Sperm Proteomics Analysis of Diabetic Induced Male Rats as Influenced by Ficus carica Leaf Extract

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of FC Aqueous Extract
2.2. Experimental Animal
2.3. Antihyperglycemic Activity
2.4. Sperm Quality Analysis
2.5. Sperm Proteomic Analysis
2.6. Statistical Analysis
3. Results
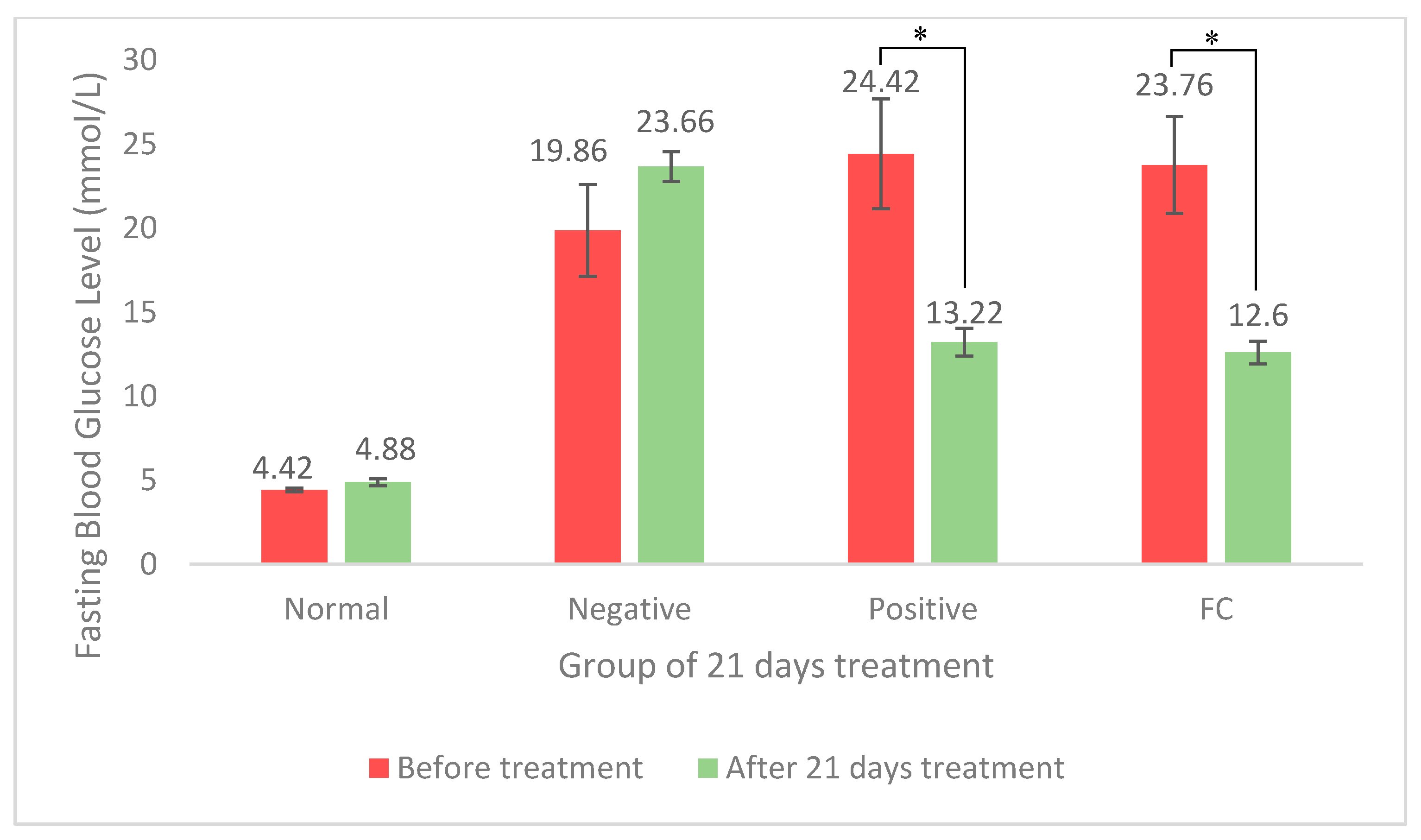
3.1. Antihyperglycemic Activity of FC Extract
3.2. Sperm Quality Analysis
3.3. Sperm Proteomics Analysis
3.3.1. Concentration of Sperm Protein
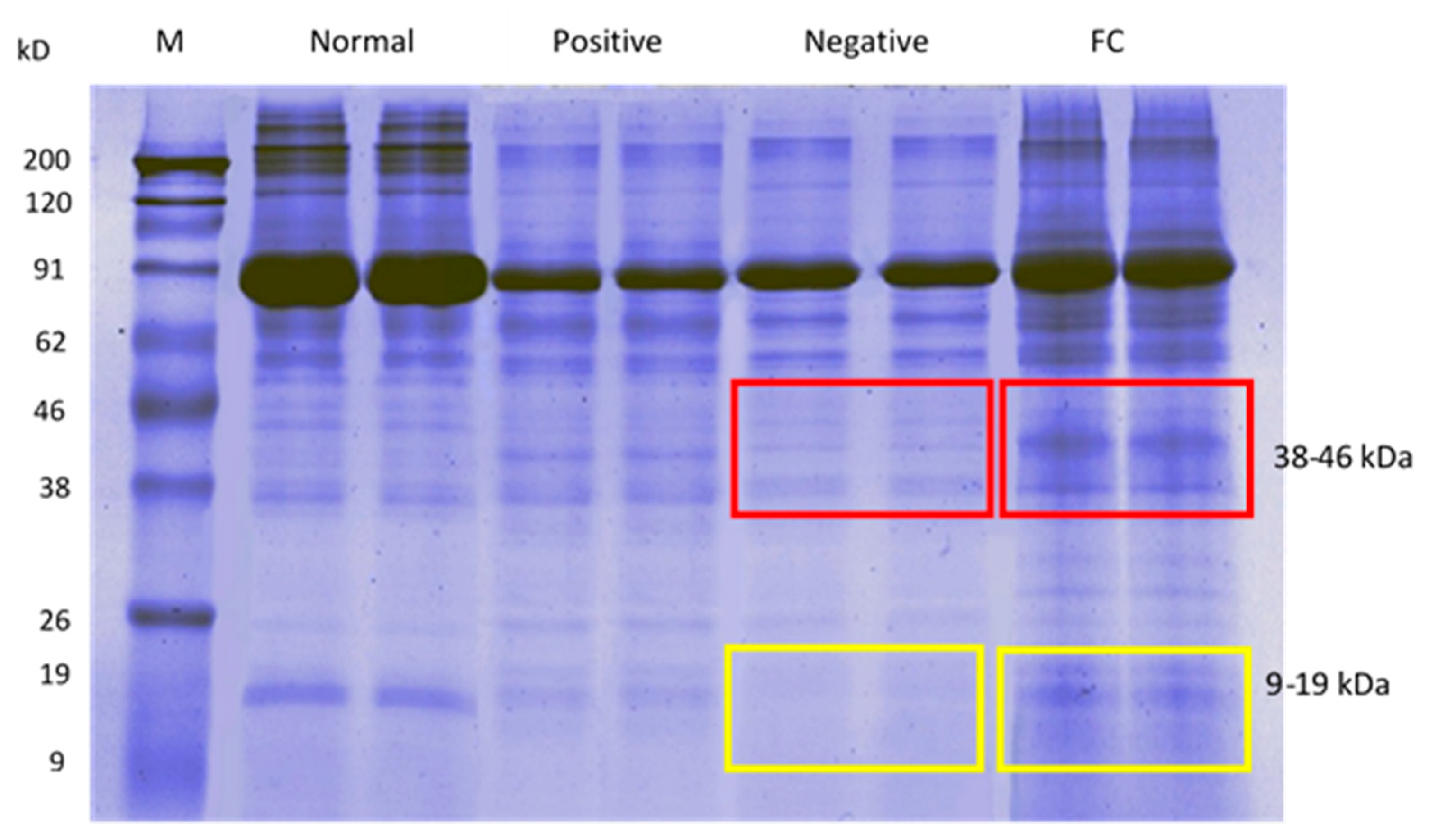
3.3.2. Sperm Protein Profile via SDS-PAGE
3.3.3. LC-MS/MS Analysis
3.3.4. Gene Ontology Analysis via Blast2GO and WEGO
3.3.5. Sperm Unique Protein upon FC Treatment
4. Discussion
4.1. Antihyperglycemic Activity of FC Extract
4.2. Sperm Quality Analysis
4.3. Sperm Proteomics Analysis
4.3.1. Sperm Protein Concentration, Protein Expression via SDS-PAGE, LC-MS/MS, and Gene Ontology Analysis
4.3.2. Sperm Unique Protein upon FC Treatment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Report on Diabetes; WHO: Geneva, Switzerland, 2016. [Google Scholar]
- El-Shobaki, F.A.; El-Bahay, A.M.A.; Esmail, R.S.A.; El-Megeid, A.A.A.; Esmail, D.S. Effect of figs fruit (Ficus carica L.) and its leaves on hyperglycemia in alloxan diabetic rats. World J. Dairy Food Sci. 2010, 5, 45–57. [Google Scholar]
- Singh, A.K.; Tomarz, S.; Chaudhari, A.R.; Sinqh, R.; Verma, N. Type 2 diabetes mellitus affects male fertility potential. Indian J. Physiol. Pharmacol. 2014, 58, 403–406. [Google Scholar] [PubMed]
- Pavlinkova, G.; Margaryan, H.; Zatecka, E.; Valaskova, E.; Elzeinova, F.; Kubatova, A.; Bohuslavova, R.; Peknicova, J. Transgenerational inheritance of susceptibility to diabetes-induced male subfertility. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Pitetti, J.; Calvel, P.; Zimmermann, C.; Conne, B.; Papaioannou, M.D.; Aubry, F.; Cederroth, C.R.; Urner, F.; Fumel, B.; Crausaz, M.; et al. An essential role for insulin and IGF1 receptors in regulating sertoli cell proliferation, testis size and FSH action in mice. Mol. Endocrinol. 2013, 27, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.L.; Liu, Y.; Liu, M.E.; Pan, J.X.; Guo, M.X.; Sheng, J.Z.; Huang, H.F. The effects of diabetes on male fertility and epigenetic regulation during spermatogenesis. Asian J. Androl. 2015, 17, 948–953. [Google Scholar] [PubMed]
- Pennington, S.R.; Wilkins, M.R.; Hochstrasser, D.F.; Dun, M.J. Proteome analysis: From protein characterization to biological function. Trends Cell Biol. 1997, 7, 168–173. [Google Scholar] [CrossRef]
- Rivas, J.D.L.; Luis, A.D. Interactome data and databases: Different types of protein interaction. Comp. Funct. Genom. 2004, 5, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Amaral, A.; Castillo, J.; Ramalho-Santos, J.; Oliva, R. The combined human sperm proteome: Cellular pathways and implications for basic and clinical science. Hum. Reprod. Update 2014, 20, 40–62. [Google Scholar] [CrossRef] [Green Version]
- An, T.; Wang, Y.; Liu, J.; Pan, Y.; Liu, Y.; He, Z.; Mo, F.; Li, H.; Gu, Y.; Lv, B.; et al. Comparative analysis of proteomes between diabetic and normal human sperm: Insights into the effects of diabetes on male reproduction based on regulation of mitochondria-related proteins. Mol. Reprod. Dev. 2017, 85, 7–16. [Google Scholar] [CrossRef]
- Bertoldo, M.J.; Faure, M.; Dupont, J.; Froment, P. Impact of metformin on reproductive tissues: an overview from gametogenesis to gestation. Ann. Transl. Med. 2014, 2, 1–13. [Google Scholar]
