [Green Version]
- Khatib, M.N.; Shankar, A.; Kirubakaran, R.; Agho, K.; Simkhada, P.; Gaidhane, S.; Saxena, D.; Unnikrishnan, B.; Gode, D.; Gaidhane, A.; et al. Effect of ghrelin on mortality and cardiovascular outcomes in experimental rat and mice models of heart failure: A systematic review and meta-analysis. PLoS ONE 2015, 10, e0126697. [Google Scholar] [CrossRef]
- Dickson, S.L.; Leng, G.; Robinson, I.C. Systemic administration of growth hormone-releasing peptide activates hypothalamic arcuate neurons. Neuroscience 1993, 53, 303–306. [Google Scholar] [CrossRef]
- Dickson, S.L.; Luckman, S.M. Induction of c-fos messenger ribonucleic acid in neuropeptide Y and growth hormone (GH)-releasing factor neurons in the rat arcuate nucleus following systemic injection of the GH secretagogue, GH-releasing peptide-6. Endocrinology 1997, 138, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, P.; Zheng, H.; Smith, R.G. Ghrelin stimulation of growth hormone release and appetite is mediated through the growth hormone secretagogue receptor. Proc. Natl. Acad. Sci. USA 2004, 101, 4679–4684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perello, M.; Dickson, S.L. Ghrelin signalling on food reward: A salient link between the gut and the mesolimbic system. J. Neuroendocrinol. 2015, 27, 424–434. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Xing, T.; Wang, M.; Miao, Y.; Tang, M.; Chen, J.; Li, G.; Ruan, D.Y. Local infusion of ghrelin enhanced hippocampal synaptic plasticity and spatial memory through activation of phosphoinositide 3-kinase in the dentate gyrus of adult rats. Eur. J. Neurosci. 2011, 33, 266–275. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, H.; Xiao, K.; Yu, M.; Cui, L.; Zhu, Q.; Zhao, R.; Li, G.D.; Zhou, Y. Ghrelin administration enhances neurogenesis but impairs spatial learning and memory in adult mice. Neuroscience 2014, 257, 175–185. [Google Scholar] [CrossRef]
- Diano, S.; Farr, S.A.; Benoit, S.C.; McNay, E.C.; da Silva, I.; Horvath, B.; Gaskin, F.S.; Nonaka, N.; Jaeger, L.B.; Banks, W.A.; et al. Ghrelin controls hippocampal spine synapse density and memory performance. Nat. Neurosci. 2006, 9, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Garcia, L.; Egecioglu, E.; Studer, E.; Westberg, L.; Jerlhag, E. Ghrelin and GHS-R1A signaling within the ventral and laterodorsal tegmental area regulate sexual behavior in sexually naïve male mice. Psychoneuroendocrinology 2015, 62, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Menzies, J.R.; Skibicka, K.P.; Leng, G.; Dickson, S.L. Ghrelin, reward and motivation. Endocr. Dev. 2013, 25, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, S.J.; Xu, L.; Clarke, M.A.; Lemus, M.; Reichenbach, A.; Geenen, B.; Kozicz, T.; Andrews, Z.B. Ghrelin regulates the hypothalamic-pituitary-adrenal axis and restricts anxiety after acute stress. Biol. Psychiatry 2012, 72, 457–465. [Google Scholar] [CrossRef]
- Kojima, M.; Hosoda, H.; Date, Y.; Nakazato, M.; Matsuo, H.; Kangawa, K. Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 1999, 402, 656–660. [Google Scholar] [CrossRef]
- Xu, J.; Wang, S.; Lin, Y.; Cao, L.; Wang, R.; Chi, Z. Ghrelin protects against cell death of hippocampal neurons in pilocarpine-induced seizures in rats. Neurosci. Lett. 2009, 453, 58–61. [Google Scholar] [CrossRef]
- Lee, J.; Lim, E.; Kim, Y.; Li, E.; Park, S. Ghrelin attenuates kainic acid-induced neuronal cell death in the mouse hippocampus. J. Endocrinol. 2010, 205, 263–270. [Google Scholar] [CrossRef]
- Zhang, R.; Yang, G.; Wang, Q.; Guo, F.; Wang, H. Acylated ghrelin protects hippocampal neurons in pilocarpine-induced seizures of immature rats by inhibiting cell apoptosis. Mol. Biol. Rep. 2013, 40, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Lucchi, C.; Curia, G.; Vinet, J.; Gualtieri, F.; Bresciani, E.; Locatelli, V.; Torsello, A.; Biagini, G. Protective but not anticonvulsant effects of ghrelin and JMV-1843 in the pilocarpine model of Status epilepticus. PLoS ONE 2013, 8, e72716. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Cao, Y.; Voogd, K.; Voodg, K.; Steiner, D.F. On the processing of proghrelin to ghrelin. J. Biol. Chem. 2006, 281, 38867–38870. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Brown, M.S.; Liang, G.; Grishin, N.V.; Goldstein, J.L. Identification of the acyltransferase that octanoylates ghrelin, an appetite-stimulating peptide hormone. Cell 2008, 132, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, J.A.; Solenberg, P.J.; Perkins, D.R.; Willency, J.A.; Knierman, M.D.; Jin, Z.; Witcher, D.R.; Luo, S.; Onyia, J.E.; Hale, J.E. Ghrelin octanoylation mediated by an orphan lipid transferase. Proc. Natl. Acad. Sci. USA 2008, 105, 6320–6325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.V.; Ren, P.G.; Avsian-Kretchmer, O.; Luo, C.W.; Rauch, R.; Klein, C.; Hsueh, A.J. Obestatin, a peptide encoded by the ghrelin gene, opposes ghrelin’s effects on food intake. Science 2005, 310, 996–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toshinai, K.; Yamaguchi, H.; Sun, Y.; Smith, R.G.; Yamanaka, A.; Sakurai, T.; Date, Y.; Mondal, M.S.; Shimbara, T.; Kawagoe, T.; et al. Des-acyl ghrelin induces food intake by a mechanism independent of the growth hormone secretagogue receptor. Endocrinology 2006, 147, 2306–2314. [Google Scholar] [CrossRef]