- Owen, M.R.; Doran, E.; Halestrap, A.P. Evidence that metformin exerts its anti-diabetic effects through inhibition complex I of the mitochondrial respiratory chain. Biochem. J. 2000, 348, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Hurtado de Llara, A.; Martin-Hidalgo, D.; Garcia-Marin, L.J.; Bragado, M.J. Metformin blocks mitochondrial membrane potential and inhibits sperm motility in fresh and refrigerated boar spermatozoa. Reprod. Dom. Anim. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Khaidatul Akmar, K.; Mahanem, M.N. Gynura procumbens leaf improves blood glucose level, restores fertility and libido of diabetic-induced male rats. Sains Malays. 2017, 46, 1471–1477. [Google Scholar] [CrossRef]
- Naglaa, Z.H.E.; Hesham, A.M.; Fadil, H.A. Impact of metformin on immunity and male fertility in rabbits with alloxan-induced diabetes. J. Am. Sci. 2010, 6, 417–426. [Google Scholar]
- Irudayaraj, S.S.; Sunil, C.; Stalin, A.; Duraipandiyan, V.; Al-Dhabi, N.A.; Ignacimuthu, S. Protective effects of Ficus carica leaves on glucose and lipids levels, carbohydrate metabolism enzymes and β-cells in type 2 diabetic rats. Pharm. Biol. 2017, 55, 1074–1081. [Google Scholar] [CrossRef] [Green Version]
- Naghdi, M.; Maghbool, M.; Seifalah-Zade, M.; Mahaldashtian, M.; Makoolati, Z.; Kouhpayeh, S.A.; Ghasemi, A.; Ferezdouni, N. Effects of common fig (Ficus carica) leaf extracts on sperm parameters and testis of mice intoxicated with formaldehyde. Evid.-Based Comp. Alt. 2016, 2016, 2539127. [Google Scholar] [CrossRef] [Green Version]
- Perez, C.; Dominguez, E.; Canal, J.R.; Campillo, J.E.; Torres, M.D. Hypoglycaemic activity of aqueous extract from Ficus carica (Fig tree) leaves in streptozotocin diabetic rats. Pharm. Biol. 2000, 38, 181–186. [Google Scholar] [CrossRef]
- Lin, M.; Zhang, X.; Murdoch, R.N.; Aitken, R.J. Swim-up of tammar wallaby (Macropus eugenii) spermatozoa in Biggers, Whitter and Whittingham (BWW) medium: maximisation of sperm motility, minimization of impairment of sperm metabolism and induction of sperm hyperactivation. Reprod. Fert. Develop. 2015, 29, 345–356. [Google Scholar] [CrossRef]
- Seed, J.; Chapin, R.E.; Clegg, E.D.; Dostal, L.A.; Foote, R.H.; Hurtt, M.E.; Klinefelter, G.R.; Makris, S.L.; Perreault, S.D.; Schrader, S.; et al. Methods for assessing sperm motility, morphology and counts in the rat, rabbit and dog: A consensus report. Reprod. Toxicol. 1996, 10, 237–244. [Google Scholar] [CrossRef]
- WHO. Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; Cambridge University Press: New York, NY, USA, 2010. [Google Scholar]
- Yunianto, I.; Nurul Ain, K.B.; Mahanem, M.N. Antifertility properties of Centella asiatica ethanolic extract as a contraceptive agent: Preliminary study of sperm proteomic. Asian Pac. J. Reprod. 2018, 6, 212–216. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- King, A.J.F. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef] [Green Version]
- Rakieten, N.; Rakieten, M.L.; Nadkarni, M.V. Studies on the diabetogenic action of streptozotocin. Cancer Chemother. Rep. 1963, 29, 91. [Google Scholar] [PubMed]
- Jitrapakdee, S. Regulation of insulin secretion: role of mitochondrial signaling. Diabetologia 2010, 53, 1019–1032. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, G. Insulin and insulin resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar]
- Duca, F.A.; Cote, C.D.; Rasmussen, B.A.; Zadeh-Tahmasebi, M.; Rutter, G.A.; Filippi, B.M.; Lam, T.K.T. Metformin activates a duodenal Ampk-dependent pathway to lower hepatic glucose production in rats. Nat. Med. 2015, 21, 506–511. [Google Scholar] [CrossRef]
- Zang, Y.; Zhang, L.; Igarashi, K.; Yu, C. The anti-obesity and anti-diabetic effects of kaempferol glycosides from unripe soybean leaves in high-fat-diet mice. Food Funct. 2015, 6, 834–841. [Google Scholar] [CrossRef]