- Portelli, J.; Coppens, J.; Demuyser, T.; Smolders, I. Des-acyl ghrelin attenuates pilocarpine-induced limbic seizures via the ghrelin receptor and not the orexin pathway. Neuropeptides 2015, 51, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gauna, C.; van de Zande, B.; van Kerkwijk, A.; Themmen, A.P.; van der Lely, A.J.; Delhanty, P.J. Unacylated ghrelin is not a functional antagonist but a full agonist of the type 1a growth hormone secretagogue receptor (GHS-R). Mol. Cell. Endocrinol. 2007, 274, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Patterson, M.; Murphy, K.G.; le Roux, C.W.; Ghatei, M.A.; Bloom, S.R. Characterization of ghrelin-like immunoreactivity in human plasma. J. Clin. Endocrinol. Metab. 2005, 90, 2205–2211. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Dave, N.; Mugundu, G.M.; Davis, H.W.; Gaylinn, B.D.; Thorner, M.O.; Tschöp, M.H.; d’Alessio, D.; Desai, P.B. The pharmacokinetics of acyl, des-acyl, and total ghrelin in healthy human subjects. Eur. J. Endocrinol. 2013, 168, 821–828. [Google Scholar] [CrossRef]
- Akamizu, T.; Takaya, K.; Irako, T.; Hosoda, H.; Teramukai, S.; Matsuyama, A.; Tada, H.; Miura, K.; Shimizu, A.; Fukushima, M.; et al. Pharmacokinetics, safety, and endocrine and appetite effects of ghrelin administration in young healthy subjects. Eur. J. Endocrinol. 2004, 150, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Docanto, M.M.; Yang, F.; Callaghan, B.; Au, C.C.; Ragavan, R.; Wang, X.; Furness, J.B.; Andrews, Z.B.; Brown, K.A. Ghrelin and des-acyl ghrelin inhibit aromatase expression and activity in human adipose stromal cells: Suppression of cAMP as a possible mechanism. Breast Cancer Res. Treat 2014, 147, 193–201. [Google Scholar] [CrossRef]
- Hopkins, A.L.; Nelson, T.A.; Guschina, I.A.; Parsons, L.C.; Lewis, C.L.; Brown, R.C.; Christian, H.C.; Davies, J.S.; Wells, T. Unacylated ghrelin promotes adipogenesis in rodent bone marrow via ghrelin O-acyl transferase and GHS-R. Sci. Rep. 2017, 7, 45541. [Google Scholar] [CrossRef]
- Murtuza, M.I.; Isokawa, M. Endogenous ghrelin-O-acyltransferase (GOAT) acylates local ghrelin in the hippocampus. J. Neurochem. 2018, 144, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.M.; Yu, H.; Palyha, O.C.; McKee, K.K.; Feighner, S.D.; Sirinathsinghji, D.J.; Smith, R.G.; van der Ploeg, L.H.; Howard, A.D. Distribution of mRNA encoding the growth hormone secretagogue receptor in brain and peripheral tissues. Brain Res. Mol. Brain Res. 1997, 48, 23–29. [Google Scholar] [CrossRef]
- Leung, P.K.; Chow, K.B.; Lau, P.N.; Chu, K.M.; Chan, C.B.; Cheng, C.H.; Wise, H. The truncated ghrelin receptor polypeptide (GHS-R1b) acts as a dominant-negative mutant of the ghrelin receptor. Cell. Signal. 2007, 19, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.D.; Feighner, S.D.; Cully, D.F.; Arena, J.P.; Liberator, P.A.; Rosenblum, C.I.; Hamelin, M.; Hreniuk, D.L.; Palyha, O.C.; Anderson, J.; et al. A receptor in pituitary and hypothalamus that functions in growth hormone release. Science 1996, 273, 974–977. [Google Scholar] [CrossRef]
- Gnanapavan, S.; Kola, B.; Bustin, S.A.; Morris, D.G.; McGee, P.; Fairclough, P.; Bhattacharya, S.; Carpenter, R.; Grossman, A.B.; Korbonits, M. The tissue distribution of the mRNA of ghrelin and subtypes of its receptor, GHS-R., in humans. J. Clin. Endocrinol. Metab. 2002, 87, 2988. [Google Scholar] [CrossRef]
- Xiao, X.; Bi, M.; Jiao, Q.; Chen, X.; Du, X.; Jiang, H. A new understanding of GHSR1a--independent of ghrelin activation. Ageing Res. Rev. 2020, 64, 101187. [Google Scholar] [CrossRef]
- Mani, B.K.; Walker, A.K.; Lopez Soto, E.J.; Raingo, J.; Lee, C.E.; Perelló, M.; Andrews, Z.B.; Zigman, J.M. Neuroanatomical characterization of a growth hormone secretagogue receptor-green fluorescent protein reporter mouse. J. Comp. Neurol. 2014, 522, 3644–3666. [Google Scholar] [CrossRef] [Green Version]
- Zigman, J.M.; Jones, J.E.; Lee, C.E.; Saper, C.B.; Elmquist, J.K. Expression of ghrelin receptor mRNA in the rat and the mouse brain. J. Comp. Neurol. 2006, 494, 528–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katayama, M.; Nogami, H.; Nishiyama, J.; Kawase, T.; Kawamura, K. Developmentally and regionally regulated expression of growth hormone secretagogue receptor mRNA in rat brain and pituitary gland. Neuroendocrinology 2000, 72, 333–340. [Google Scholar] [CrossRef]
- Kim, K.; Arai, K.; Sanno, N.; Osamura, R.Y.; Teramoto, A.; Shibasaki, T. Ghrelin and growth hormone (GH) secretagogue receptor (GHSR) mRNA expression in human pituitary adenomas. Clin. Endocrinol. 2001, 54, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Nakata, S.; Yoshino, Y.; Okita, M.; Kawabe, K.; Yamazaki, K.; Ozaki, Y.; Mori, Y.; Ochi, S.; Iga, J.I.; Ueno, S.I. Differential expression of the ghrelin-related mRNAs GHS-R1a, GHS-R1b, and MBOAT4 in Japanese patients with schizophrenia. Psychiatry Res. 2019, 272, 334–339. [Google Scholar] [CrossRef]
- Kim, M.S.; Yoon, C.Y.; Park, K.H.; Shin, C.S.; Park, K.S.; Kim, S.Y.; Cho, B.Y.; Lee, H.K. Changes in ghrelin and ghrelin receptor expression according to feeding status. Neuroreport 2003, 14, 1317–1320. [Google Scholar] [CrossRef] [PubMed]
- Evron, T.; Peterson, S.M.; Urs, N.M.; Bai, Y.; Rochelle, L.K.; Caron, M.G.; Barak, L.S. G Protein and β-arrestin signaling bias at the ghrelin receptor. J. Biol. Chem. 2014, 289, 33442–33455. [Google Scholar] [CrossRef] [Green Version]
- Sivertsen, B.; Lang, M.; Frimurer, T.M.; Holliday, N.D.; Bach, A.; Els, S.; Engelstoft, M.S.; Petersen, P.S.; Madsen, A.N.; Schwartz, T.W.; et al. Unique interaction pattern for a functionally biased ghrelin receptor agonist. J. Biol. Chem. 2011, 286, 20845–20860. [Google Scholar] [CrossRef] [Green Version]
- Damian, M.; Marie, J.; Leyris, J.P.; Fehrentz, J.A.; Verdié, P.; Martinez, J.; Banères, J.L.; Mary, S. High constitutive activity is an intrinsic feature of ghrelin receptor protein: A study with a functional monomeric GHS-R1a receptor reconstituted in lipid discs. J. Biol. Chem. 2012, 287, 3630–3641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, K.K.; Palyha, O.C.; Feighner, S.D.; Hreniuk, D.L.; Tan, C.P.; Phillips, M.S.; Smith, R.G.; van der Ploeg, L.H.; Howard, A.D. Molecular analysis of rat pituitary and hypothalamic growth hormone secretagogue receptors. Mol. Endocrinol. 1997, 11, 415–423. [Google Scholar] [CrossRef]
- Hedegaard, M.A.; Holst, B. The Complex Signaling Pathways of the Ghrelin Receptor. Endocrinology 2020, 161, bqaa020. [Google Scholar] [CrossRef]
- Camiña, J.P.; Lodeiro, M.; Ischenko, O.; Martini, A.C.; Casanueva, F.F. Stimulation by ghrelin of p42/p44 mitogen-activated protein kinase through the GHS-R1a receptor: Role of G-proteins and beta-arrestins. J. Cell. Physiol. 2007, 213, 187–200. [Google Scholar] [CrossRef]
- Offermanns, S.; Mancino, V.; Revel, J.P.; Simon, M.I. Vascular system defects and impaired cell chemokinesis as a result of Galpha13 deficiency. Science 1997, 275, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Hajicek, N.; Kozasa, T. Regulation and physiological functions of G12/13-mediated signaling pathways. Neurosignals 2009, 17, 55–70. [Google Scholar] [CrossRef] [Green Version]
- Bouzo-Lorenzo, M.; Santo-Zas, I.; Lodeiro, M.; Nogueiras, R.; Casanueva, F.F.; Castro, M.; Pazos, Y.; Tobin, A.B.; Butcher, A.J.; Camiña, J.P. Distinct phosphorylation sites on the ghrelin receptor, GHSR1a, establish a code that determines the functions of β-arrestins. Sci. Rep. 2016, 6, 22495. [Google Scholar] [CrossRef]
- Gurevich, V.V.; Gurevich, E.V. The structural basis of arrestin-mediated regulation of G-protein-coupled receptors. Pharmacol. Ther. 2006, 110, 465–502. [Google Scholar] [CrossRef] [Green Version]
- Wootten, D.; Christopoulos, A.; Marti-Solano, M.; Babu, M.M.; Sexton, P.M. Mechanisms of signalling and biased agonism in G protein-coupled receptors. Nat. Rev. Mol. Cell Biol. 2018, 19, 638–653. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.; Cygankiewicz, A.; Jensen, T.H.; Ankersen, M.; Schwartz, T.W. High constitutive signaling of the ghrelin receptor--identification of a potent inverse agonist. Mol. Endocrinol. 2003, 17, 2201–2210. [Google Scholar] [CrossRef] [PubMed]
- Holliday, N.D.; Holst, B.; Rodionova, E.A.; Schwartz, T.W.; Cox, H.M. Importance of constitutive activity and arrestin-independent mechanisms for intracellular trafficking of the ghrelin receptor. Mol. Endocrinol. 2007, 21, 3100–3112. [Google Scholar] [CrossRef] [Green Version]
- Holst, B.; Holliday, N.D.; Bach, A.; Elling, C.E.; Cox, H.M.; Schwartz, T.W. Common structural basis for constitutive activity of the ghrelin receptor family. J. Biol. Chem. 2004, 279, 53806–53817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, W.A.; Tschöp, M.; Robinson, S.M.; Heiman, M.L. Extent and direction of ghrelin transport across the blood-brain barrier is determined by its unique primary structure. J. Pharmacol. Exp. Ther. 2002, 302, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Rhea, E.M.; Salameh, T.S.; Gray, S.; Niu, J.; Banks, W.A.; Tong, J. Ghrelin transport across the blood-brain barrier can occur independently of the growth hormone secretagogue receptor. Mol. Metab. 2018, 18, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Cabral, A.; Valdivia, S.; Fernandez, G.; Reynaldo, M.; Perello, M. Divergent neuronal circuitries underlying acute orexigenic effects of peripheral or central ghrelin: Critical role of brain accessibility. J. Neuroendocrinol. 2014, 26, 542–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uriarte, M.; de Francesco, P.N.; Fernandez, G.; Cabral, A.; Castrogiovanni, D.; Lalonde, T.; Luyt, L.G.; Trejo, S.; Perello, M. Evidence Supporting a Role for the Blood-Cerebrospinal Fluid Barrier Transporting Circulating Ghrelin into the Brain. Mol. Neurobiol. 2019, 56, 4120–4134. [Google Scholar] [CrossRef]