- Jensen, J.; Rustad, P.I.; Kolnes, A.J.; Lai, Y.C. The role of skeletal muscle glycogen breakdown for regulation of insulin sensitivity by exercise. Front. Physiol. 2011, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mopuri, R.; Ganjayi, M.; Meriga, B.; Koorbanally, N.A.; Islam, M.S. The effects of Ficus carica on the activity of enzymes related to metabolic syndrome. J. Food Drug Anal. 2018, 26, 201–210. [Google Scholar] [CrossRef]
- Amira, K.; Malina, J.; Mahanem, M.N. Ameliorative effect of Moringa oleifera fruit extract on reproductive parameters in diabetic-induced male rats. Pharmacogn. J. 2018, 10, S54–S58. [Google Scholar]
- Noor, A.; Gunasekaran, S.; Vijayalakshmi, M.A. Improvement of insulin secretion and pancreatic β-cell function in streptozotocin-induced diabetic rats treated with Aloe vera extract. Pharmacogn. Res. 2017, 9, S99–S104. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Li, S.; Liu, Y.; Huang, M.; Ho, C. Anti-diabetic activity of chemically profiled green tea and black tea extracts in a type 2 diabetes mice model via different mechanisms. J. Funct. Foods 2013, 5, 1784–1793. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- Abdel-Nasser, B.S.; Nahla, A.A.; Eman, N.A.; Nada, M.M. Antioxidant and hepatoprotective activities of Egyptian moraceous plants against carbon tetrachloride-induced oxidative stress and liver damage in rats. Pharm. Biol. 2010, 48, 1255–1264. [Google Scholar]
- Boukhalfa, F.; Kadri, N.; Bouchemel, S.; Cheikh, S.A.; Chebout, I.; Madani, K.; Chibane, M. Antioxidant activity and hypolipidemic effect of Ficus carica leaf and twig extracts in Triton-WR-1339-induced hyperlipidemic mice. Med. J. Nutr. Metab. 2018, 11, 37–50. [Google Scholar] [CrossRef]
- Takahashi, T.; Okiura, A.; Saito, K.; Kohno, M. Identification of phenylpropanoids in fig (Ficus carica L.) leaves. J. Agric. Food Chem. 2014, 62, 10076–10083. [Google Scholar] [CrossRef]
- Irudayaraj, S.S.; Stalin, A.; Sunil, C.; Duraipandiyan, V.; Al-Dhabi, N.A.; Ignacimuthu, S. Antioxidant, antilipidemic and antidiabetic effects of ficusin with their effects on GLUT4 translocation and PPARγ expression in type 2 diabetic rats. Chem. Biol. Interact. 2016, 2016, 1–38. [Google Scholar] [CrossRef]
- Kannappan, S.; Anuradha, C.V. Insulin sensitizing actions of fenugreek seed polyphenols, quercetin and metformin in a rat model. Indian J. Med. Res. 2009, 129, 401–408. [Google Scholar]
- Fang, X.K.; Gao, J.; Zhu, D.N. Kaempferol and quercetin isolated from Euonymus alatus improve glucose uptake of 3T3-L1 cells without adipogenesis activity. Life Sci. 2008, 82, 615–622. [Google Scholar] [CrossRef]
- Ibitoye, O.B.; Uwazie, J.N.; Ajiboye, T.O. Bioactivity-guided isolation of kaempferol as the antidiabetic principle from Cucumis sativus L. fruits. J. Food Biochem. 2017, 2017, e12479. [Google Scholar] [CrossRef]
- Narasimhan, A.; Chinnaiyan, M.; Karundevi, B. Ferulic acid exerts its antidiabetic effect by modulating insulin signaling molecules in the liver of high fat diet and fructose-induced type-2 diabetic adult male rat. Appl. Physiol. Nutr. Metab. 2015, 40, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Talarczyk-Desole, J.; Berger, A.; Taszarek-Hauke, G.; Hauke, J.; Pawelczyk, L.; Jedrzejczak, P. Manual vs. computer-assisted sperm analysis: can CASA replace manual assessment of human semen in clinical practice? Ginekol. Pol. 2017, 88, 56–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eravuchira, P.J.; Mirsky, S.K.; Barnea, I.; Balberg, M.; Shaked, N.T. Individual sperm selection by microfluidics integrated with interferometric phase microscopy. Methods 2018, 136, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.M. Tales from the tail: What do we really know review about sperm motility? J. Androl. 