- Uriarte, M.; de Francesco, P.N.; Fernández, G.; Castrogiovanni, D.; d’Arcangelo, M.; Imbernon, M.; Cantel, S.; Denoyelle, S.; Fehrentz, J.A.; Praetorius, J.; et al. Circulating ghrelin crosses the blood-cerebrospinal fluid barrier via growth hormone secretagogue receptor dependent and independent mechanisms. Mol. Cell. Endocrinol. 2021, 538, 111449. [Google Scholar] [CrossRef]
- Grouselle, D.; Chaillou, E.; Caraty, A.; Bluet-Pajot, M.T.; Zizzari, P.; Tillet, Y.; Epelbaum, J. Pulsatile cerebrospinal fluid and plasma ghrelin in relation to growth hormone secretion and food intake in the sheep. J. Neuroendocrinol. 2008, 20, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Collden, G.; Balland, E.; Parkash, J.; Caron, E.; Langlet, F.; Prevot, V.; Bouret, S.G. Neonatal overnutrition causes early alterations in the central response to peripheral ghrelin. Mol. Metab. 2015, 4, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.; Abizaid, A. Clarifying the Ghrelin System’s Ability to Regulate Feeding Behaviours Despite Enigmatic Spatial Separation of the GHSR and Its Endogenous Ligand. Int. J. Mol. Sci. 2017, 18, 859. [Google Scholar] [CrossRef] [Green Version]
- Nass, R.; Farhy, L.S.; Liu, J.; Pezzoli, S.S.; Johnson, M.L.; Gaylinn, B.D.; Thorner, M.O. Age-dependent decline in acyl-ghrelin concentrations and reduced association of acyl-ghrelin and growth hormone in healthy older adults. J. Clin. Endocrinol. Metab. 2014, 99, 602–608. [Google Scholar] [CrossRef] [Green Version]
- Whatmore, A.J.; Hall, C.M.; Jones, J.; Westwood, M.; Clayton, P.E. Ghrelin concentrations in healthy children and adolescents. Clin. Endocrinol. 2003, 59, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Dag, E.; Ozkan, Y.; Erman, F.; Dagli, A.F.; Kilic, N.; Sahin, I.; Karatas, F.; Yoldas, T.; Barim, A.O.; et al. Nesfatin-1 and ghrelin levels in serum and saliva of epileptic patients: Hormonal changes can have a major effect on seizure disorders. Mol. Cell. Biochem. 2009, 328, 49–56. [Google Scholar] [CrossRef]
- Dag, E.; Aydin, S.; Ozkan, Y.; Erman, F.; Dagli, A.F.; Gurger, M. Alteration in chromogranin A, obestatin and total ghrelin levels of saliva and serum in epilepsy cases. Peptides 2010, 31, 932–937. [Google Scholar] [CrossRef]
- Erkec, O.E.; Milanlıoğlu, A.; Komuroglu, A.U.; Kara, M.; Huyut, Z.; Keskin, S. Evaluation of serum ghrelin, nesfatin-1, irisin, and vasoactive intestinal peptide levels in temporal lobe epilepsy patients with and without drug resistance: A cross-sectional study. Rev. Assoc. Med. Bras. 2021, 67, 207–212. [Google Scholar] [CrossRef]
- Greco, R.; Latini, G.; Chiarelli, F.; Iannetti, P.; Verrotti, A. Leptin, ghrelin, and adiponectin in epileptic patients treated with valproic acid. Neurology 2005, 65, 1808–1809. [Google Scholar] [CrossRef]
- Varrasi, C.; Strigaro, G.; Sola, M.; Falletta, L.; Moia, S.; Prodam, F.; Cantello, R. Interictal ghrelin levels in adult patients with epilepsy. Seizure 2014, 23, 852–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berilgen, M.S.; Mungen, B.; Ustundag, B.; Demir, C. Serum ghrelin levels are enhanced in patients with epilepsy. Seizure 2006, 15, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, W.S.; Nageeb, R.S.; Elsaid, H.H. Serum and urine ghrelin in adult epileptic patients. Egypt. J. Neurol. Psychiatry Neurosurg. 2019, 55, 82. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Nishi, Y.; Yamashita, Y.; Hirata, R.; Takahashi, S.; Nagamitsu, S.; Hosoda, H.; Kangawa, K.; Kojima, M.; Matsuishi, T. Relation between circulating levels of GH, IGF-1, ghrelin and somatic growth in Rett syndrome. Brain Dev. 2014, 36, 794–800. [Google Scholar] [CrossRef]
- Deschaine, S.L.; Leggio, L. Understanding plasma treatment effect on human acyl-ghrelin concentrations. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1585–1589. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Dag, E.; Ozkan, Y.; Arslan, O.; Koc, G.; Bek, S.; Kirbas, S.; Kasikci, T.; Abasli, D.; Gokcil, Z.; et al. Time-dependent changes in the serum levels of prolactin, nesfatin-1 and ghrelin as a marker of epileptic attacks young male patients. Peptides 2011, 32, 1276–1280. [Google Scholar] [CrossRef]
- Ataie, Z.; Golzar, M.G.; Babri, S.; Ebrahimi, H.; Mohaddes, G. Does ghrelin level change after epileptic seizure in rats? Seizure 2011, 20, 347–349. [Google Scholar] [CrossRef] [Green Version]
- Prodam, F.; Bellone, S.; Casara, G.; de Rienzo, F.; Grassino, E.C.; Bonsignori, I.; Demarchi, I.; Rapa, A.; Radetti, G.; Bona, G. Ghrelin levels are reduced in prepubertal epileptic children under treatment with carbamazepine or valproic acid. Epilepsia 2010, 51, 312–315. [Google Scholar] [CrossRef]
- Cansu, A.; Serdaroglu, A.; Camurdan, O.; Hirfanoglu, T.; Cinaz, P. Serum Insulin, Cortisol, Leptin, Neuropeptide Y, Galanin and Ghrelin Levels in Epileptic Children Receiving Valproate. Horm. Res. Paediatr. 2011, 76, 65–71. [Google Scholar] [CrossRef]
- Taskin, E.; Atli, B.; Kiliç, M.; Sari, Y.; Aydin, S. Serum, urine, and saliva levels of ghrelin and obestatin pre- and post-treatment in pediatric epilepsy. Pediatr. Neurol. 2014, 51, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Çiçek, N.P.; Kamaşak, T.; Serin, M.; Okten, A.; Alver, A.; Cansu, A. The effects of valproate and topiramate use on serum insulin, leptin, neuropeptide Y and ghrelin levels in epileptic children. Seizure 2018, 58, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Cansu, A.; Serdaroglu, A.; Cinaz, P. Serum insulin, cortisol, leptin, neuropeptide Y, galanin and ghrelin levels in epileptic children receiving oxcarbazepine. Eur. J. Paediatr. Neurol. 2011, 15, 527–531. [Google Scholar] [CrossRef]
- Tokgoz, H.; Aydin, K.; Oran, B.; Kiyici, A. Plasma leptin, neuropeptide Y, ghrelin, and adiponectin levels and carotid artery intima media thickness in epileptic children treated with valproate. Childs Nerv. Syst. 2012, 28, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Ozcelik, A.A.; Serdaroglu, A.; Bideci, A.; Arhan, E.; Soysal, Ş.; Demir, E.; Gücüyener, K. The effect of topiramate on body weight and ghrelin, leptin, and neuropeptide-Y levels of prepubertal children with epilepsy. Pediatr. Neurol. 2014, 51, 220–224. [Google Scholar] [CrossRef]
- Cansu, A.; Yesilkaya, E.; Serdaroglu, A.; Camurdan, O.; Hirfanoglu, T.L.; Karaoglu, A.; Bideci, A.; Cinaz, P. The Effects of Oxcarbazepine and Valproate Therapies on Growth in Children with Epilepsy. Endocr. Res. 2012, 37, 163–174. [Google Scholar] [CrossRef]
- Gungor, S.; Yücel, G.; Akinci, A.; Tabel, Y.; Ozerol, I.H.; Yologlu, S. The role of ghrelin in weight gain and growth in epileptic children using valproate. J. Child Neurol. 2007, 22, 1384–1388. [Google Scholar] [CrossRef]
- Hasaneen, B.; Salem, N.A.; El Sallab, S.; Elgaml, D.; Elhelaly, R. Body weight, body composition, and serum ghrelin in epileptic children receiving levetiracetam monotherapy. Egypt. Pediatr. Assoc. Gaz. 2016, 64, 154–159. [Google Scholar] [CrossRef] [Green Version]
- Marchiò, M.; Roli, L.; Giordano, C.; Trenti, T.; Guerra, A.; Biagini, G. Decreased ghrelin and des-acyl ghrelin plasma levels in patients affected by pharmacoresistant epilepsy and maintained on the ketogenic diet. Clin. Nutr. 2019, 38, 954–957. [Google Scholar] [CrossRef] [PubMed]
- Marchiò, M.; Roli, L.; Lucchi, C.; Costa, A.M.; Borghi, M.; Iughetti, L.; Trenti, T.; Guerra, A.; Biagini, G. Ghrelin Plasma Levels After 1 Year of Ketogenic Diet in Children With Refractory Epilepsy. Front. Nutr. 2019, 6, 112. [Google Scholar] [CrossRef] [Green Version]
- De Amicis, R.; Leone, A.; Lessa, C.; Foppiani, A.; Ravella, S.; Ravasenghi, S.; Trentani, C.; Ferraris, C.; Veggiotti, P.; de Giorgis, V.; et al. Long-Term Effects of a Classic Ketogenic Diet on Ghrelin and Leptin Concentration: A 12-Month Prospective Study in a Cohort of Italian Children and Adults with GLUT1-Deficiency Syndrome and Drug Resistant Epilepsy. Nutrients 2019, 11, 1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, C.; Marchiò, M.; Timofeeva, E.; Biagini, G. Neuroactive peptides as putative mediators of antiepileptic ketogenic diets. Front. Neurol. 2014, 5, 63. [Google Scholar] [CrossRef]
- Obay, B.D.; Tasdemir, E.; Tümer, C.; Bilgin, H.M.; Sermet, A. Antiepileptic effects of ghrelin on pentylenetetrazole-induced seizures in rats. Peptides 2007, 28, 1214–1219. [Google Scholar] [CrossRef]
- Ghahramanian Golzar, M.; Babri, S.; Ataie, Z.; Ebrahimi, H.; Mirzaie, F.; Mohaddes, G. NPY Receptors Blockade Prevents Anticonvulsant Action of Ghrelin in the Hippocampus of Rat. Adv. Pharm. Bull. 2013, 3, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Ataie, Z.; Babri, S.; Ghahramanian Golzar, M.; Ebrahimi, H.; Mirzaie, F.; Mohaddes, G. GABAB Receptor Blockade Prevents Antiepileptic Action of Ghrelin in the Rat Hippocampus. Adv. Pharm. Bull. 2013, 3, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Zendehdel, M.; Kaboutari, J.; Ghadimi, D.; Hassanpour, S. The Antiepileptic Effect of Ghrelin during Different Phases of the Estrous Cycle in PTZ-Induced Seizures in Rat. Int. J. Pept. Res. Ther. 2014, 20, 511–517. [Google Scholar] [CrossRef]
- Babri, S.; Amani, M.; Mohaddes, G.; Mirzaei, F.; Mahmoudi, F. Effects of intrahippocampal injection of ghrelin on spatial memory in PTZ-induced seizures in male rats. Neuropeptides 2013, 47, 355–360. [Google Scholar] [CrossRef]
- Keezer, M.R.; Sisodiya, S.M.; Sander, J.W. Comorbidities of epilepsy: Current concepts and future perspectives. Lancet Neurol. 2016, 15, 106–115. [Google Scholar] [CrossRef]
- Ergul Erkec, O.; Algul, S.; Kara, M. Evaluation of ghrelin, nesfatin-1 and irisin levels of serum and brain after acute or chronic pentylenetetrazole administrations in rats using sodium valproate. Neurol. Res. 2018, 40, 923–929. [Google Scholar] [CrossRef] [PubMed]