2003, 24, 790–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasan, S.S. Semen analysis and sperm function tests: How much to test? Indian J. Urol. 2011, 27, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Li, B.; Fan, L.Q.; Zhu, W.B.; Chen, X.J.; Feng, J.H.; Yang, C.L.; Zhang, Y.H. Does sperm morphology affect the outcome of intrauterine insemination in patients with normal sperm concentration and motility? Andrologia 2012, 44, 299–304. [Google Scholar] [CrossRef]
- Kim, S.T.; Moley, K.H. Paternal effect on embryo quality in diabetic mice is related to poor sperm quality and associated with decreased glucose transporter expression. Reproduction 2008, 136, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.G.; Martins, A.D.; Cavaco, J.E.; Socorro, S.; Oliveira, P.F. Diabetes, insulin-mediated glucose metabolism and Sertoli/blood-testis barrier function. Tissue Barriers 2013, 1, e23992. [Google Scholar] [CrossRef] [Green Version]
- Mita, M.; Hall, P.F. Metabolism of round spermatids from rats: Lactate as the preferred substrate. Biol. Reprod. 1982, 26, 445–455. [Google Scholar] [CrossRef]
- Mahanem, M.N.; Nani, R.M.R. Anti-hyperglycemic effect of Gynura procumbens methanolic extract on fertility and libido of induced diabetic male rats. Sains Malays. 2012, 41, 1549–1556. [Google Scholar]
- Jayakumar, S.P.; Sen, M.; Jagadeesan, M.; Sundararajan, R. Antidiabetic and antioxidant effects of ethanolic extract of Anjir leaves (Ficus carica) in alloxan induced diabetic rat. Asian J. Phytomed. Clin. Res. 2014, 2, 1–10. [Google Scholar]
- Shimaa, A.H.; Tamer, S.I.; Omar, A.A. Combination of Ficus carica leaves extract and ubiquinone in a chronic model of lithium induce reproductive toxicity in rats: Hindrance of oxidative stress and apoptotic marker of sperm cell degradation. IOSR J. Pharm. Biol. Sci. 2017, 12, 64–73. [Google Scholar]
- Shahinuzzaman, M.; Yaakob, Z.; Abdullah Sani, N.; Akhtar, P.; Zahidul Islam, M.D.; Afsana Mimi, M.S.T.; Shamsudin, S.A. Optimization of extraction parameters for antioxidant and total phenolic content of Ficus carica L. latex from White Genoa cultivar. Asian J. Chem. 2019, 31, 1859–1865. [Google Scholar] [CrossRef]
- Ambiye, V.R.; Langade, D.; Dongre, S.; Aptikar, P.; Kulkarni, M.; Dongre, A. Clinical evaluation of the spermatogenic activity of the root extract of aswagandha (Withania somnifera) in oligospermic males: A pilot study. Evid.-Based Compl. Altern. Med. 2013, 2013, 571420. [Google Scholar]
- Ying, X.; Liu, Y.; Guo, Q.; Qu, F.; Guo, W.; Zhu, Y.; Ding, Z. Endoplasmic reticulum protein 29 (ERp29), a protein related to sperm maturation is involved in sperm-oocyte fusion in mouse. Reprod. Biol. Endocrinol. 2010, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Li, J.; Chen, Y.; Wang, J.J.; Ratan, R.; Zhang, S.X. Activation of endoplasmic reticulum stress by hyperglycemia is essential for Muller cell-derived inflammatory cytokine production in diabetes. Diabetes 2012, 61, 492–504. [Google Scholar] [CrossRef] [Green Version]
- Back, S.H.; Kaufman, R.J. Endoplasmic reticulum stress and type 2 diabetes. Annu. Rev. Biochem. 2012, 81, 767–793. [Google Scholar] [CrossRef] [Green Version]
- Kriegel, T.M.; Heldenreich, F.; Kettner, K.; Pursche, T.; Hoflack, B.; Grunewald, S.; Poenicke, K.; Glander, H.; Paasch, U. Identification of diabetes and obesity associated proteomic changes in human spermatozoa by difference gel electrophoresis. Reprod. BioMed Online 2009, 19, 660–670. [Google Scholar] [CrossRef] [Green Version]