- Turkmen, A.Z.; Nurten, A. Investigation of Ghrelin Levels in Antimuscarinic Induced Convulsions in Fasted Animals after Food Intake. Bezmialem Sci. 2020, 8, 138–143. [Google Scholar] [CrossRef]
- Aslan, A.; Yildirim, M.; Ayyildiz, M.; Güven, A.; Agar, E. The role of nitric oxide in the inhibitory effect of ghrelin against penicillin-induced epileptiform activity in rat. Neuropeptides 2009, 43, 295–302. [Google Scholar] [CrossRef]
- Oztas, B.; Sahin, D.; Kir, H.; Kuskay, S.; Ates, N. Effects of leptin, ghrelin and neuropeptide y on spike-wave discharge activity and certain biochemical parameters in WAG/Rij rats with genetic absence epilepsy. J. Neuroimmunol. 2021, 351, 577454. [Google Scholar] [CrossRef]
- Biagini, G.; Torsello, A.; Marinelli, C.; Gualtieri, F.; Vezzali, R.; Coco, S.; Bresciani, E.; Locatelli, V. Beneficial effects of desacyl-ghrelin, hexarelin and EP-80317 in models of status epilepticus. Eur. J. Pharmacol. 2011, 670, 130–136. [Google Scholar] [CrossRef]
- Portelli, J.; Thielemans, L.; ver Donck, L.; Loyens, E.; Coppens, J.; Aourz, N.; Aerssens, J.; Vermoesen, K.; Clinckers, R.; Schallier, A.; et al. Inactivation of the constitutively active ghrelin receptor attenuates limbic seizure activity in rodents. Neurotherapeutics 2012, 9, 658–672. [Google Scholar] [CrossRef] [Green Version]
- Oztas, B.; Sahin, D.; Kir, H.; Eraldemir, F.C.; Musul, M.; Kuskay, S.; Ates, N. The effect of leptin, ghrelin, and neuropeptide-Y on serum Tnf-A, Il-1β, Il-6, Fgf-2, galanin levels and oxidative stress in an experimental generalized convulsive seizure model. Neuropeptides 2017, 61, 31–37. [Google Scholar] [CrossRef]
- Kilinc, E.; Gunes, H. Modulatory effects of neuropeptides on pentylenetetrazol-induced epileptic seizures and neuroinflammation in rats. Rev. Assoc. Med. Bras. 2019, 65, 1188–1192. [Google Scholar] [CrossRef] [Green Version]
- Arslan, G.; Ayyildiz, M.; Agar, E. The interaction between ghrelin and cannabinoid systems in penicillin-induced epileptiform activity in rats. Neuropeptides 2014, 48, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Guerlavais, V.; Boeglin, D.; Mousseaux, D.; Oiry, C.; Heitz, A.; Deghenghi, R.; Locatelli, V.; Torsello, A.; Ghé, C.; Catapano, F.; et al. New active series of growth hormone secretagogues. J. Med. Chem. 2003, 46, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.M.; Biller, B.M.K.; Korbonits, M.; Popovic, V.; Luger, A.; Strasburger, C.J.; Chanson, P.; Medic-Stojanoska, M.; Schopohl, J.; Zakrzewska, A.; et al. Macimorelin as a Diagnostic Test for Adult GH Deficiency. J. Clin. Endocrinol. Metab. 2018, 103, 3083–3093. [Google Scholar] [CrossRef] [PubMed]
- Klaus, B.; Sachse, R.; Ammer, N.; Kelepouris, N.; Ostrow, V. Safety, tolerability, pharmacokinetics, and pharmacodynamics of macimorelin in healthy adults: Results of a single-dose, randomized controlled study. Growth Horm. IGF Res. 2020, 52, 101321. [Google Scholar] [CrossRef] [PubMed]
- M’Kadmi, C.; Leyris, J.P.; Onfroy, L.; Galés, C.; Saulière, A.; Gagne, D.; Damian, M.; Mary, S.; Maingot, M.; Denoyelle, S.; et al. Agonism, Antagonism, and Inverse Agonism Bias at the Ghrelin Receptor Signaling. J. Biol. Chem. 2015, 290, 27021–27039. [Google Scholar] [CrossRef] [Green Version]
- Coppens, J.; Aourz, N.; Walrave, L.; Fehrentz, J.A.; Martinez, J.; de Bundel, D.; Portelli, J.; Smolders, I. Anticonvulsant effect of a ghrelin receptor agonist in 6Hz corneally kindled mice. Epilepsia 2016, 57, e195–e199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckinx, A.; van den Herrewegen, Y.; Pierre, A.; Cottone, E.; Ben Haj Salah, K.; Fehrentz, J.A.; Kooijman, R.; de Bundel, D.; Smolders, I. Differential Effects of a Full and Biased Ghrelin Receptor Agonist in a Mouse Kindling Model. Int. J. Mol. Sci. 2019, 20, 2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckinx, A.; Pierre, A.; van den Herrewegen, Y.; Guenther, E.; Gerlach, M.; van Laethem, G.; Kooijman, R.; de Bundel, D.; Smolders, I. Translational potential of the ghrelin receptor agonist macimorelin for seizure suppression in pharmacoresistant epilepsy. Eur. J. Neurol. 2021, 28, 3100–3112. [Google Scholar] [CrossRef] [PubMed]
- Carpino, P.A.; Lefker, B.A.; Toler, S.M.; Pan, L.C.; Hadcock, J.R.; Murray, M.C.; Cook, E.R.; DiBrino, J.N.; DeNinno, S.L.; Chidsey-Frink, K.L.; et al. Discovery and biological characterization of capromorelin analogues with extended half-lives. Bioorg. Med. Chem. Lett. 2002, 12, 3279–3282. [Google Scholar] [CrossRef]
- Zollers, B.; Rhodes, L.; Heinen, E. Capromorelin oral solution (ENTYCE®) increases food consumption and body weight when administered for 4 consecutive days to healthy adult Beagle dogs in a randomized, masked, placebo controlled study. BMC Vet. Res. 2017, 13, 10. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, L.; Zollers, B.; Wofford, J.A.; Heinen, E. Capromorelin: A ghrelin receptor agonist and novel therapy for stimulation of appetite in dogs. Vet. Med. Sci. 2018, 4, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Deghenghi, R.; Cananzi, M.M.; Torsello, A.; Battisti, C.; Muller, E.E.; Locatelli, V. GH-releasing activity of Hexarelin, a new growth hormone releasing peptide, in infant and adult rats. Life Sci. 1994, 54, 1321–1328. [Google Scholar] [CrossRef]
- Giordano, C.; Costa, A.M.; Lucchi, C.; Leo, G.; Brunel, L.; Fehrentz, J.A.; Martinez, J.; Torsello, A.; Biagini, G. Progressive Seizure Aggravation in the Repeated 6-Hz Corneal Stimulation Model Is Accompanied by Marked Increase in Hippocampal p-ERK1/2 Immunoreactivity in Neurons. Front. Cell. Neurosci. 2016, 10, 281. [Google Scholar] [CrossRef] [Green Version]
- Lucchi, C.; Costa, A.M.; Giordano, C.; Curia, G.; Piat, M.; Leo, G.; Vinet, J.; Brunel, L.; Fehrentz, J.A.; Martinez, J.; et al. Involvement of PPARγ in the Anticonvulsant Activity of EP-80317, a Ghrelin Receptor Antagonist. Front. Pharmacol. 2017, 8, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azimzadeh, M.; Beheshti, S. Antagonism of the ghrelin receptor type 1a in the rat brain induces status epilepticus in an electrical kindling model of epilepsy. Psychopharmacology 2021, 238, 1–9. [Google Scholar] [CrossRef]
- Barker-Haliski, M.; White, H.S. Glutamatergic Mechanisms Associated with Seizures and Epilepsy. Cold Spring Harb. Perspect. Med. 2015, 5, a022863. [Google Scholar] [CrossRef] [Green Version]
- Viviani, B.; Bartesaghi, S.; Gardoni, F.; Vezzani, A.; Behrens, M.M.; Bartfai, T.; Binaglia, M.; Corsini, E.; di Luca, M.; Galli, C.L.; et al. Interleukin-1beta enhances NMDA receptor-mediated intracellular calcium increase through activation of the Src family of kinases. J. Neurosci. 2003, 23, 8692–8700. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; French, J.; Bartfai, T.; Baram, T.Z. The role of inflammation in epilepsy. Nat. Rev. Neurol. 2011, 7, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, H.; Jin, S.; Wang, J.; Zhang, G.; Kawanokuchi, J.; Kuno, R.; Sonobe, Y.; Mizuno, T.; Suzumura, A. Tumor necrosis factor-alpha induces neurotoxicity via glutamate release from hemichannels of activated microglia in an autocrine manner. J. Biol. Chem. 2006, 281, 21362–21368. [Google Scholar] [CrossRef] [Green Version]
- Rana, A.; Musto, A.E. The role of inflammation in the development of epilepsy. J. Neuroinflamm. 2018, 15, 144. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Zhao, J.; Yang, J.; Zhang, Z.; Du, J.; Tang, C. Therapeutic effects of ghrelin on endotoxic shock in rats. Eur. J. Pharmacol. 2003, 473, 171–176. [Google Scholar] [CrossRef]
- Han, K.; Wang, Q.Y.; Wang, C.X.; Luan, S.Y.; Tian, W.P.; Wang, Y.; Zhang, R.Y. Ghrelin improves pilocarpine-induced cerebral cortex inflammation in epileptic rats by inhibiting NF-κ B and TNF-α. Mol. Med. Rep. 2018, 18, 3563–3568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, E.J.; Jeong, J.H.; Chung, Y.H.; Kim, W.K.; Ko, K.H.; Bach, J.H.; Hong, J.S.; Yoneda, Y.; Kim, H.C. Role of oxidative stress in epileptic seizures. Neurochem. Int. 2011, 59, 122–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obay, B.D.; Taşdemir, E.; Tümer, C.; Bilgin, H.; Atmaca, M. Dose dependent effects of ghrelin on pentylenetetrazole-induced oxidative stress in a rat seizure model. Peptides 2008, 29, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Kern, A.; Mavrikaki, M.; Ullrich, C.; Albarran-Zeckler, R.; Brantley, A.F.; Smith, R.G. Hippocampal Dopamine/DRD1 Signaling Dependent on the Ghrelin Receptor. Cell 2015, 163, 1176–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisani, A.; Bonsi, P.; Martella, G.; de Persis, C.; Costa, C.; Pisani, F.; Bernardi, G.; Calabresi, P. Intracellular calcium increase in epileptiform activity: Modulation by levetiracetam and lamotrigine. Epilepsia 2004, 45, 719–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, S.; Sun, D.; Limbrick, D.; Rafiq, A.; DeLorenzo, R.J. Epileptogenesis induces long-term alterations in intracellular calcium release and sequestration mechanisms in the hippocampal neuronal culture model of epilepsy. Cell Calcium 2001, 30, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Cavalier, M.; Crouzin, N.; Ben Sedrine, A.; de Jesus Ferreira, M.C.; Guiramand, J.; Cohen-Solal, C.; Fehrentz, J.A.; Martinez, J.; Barbanel, G.; Vignes, M. Involvement of PKA and ERK pathways in ghrelin-induced long-lasting potentiation of excitatory synaptic transmission in the CA1 area of rat hippocampus. Eur. J. Neurosci. 2015, 42, 2568–2576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, M.T.; Herman, M.A.; Cote, D.M.; Ryabinin, A.E.; Roberto, M. Ghrelin increases GABAergic transmission and interacts with ethanol actions in the rat central nucleus of the amygdala. Neuropsychopharmacology 2013, 38, 364–375. [Google Scholar] [CrossRef] [PubMed]