- Khaidatul Akmar, K.; Mohd Aizat, W.; Mahanem, M.N. Gynura procumbens improved fertility of diabetic rats: Preliminary study of sperm proteomic. Evid.-Based Compl. Altern. Med. 2018. [Google Scholar] [CrossRef] [Green Version]
- Dimmer, E.C.; Huntley, R.P.; Barrell, D.G.; Binns, D.; Draghici, S.; Camon, E.B.; Hubank, M.; Talmud, P.J.; Apweiler, R.; Lovering, R.C. The gene ontology—Providing a functional role in proteomic studies. Pract. Proteom. 2008, 1, 2–11. [Google Scholar] [CrossRef]
- Ghosh, A.; Jana, K.; Ali, K.M.; De, D.; Chatterjee, K.; Ghosh, D. Corrective role of Eugenia jambolana on testicular impairment in streptozotocin-induced diabetic male albino rat: An approach through genomic and proteomic study. Andrologia 2013, 46, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Bruey, J.; Bruey-Sedano, N.; Luciano, F.; Zhai, D.; Balpai, R.; Xu, C.; Kress, C.L.; Bailly-Maitre, B.; Li, X.; Osteman, A.; et al. Bcl-2 and Bcl-XL regulate proinflammatory caspase-1 activation by interaction with NALP1. Cell 2007, 129, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pena-Ramos, E.A.; Xiong, Y.L. Antioxidative activity of soy protein hydrolysates in a liposomal system. J. Food Sci. 2002, 67, 2952–2956. [Google Scholar] [CrossRef]
- Pierro, G.D.; O’Keeffe, M.B.; Poyarkov, A.; Lomolino, G.; FitzGerald, R.J. Antioxidant activity of bovine casein hydrolysates produced by Ficus carica L.—derived proteinase. Food Chem. 2014, 156, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, H.; Shiau, C. Free amino acids and peptide as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Guo, W.; Adachi, T.; Matsui, R.; Xu, S.; Jiang, B.; Zou, M.; Kirber, M.; Lieberthal, W.; Cohen, R.A. Quantitative assessment tyrosine nitration of manganese superoxide dismutase in angiotensin II-infused rat kidney. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H1396–H1403. [Google Scholar] [CrossRef] [Green Version]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B. Diabetes, oxidative stress an antioxidants: A review. J. Biochem. Mol. Toxicol. 2002, 17, 24–38. [Google Scholar] [CrossRef]
- Li, Y.; Huang, T.; Carlson, E.J.; Melov, S.; Ursell, P.C.; Olson, J.L.; Noble, L.J.; Yoshimura, M.P.; Berger, C.; Chan, P.H.; et al. Dilated cardiomyopathy and neeonatal lethality in mutant mice lacking manganese superoxide dismutase. Nat. Genet. 1995, 11, 376–381. [Google Scholar] [CrossRef]
- Nishikawa, T.; Edelstein, D.; Du, X.L.; Yamagishi, S.; Matsumura, T.; Kaneda, Y.; Yorek, M.A.; Beebe, D.; Oates, P.J.; Hammes, H.; et al. Normalizing mitochondrial superoxide production blocks three pathways of hyperglycemic damage. Nature 2000, 404, 787–790. [Google Scholar] [CrossRef]
- Kowluru, R.A.; Kowluru, V.; Xiong, Y.; Ho, Y. Overexpression of mitochondrial superoxide dismutase in mice protects the retina from diabetes-induced oxidative stress. Free Radic. Biol. Med. 2006, 41, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Zheng, S.; Metreveli, N.S.; Epstein, P.N. Protection of cardiac mitochondria by overexpression of MnSOD reduces diabetic cardiomyopathy. Diabetes 2006, 55, 798–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cocchia, N.; Pasolini, M.P.; Mancini, R.; Petrazzuolo, O.; Cristofaro, I.; Rosapane, I.; Sica, A.; Tortora, G.; Lorizio, R.; Paraggio, G.; et al. Effect of SOD (superoxide dismutase) protein supplementation in semen extenders on motility, viability, acrosome status and ERK (extracellular signal-regulated kinase) protein phosphorylation of chilled stallion spermatozoa. Theriogenology 2011, 75, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.G.; Winterbourn, C.; Hampton, M.B. Mitochondrial peroxiredoxin involvement in antioxidant defence and redox signaling. Biochem. J. 2010, 425, 313–325. [Google Scholar] [CrossRef] [Green Version]