- Mir, J.F.; Zagmutt, S.; Lichtenstein, M.P.; García-Villoria, J.; Weber, M.; Gracia, A.; Fabriàs, G.; Casas, J.; López, M.; Casals, N.; et al. Ghrelin Causes a Decline in GABA Release by Reducing Fatty Acid Oxidation in Cortex. Mol. Neurobiol. 2018, 55, 7216–7228. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Xu, F.; Yu, M.; Qiao, Z.; Zhou, Y. Selectively increasing GHS-R1a expression in dCA1 excitatory/inhibitory neurons have opposite effects on memory encoding. Mol. Brain 2021, 14, 157. [Google Scholar] [CrossRef] [PubMed]

| ASD | Ghrelin Form | Controls (pg/mL) | * | Epilepsy Patients (pg/mL) | ** | Ref | |
|---|---|---|---|---|---|---|---|
| Focal Seizures | Generalized Seizures | ||||||
| VPA, PHT, CBZ | Total | 93 | ↑ | 234 | 134 | ↓ | [85] |
| VPA, PHT, CBZ | DAG AG | DAG: 585 AG: 46 | ↓ | DAG: 439 AG: 35 | DAG: 267 AG: 23 | ↓ | [80] |
| VPA, PHT, CBZ | Total | 700 | ↓ | 500 | 300 | ↓ | [81] |
| VPA | N/A | 381 | = | 364 (both types) | / | [83] | |
| VPA, CBZ | DAG AG | DAG: 196 AG: 7 | = | DAG: 207-239, AG: 7-22 | DAG: 250 AG: 8 | = | [84] |
| N/A | DAG AG | DAG 242 AG: 13 | ↑ | DAG: 238 AG: 14.5 | DAG: 245 AG: 19 | ↑ | [86] |
| N/A | N/A | 1320 | ↓ | TLE: 1010 | DR-TLE: 910 | / | [82] |
| ASD | Ghrelin Form | Controls (pg/mL) | * | Epilepsy Patients (pg/mL) | ** | Age (Years) | Ref | |
|---|---|---|---|---|---|---|---|---|
| VPA, CBZ | Total | 554 | ↓ | VPA: 381 | CBZ: 283 | ↓ | 5 | [91] |
| VPA, TPM | N/A | 267 | = | VPA: 240 | TPM: 267 | = | 6–15 | [94] |
| VPA | DAG AG | DAG: 446 AG: 45 | ↓ | Pretreatment: DAG: 420 AG: 36 | Post-treatment: DAG: 459 AG: 51 | ↑ | 9 | [93] |
| VPA | N/A | 333 | ↓ | Pretreatment: 355 | Post-treatment: 263 | ↓ | 11 | [92] |
| Int. | Ghrelin Form | Baseline (pg/mL) | T1 | T2 | T3 | T4 | * | Age (Years) | Ref |
|---|---|---|---|---|---|---|---|---|---|
| OXC | N/A | 327 | 6 m: 306 | 18 m: 320 | / | / | = | 9 | [95] |
| OXC | N/A | 310 | 6 m: 288 | 18 m: 345 | / | / | = | 13 | [98] |
| VPA | N/A | 18 | 6 m: 18 | 12 m: 18 | / | / | = | 9 | [96] |
| VPA | N/A | 334 | 6 m: 275 | 18 m: 245 | / | / | ↓ | 14 | [98] |
| VPA | N/A | 1.37 | 6 m: 2.19 | / | / | / | ↑ | 8 | [99] |
| TPM | N/A | 1121 | 3 m: 1184 | 6 m: 1292 | / | / | = | 8 | [97] |
| LEV | N/A | 1900 | 6 m: 2950 | / | / | / | = | 7 | [100] |
| KD | DAG AG | DAG: ±160 AG: ±250 | 15 d: DAG: ±110 AG: ±210 | 30 d: DAG: ±100 AG: ±140 | 90 d: DAG: ±140 AG: ±110 | / | ↓ | 7 | [101] |
| KD | AG | ±400 | 15 d: ±250 | 30 d: ±200 | 90 d: ±200 | 1 y: ±200 | ↓ | 6 | [102] |
| KD | N/A | 20 | 6 m: 19 | 12 m: 19 | / | / | / | 8 | [103] |
| Dose | Administration Regimen | Anticonvulsant | Animal Model | Ref |
|---|---|---|---|---|
| 0.02–0.08 mg/kg | i.p. 30 min prior to PTZ | yes | PTZ i.p. rat model | [105,117] |
| 0.08 mg/kg | i.p. 30 min prior to PTZ | no | PTZ i.p. rat model | [118] |
| 0.3 nmol/µL | i.h. infusion 1 x 30 min prior to PTZ or 10 days | yes | PTZ i.p. rat model | [106,107] |
| 0.08 mg/kg | i.c.v. 30 min prior to PTZ | yes | PTZ i.p. rat model (female rats) | [108] |
| 0.5, 1 and 2 µg | i.c.v. 30 min after pen | yes | Intracortical penicillin rat model | [113,119] |
| 0.08 mg/kg | i.p., immediate assessment | no | WAG/Rij rat model | [114] |
| 0.01–10 µM | i.h. infusion, 120 min prior to pilo | yes | Pilocarpine i.h. infusion rat model | [116] |
| 1.5 mg/kg | i.p. 10 min prior to pilo | no | Pilocarpine i.p. rat model | [31,115] |
| 1.5 mg/kg | i.p. 10 min prior to KA | no | KA i.p. rat model | [115] |
| 0.08 mg/kg | i.p. 30 min prior to KA, and 24 h after KA | yes | KA i.p. mouse model | [29] |
| 1.8 mg/kg | i.p. 30 min prior to pilo | yes | Pilocarpine tail infusion mouse model | [116] |
| Desacyl Ghrelin | ||||
|---|---|---|---|---|
| Dose | Administration Regimen | Anticonvulsant | Animal Model | Ref |
| 1.5 mg/kg | i.p. 10 min prior to pilo | yes/no (p = 0.07) | Pilocarpine i.p. rat model | [115] |
| 1.5 mg/kg | i.p. 10 min prior to KA | yes | KA i.p. rat model | [115] |
| 1–10 µM | i.c.v. 2 h prior to pilo | yes | Pilocarpine i.c.v. rat model | [37] |
| 3/5 mg/kg | i.p. 30 min prior to pilo | yes | Pilocarpine tail infusion mouse model | [37] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buckinx, A.; De Bundel, D.; Kooijman, R.; Smolders, I. Targeting the Ghrelin Receptor as a Novel Therapeutic Option for Epilepsy. Biomedicines 2022, 10, 53. https://doi.org/10.3390/biomedicines10010053
Buckinx A, De Bundel D, Kooijman R, Smolders I. Targeting the Ghrelin Receptor as a Novel Therapeutic Option for Epilepsy. Biomedicines. 2022; 10(1):53. https://doi.org/10.3390/biomedicines10010053
Chicago/Turabian StyleBuckinx, An, Dimitri De Bundel, Ron Kooijman, and Ilse Smolders. 2022. "Targeting the Ghrelin Receptor as a Novel Therapeutic Option for Epilepsy" Biomedicines 10, no. 1: 53. https://doi.org/10.3390/biomedicines10010053
APA StyleBuckinx, A., De Bundel, D., Kooijman, R., & Smolders, I. (2022). Targeting the Ghrelin Receptor as a Novel Therapeutic Option for Epilepsy. Biomedicines, 10(1), 53. https://doi.org/10.3390/biomedicines10010053