- Knoops, B.; Goemaere, J.; Eecken, V.V.D.; Declercq, J. Peroxiredoxin 5: Structure, mechanism and function of the mammalian atypical 2-Cys Peroxiredoxin. Antioxid. Redox Sign. 2011, 15, 817–829. [Google Scholar] [CrossRef]
- Kubo, E.; Singh, D.P.; Fatma, N.; Akagi, Y. TAT-mediated peroxiredoxin 5 and 6 protein transduction protects against high glucose-induced cytotoxicity in retinal pericytes. Life Sci. 2009, 84, 857–864. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, C. Peroxiredoxins: Hidden players in the antioxidant defence of human spermatozoa. Basic Clin. Androl. 2014, 24, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chow, C.Y.; Avila, F.W.; Clark, A.G.; Wolfner, M.F. Induction of excessive endoplasmic reticulum stress in the drosophila male accessory gland results in infertility. PLoS ONE 2015, 10, e0119386. [Google Scholar] [CrossRef] [Green Version]
- Guzel, E.; Arlier, S.; Guzeloglu-Kayisli, O.; Tabak, M.S.; Ekiz, T.; Semerci, N.; Larsen, K.; Schatz, F.; Lockwood, C.J.; Kayisli, U.A. Endoplasmic reticulum stress and homeostasis in reproductive physiology and pathology. Int. J. Mol. Sci. 2017, 18, 792. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Tang, W.K.; Zhang, T.; Xia, D. A mighty “Protein Extractor” of the cell: Structure and function of the p97/CDC48 ATPase. Front. Mol. Biosci. 2017, 4, 1–20. [Google Scholar] [CrossRef]
- Sitia, R.; Braakman, I. Quality control in the endoplasmic reticulum protein factory. Nature 2003, 426, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, C.; Rowicka, M.; Kudlicki, A.; Nowis, D.; McConnell, E.; Kujawa, M.; DeMartino, G.N. Endoplasmic reticulum stress and of the degradation of N-end rule and ubiquitin-fusion degradation pathway substrates in mammalian cells. Mol. Biol. Cell. 2006, 17, 4606–4618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, R.; Zheng, D.; Xiong, S.; Hill, D.J.; Sun, T.; Gardiner, R.B.; Fan, G.; Lu, Y.; Abel, E.D.; Greer, P.A.; et al. Mitochondrial calpain-1 disrupts ATP synthase and induces superoxide generation in type 1 diabetic hearts: A novel mechanism contributing to diabetic cardiomyopathy. Diabetes 2016, 65, 255–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, W.; Keenan, H.A.; Li, Q.; Ishikado, A.; Kannt, A.; Sadowski, T.; Yorek, M.A.; Wu, I.; Lockhart, S.; Coppey, L.J.; et al. Pyruvate kinase M2 activation may protect against the progression of diabetic glomerular pathology and mitochondrial dysfunction. Nat. Med. 2017, 23, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, Y.; Camporez, J.G.; Petersen, M.C.; Pesta, D.; Perry, R.J.; Jurczak, M.J.; Cline, G.W.; Shulman, G.I. Genetic activation of pyruvate dehydrogenase alters oxidative substrate selection to induce skeletal muscle insulin resistance. Proc. Natl. Acad. Sci. USA 2014, 111, 16508–16513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.; Yeh, T.; Chen, S.; Tsai, Y.; Chou, C.; Yang, Y.; Huang, H. Nonmuscle Myosin IIA (Myosin Heavy Polypeptide 9): A novel class of signal transducer mediating the activation of Gαh/Phospholipase C-δ1 pathway. Endocrinology 2010, 151, 876–885. [Google Scholar] [CrossRef] [Green Version]
- Adams, I.R.; McLaren, A. Sexually dimorphic development of mouse primordial germ cells: switching from oogenesis to spermatogenesis. Development 2002, 129, 1155–1164. [Google Scholar]
- Moniot, B.; Ujjan, S.; Champagne, J.; Hirai, H.; Aritake, K.; Nagata, K.; Dubois, E.; Nidelet, S.; Nakamura, M.; Urade, Y.; et al. Prostaglandin D2 acts through the Dp2 receptor to influence male germ cell differentiation in the foetal mouse testis. Development 2014, 141, 3561–3571. [Google Scholar] [CrossRef] [Green Version]
- Moniot, B.; Declosmenil, F.; Barrionuevo, F.; Scherer, G.; Aritake, K.; Malki, A.; Marzi, L.; Cohen-Solal, A.; Georg, I.; Klattig, J.; et al. The PGD2 pathway, independently of FGF9, amplifies SOX9 activity in Sertoli cells during male sexual differentiation. Development 2009, 136, 1813–1821. [Google Scholar] [CrossRef] [Green Version]
- Samy, E.T.; Li, J.C.H.; Grima, J.; Lee, W.M.; Silvestrini, B.; Cheng, C.Y. Sertoli cell Prostaglandin D2 Synthetase is a mulitifunctional molecule: Its expression and regulation. Endocrinology 2000, 141, 710–721. [Google Scholar] [CrossRef]
- Gelato, K.A.; Fischle, W. Role of histone modifications in defining chromatin structure and function. J. Biol. Chem. 2008, 389, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Xie, Y.M.; Li, X.; Luo, J.; Shi, X.Q.; Hong, X.; Pan, Y.H.; Ma, X. Mass spectrometry analysis of dynamic post-translational modifications of TH2B during spermatogenesis. Mol. Hum. Reprod. 2009, 15, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, S.M. The dynein microtubule motor. BBA 2000, 1496, 60–75. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhao, M.; Meistrich, M.L.; Kumar, R. Stage-specific expression of Dynein Light Chain-1 and its interacting Kinase, p21-activated Kinase-1, in rodent testes: Implications in spermatogenesis. J. Histochem. Cytochem. 2005, 53, 1235–1243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, Y.; Wei, B.; Lai, Y.; Yan, Q.; Gui, Y.; Cai, Z. Functional expression of Ropporin in human testis and ejaculated spermatozoa. J. Androl. 2011, 32, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, S.E.; Sisson, J.H.; Wyatt, T.A.; Pavlik, J.A.; Gambling, T.M.; Carson, J.L.; Carr, D.W. Loss of ASP but not ROPN1 reduces mammalian ciliary motility. Cytoskeleton 2012, 69, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubota, H.; Hynes, G.; Willison, K. The chaperonin containing t-complex polypeptide 1 (TCP-1): Multisubunit machinery assisting in protein folding and assembly in the eukaryotic cytosol. Eur. J. Biochem. 1995, 230, 3–16. [Google Scholar] [CrossRef]
- Redgrove, K.A.; Anderson, A.L.; Dun, M.D.; McLaughlin, E.A.; O’Bryan, M.K.; Aitken, R.J.; Nixon, B. Involvement of multimeric protein complexes in mediating the capacitation-dependent binding of human spermatozoa to homologous zonae pellucidae. Dev. Biol. 2011, 356, 460–474. [Google Scholar] [CrossRef] [Green Version]
- Dun, M.D.; Smith, N.D.; Baker, M.A.; Lin, M.; Aitken, R.J.; Nixon, B. The chaperoning containing TCP1 complex (CCT/TRiC) is involved in mediating sperm-oocyte interaction. J. Biol. Chem. 2011, 286, 36875–36887. [Google Scholar] [CrossRef] [Green Version]
- Panneerdoss, S.; Siva, A.B.; Kameshwari, D.B.; Rangaraj, N.; Shivaji, S. Association of lactate, intracellular pH and intracellular calcium during capacitation and acrosome reaction: Contribution of hamster sperm Dihydrolipoamide dehydrogenase, the E3 subunit of Pyruvate dehydrogenase complex. J. Androl. 2012, 33, 699–710. [Google Scholar] [CrossRef]
- Kumar, V.; Kota, V.; Shivaji, S. Hamster sperm capacitation: Role of Pyruvate dehydrogenase A and Dihydrolipoamide dehydrogenase. Biol. Reprod. 2008, 79, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, M.T.; Yang, H.; Magnuson, T.; Patel, M.S. Targeted disruption of the murine dihydrolipoamide dehydrogenase gene (Dld) results in perigastrulation lethality. Proc. Natl. Acad. Sci. USA 1997, 94, 14512–14517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitra, K.; Shivaji, S. Novel tyrosine-phosphorylated post-pyruvate metabolic enzyme, Dihydrolipoamide dehydrogenase, involved in capacitation of hamster spermatozoa. Biol. Reprod. 2004, 70, 887–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, N.; Ikawa, M.; Isotani, A.; Okabe, M. The immunoglobulin superfamily protein Izumo is required for sperm to fuse with eggs. Nature 2005, 434, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, E.; Doe, B.; Goulding, D.; Wright, G.J. Juno is the egg Izumo receptor and is essential for mammalian fertilization. Nature 2014, 508, 483–487. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
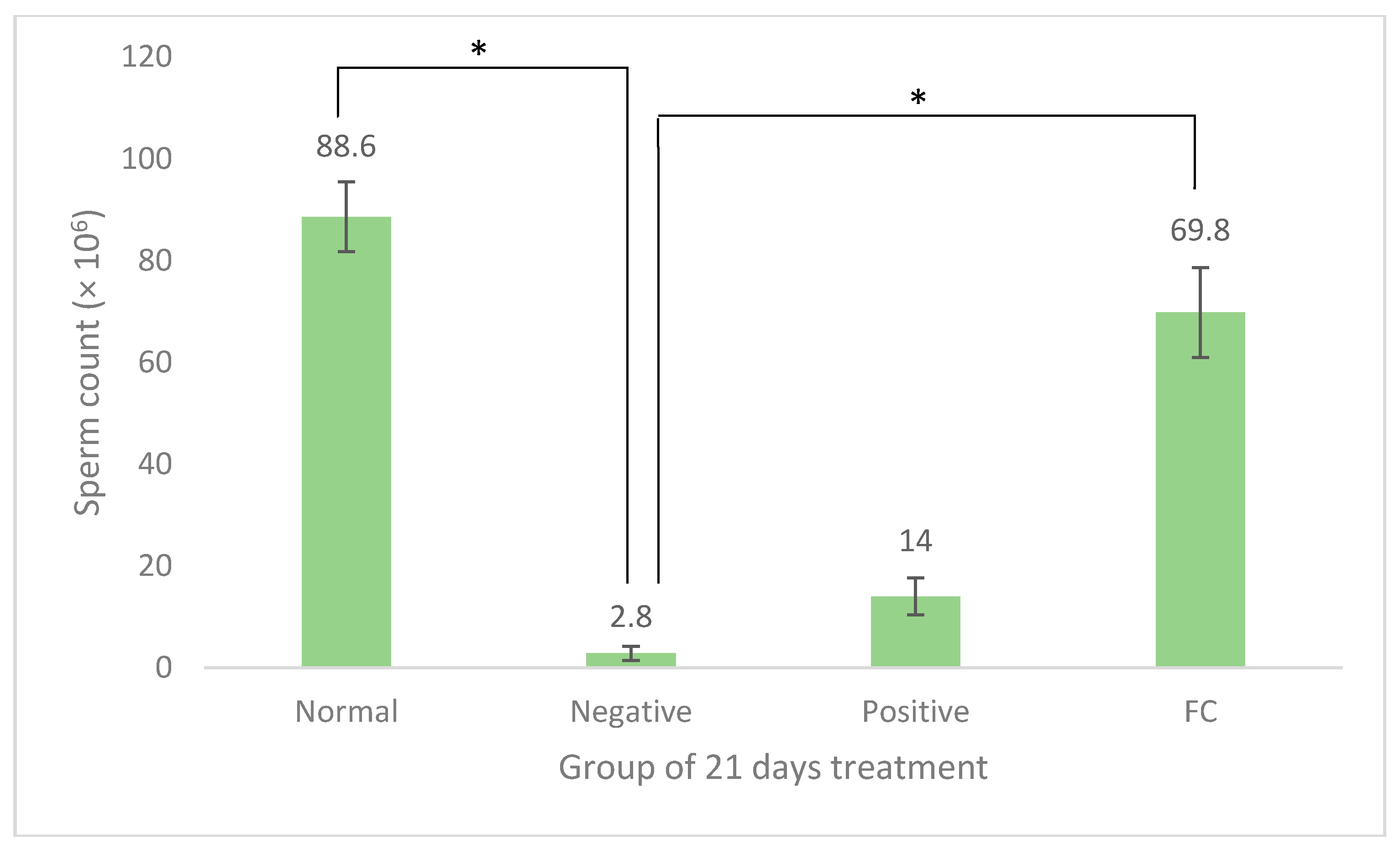{kind=link}
{kind=link}
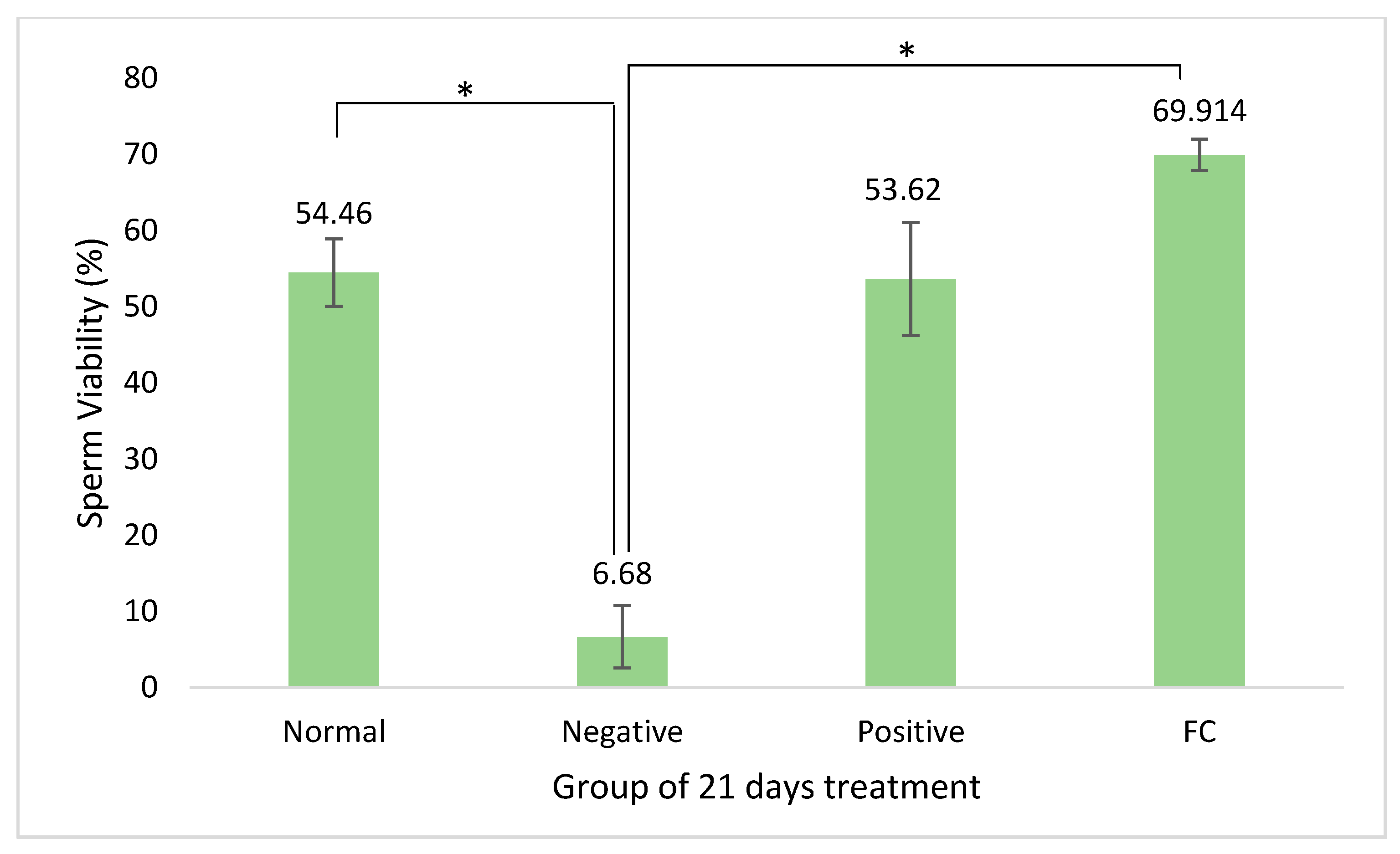{kind=link}
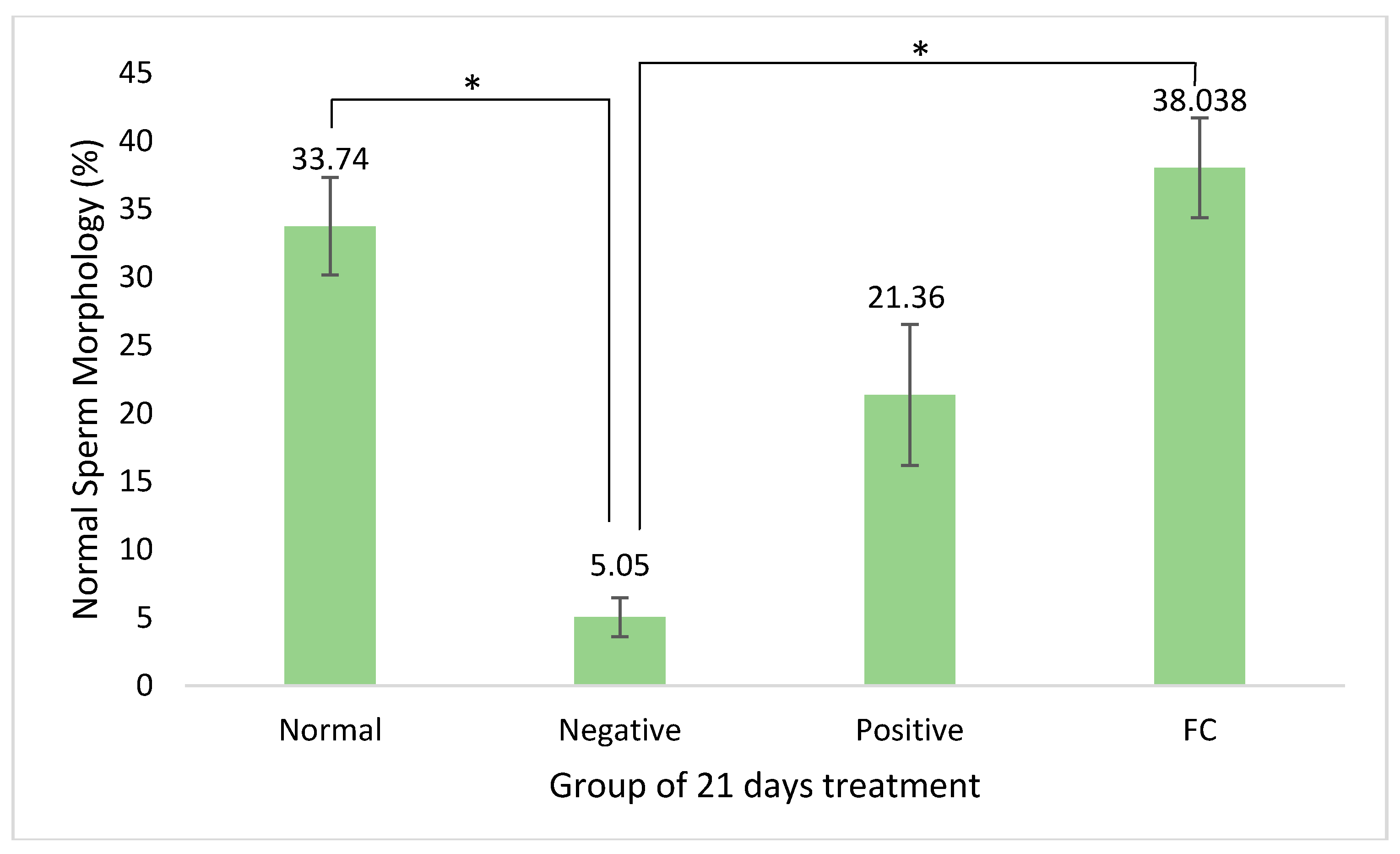{kind=link}
{kind=link}
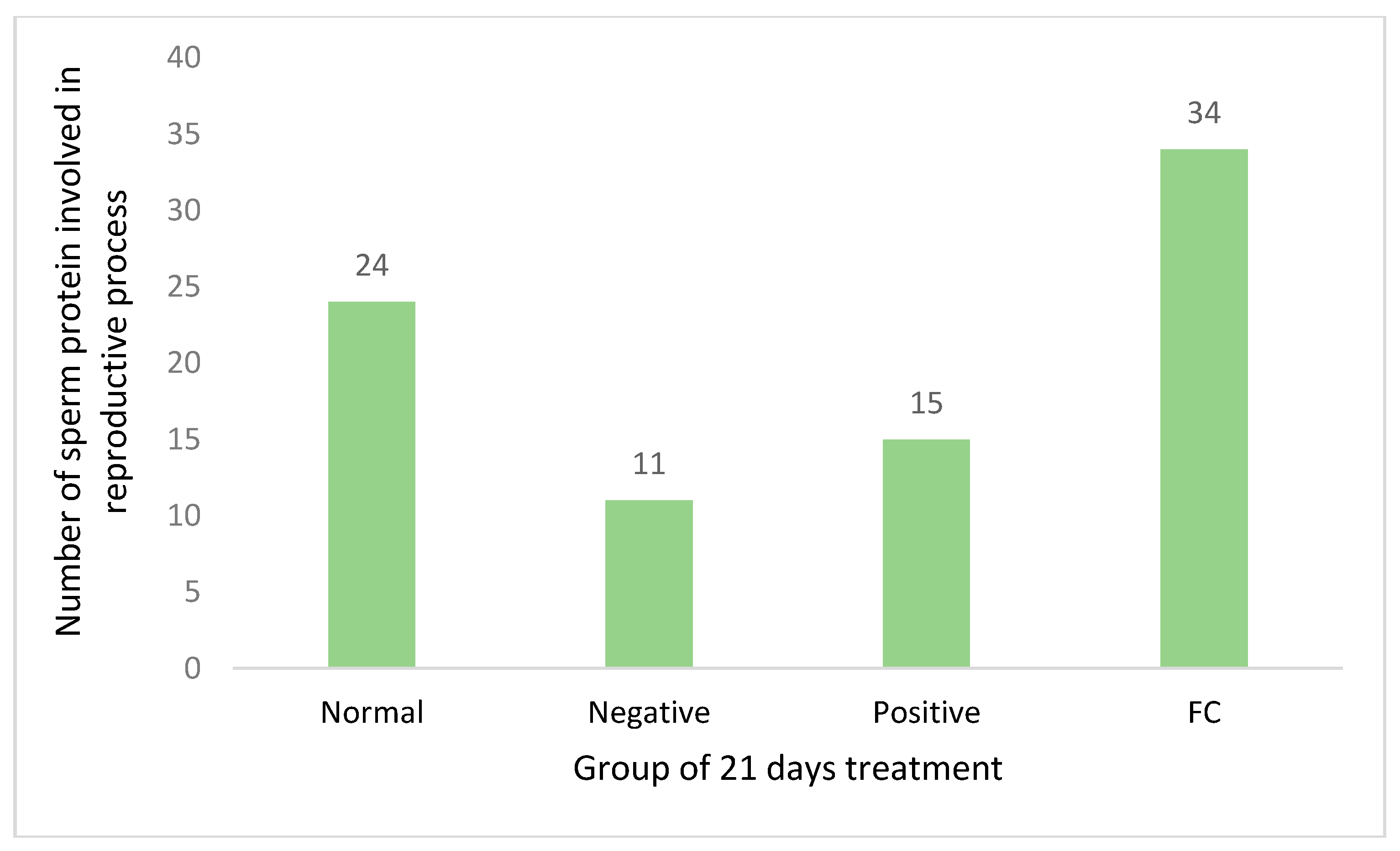{kind=link}
{kind=link}
| Group | Concentration of Protein (mg/mL) |
|---|---|
| Normal | 1.13 |
| Negative (Diabetic without treatment) | 0.8 |
| Positive (Diabetic treated with metformin) | 1.05 |
| FC (Diabetic treated with F. carica extract) | 1.15 |
| Group | Total Number of Protein |
|---|---|
| Normal | 149 |
| Negative (Diabetic without treatment) | 55 |
| Positive (Diabetic treated with metformin) | 90 |
| FC (Diabetic treated with F. carica extract) | 155 |
| Protein ID (Uniprot) | Name | Location | Molecular Weight (kDa) |
|---|---|---|---|
| 1. Protective unique protein | |||
| P07895 | Manganese superoxide dismutase | mitochondria | 24.66 |
| Q9R063 | Peroxiredoxin-5, mitochondrial | mitochondria | 22.17 |
| P46462 | Transitional endoplasmic reticulum ATPase | nucleus | 89.29 |
| P11980 | Pyruvate kinase PKM | cytoplasm | 57.78 |
| Q06647 | ATP synthase subunit O | mitochondria | 23.38 |
| Q06437 | Pyruvate dehydrogenase E1 component subunit alpha | mitochondria | 43.37 |
| 2. Reproductive unique protein | |||
| i. Spermatogenesis | |||
| Q62812 | Myosin-9 | cytoplasm | 226.2 |
| P22057 | Prostaglandin-H2 D-isomerase | nucleus | 21.29 |
| Q00729 | Histone H2B type 1-A | nucleosome | 14.22 |
| P63170 | Dynein light chain 1 | nucleus | 10.36 |
| ii. Fertilization | |||
| Q4KLL5 | Ropporin-1 | cytoplasm | 23.95 |
| Q5XIM9 | T-complex protein 1 subunit beta | cytoplasm | 57.42 |
| Q6P6R2 | Dihydrolipoyl dehydrogenase | mitochondria | 54.0 |
| Q6AY06 | Izumo sperm-egg fusion protein 1 | acrosome | 43.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abu Bakar, U.; Subramaniam, P.; Kamar Bashah, N.A.; Kamalrudin, A.; Kamaruzaman, K.A.; Jasamai, M.; Aizat, W.M.; Shahinuzzaman, M.; Mat Noor, M. Sperm Proteomics Analysis of Diabetic Induced Male Rats as Influenced by Ficus carica Leaf Extract. Processes 2020, 8, 395. https://doi.org/10.3390/pr8040395
Abu Bakar U, Subramaniam P, Kamar Bashah NA, Kamalrudin A, Kamaruzaman KA, Jasamai M, Aizat WM, Shahinuzzaman M, Mat Noor M. Sperm Proteomics Analysis of Diabetic Induced Male Rats as Influenced by Ficus carica Leaf Extract. Processes. 2020; 8(4):395. https://doi.org/10.3390/pr8040395
Chicago/Turabian StyleAbu Bakar, Umarqayum, Puvaratnesh Subramaniam, Nurul Ain Kamar Bashah, Amira Kamalrudin, Khaidatul Akmar Kamaruzaman, Malina Jasamai, Wan Mohd Aizat, M. Shahinuzzaman, and Mahanem Mat Noor. 2020. "Sperm Proteomics Analysis of Diabetic Induced Male Rats as Influenced by Ficus carica Leaf Extract" Processes 8, no. 4: 395. https://doi.org/10.3390/pr8040395
APA StyleAbu Bakar, U., Subramaniam, P., Kamar Bashah, N. A., Kamalrudin, A., Kamaruzaman, K. A., Jasamai, M., Aizat, W. M., Shahinuzzaman, M., & Mat Noor, M. (2020). Sperm Proteomics Analysis of Diabetic Induced Male Rats as Influenced by Ficus carica Leaf Extract. Processes, 8(4), 395. https://doi.org/10.3390/pr8040395