
Factors Affecting Detoxification of Softwood Enzymatic Hydrolysates Using Sodium Dithionite



Abstract
:1. Introduction
2. Materials and Methods
2.1. Pretreatment
2.2. Enzymatic Hydrolysis
2.3. Conditioning of Hydrolysate
2.4. Fermentation
2.5. Analysis of Ethanol and Glucose
2.6. Analysis of Monosaccharides
2.7. Determination of Microbial Inhibitors
2.8. Calculations
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ragauskas, A.J.; Williams, C.K.; Davison, B.H.; Britovsek, G.; Cairney, J.; Eckert, C.A.; Frederick, W.J., Jr.; Hallett, J.P.; Leak, D.J.; Liotta, C.L.; et al. The path forward for biofuels and biomaterials. Science 2006, 311, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Galbe, M.; Wallberg, O. Pretreatment for biorefineries: A review of common methods for efficient utilisation of lignocellulosic materials. Biotechnol. Biofuels 2019, 12, 294. [Google Scholar] [PubMed] [Green Version]
- Forest Statistics; Official Statistics of Sweden; Swedish University of Agricultural Sciences: Umeå, Sweden, 2018; Available online: https://www.slu.se/globalassets/ew/org/centrb/rt/dokument/skogsdata/skogsdata_2018_webb.pdf (accessed on 14 May 2021).
- Sun, S.; Sun, S.; Cao, X.; Sun, R. The role of pretreatment in improving the enzymatic hydrolysis of lignocellulosic materials. Bioresour. Technol. 2016, 199, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Öhgren, K.; Bura, R.; Lesnicki, G.; Saddler, J.; Zacchi, G. A comparison between simultaneous saccharification and fermenta-tion and separate hydrolysis and fermentation using steam-pretreated corn stover. Process Biochem. 2007, 42, 834–839. [Google Scholar] [CrossRef]
- Lynd, L.R.; Liang, X.; Biddy, M.J.; Allee, A.; Cai, H.; Foust, T.; Himmel, M.E.; Laser, M.S.; Wang, M.; Wyman, C.E. Cellulosic ethanol: Status and innovation. Curr. Opin. Biotechnol. 2017, 45, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Zhong, N.; Chandra, R.; Saddler, J.J.N. Sulfite post-treatment to simultaneously detoxify and improve the enzymatic hydrol-ysis and fermentation of a steam-pretreated softwood lodgepole pine whole slurry. ACS Sustain. Chem. Eng. 2019, 7, 5192−5199. [Google Scholar] [CrossRef]
- Hu, F.; Ragauskas, A.J. Pretreatment and lignocellulosic chemistry. Bioenergy Res. 2012, 5, 1043–1066. [Google Scholar] [CrossRef]
- Gandla, M.L.; Martín, C.; Jönsson, L.J. Analytical enzymatic saccharification of lignocellulosic biomass for conversion to bio-fuels and bio-based chemicals. Energies 2018, 11, 2936. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, L.J.; Martín, C. Pretreatment of lignocellulose: Formation of inhibitory by-products and strategies for minimizing their effects. Bioresour. Technol. 2016, 199, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Berlin, A.; Balakshin, M.; Gilkes, N.; Kadla, J.; Maximenko, V.; Kubo, S.; Saddler, J. Inhibition of cellulase, xylanase and β-glucosidase activities by softwood lignin preparations. J. Biotechnol. 2006, 125, 198–209. [Google Scholar] [CrossRef]
- Ximenes, E.; Kim, Y.; Mosier, N.; Dien, B.; Ladisch, M. Inhibition of cellulases by phenols. Enzym. Microb. Technol. 2010, 46, 170–176. [Google Scholar] [CrossRef]
- Jönsson, L.J.; Alriksson, B.; Nilvebrant, N.-O. Bioconversion of lignocellulose: Inhibitors and detoxification. Biotechnol. Biofuels 2013, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.K.; Um, Y.; Park, Y.-C.; Seo, J.-H.; Kim, K.H. Compounds inhibiting the bioconversion of hydrothermally pretreated lignocellulose. Appl. Microbiol. Biotechnol. 2015, 99, 4201–4212. [Google Scholar] [CrossRef]
- Cavka, A.; Stagge, S.; Jönsson, L.J. Identification of small aliphatic aldehydes in pretreated lignocellulosic feedstocks and evaluation of their inhibitory effects on yeast. J. Agric. Food Chem. 2015, 63, 9747–9754. [Google Scholar] [CrossRef] [PubMed]
- Stagge, S.; Cavka, A.; Jönsson, L.J. Identification of benzoquinones in pretreated lignocellulosic feedstocks and inhibitory effects on yeast. AMB Express 2015, 5, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, C.; Wu, G.; Wang, Z.; Stagge, S.; Jönsson, L.J. Formation of microbial inhibitors in steam-explosion pretreatment of softwood impregnated with sulfuric acid and sulfur dioxide. Bioresour. Technol. 2018, 262, 242–250. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhang, X.; Gregg, D.J.; Saddler, J.N. Effects of sugar inhibition on cellulases and β-glucosidase during enzymatic hydrolysis of softwood substrates. Appl. Biochem. Biotechnol. 2004, 115, 1115–1126. [Google Scholar] [CrossRef]
- Malgas, S.; Van Dyk, J.S.; Abboo, S.; Pletschke, B.I. The inhibitory effects of various substrate pre-treatment by-products and wash liquors on mannanolytic enzymes. J. Mol. Catal. B Enzym. 2016, 123, 132–140. [Google Scholar] [CrossRef]
- Zhai, R.; Hu, J.; Saddler, J.N. What are the major components in steam pretreated lignocellulosic biomass that inhibit the efficacy of cellulase enzyme mixtures? ACS Sustain. Chem. Eng. 2016, 4, 3429–3436. [Google Scholar] [CrossRef]
- Pienkos, P.T.; Zhang, M. Role of pretreatment and conditioning processes on toxicity of lignocellulosic biomass hydrolysates. Cellulous 2009, 16, 743–762. [Google Scholar] [CrossRef]
- Alriksson, B.; Cavka, A.; Jönsson, L.J. Improving the fermentability of enzymatic hydrolysates of lignocellulose through chemical in-situ detoxification with reducing agents. Bioresour. Technol. 2011, 102, 1254–1263. [Google Scholar] [CrossRef]
- Cavka, A.; Alriksson, B.; Ahnlund, M.; Jönsson, L.J. Effect of sulfur oxyanions on lignocellulose-derived fermentation inhibi-tors. Biotechnol. Bioeng. 2011, 108, 2592–2599. [Google Scholar] [CrossRef]
- Cavka, A.; Jönsson, L.J. Detoxification of lignocellulosic hydrolysates using sodium borohydride. Bioresour. Technol. 2013, 136, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Overend, R.P.; Chornet, E. Fractionation of lignocellulosics by steam-aqueous pretreatments. Philos. Trans. R. Soc. Lond. Ser. A Math. Phys. Sci. 1987, 321, 523–536. [Google Scholar] [CrossRef]
- Chum, H.L.; Johnson, D.K.; Black, S.K.; Overend, R.P. Pretreatment-catalyst effects and the combined severity parameter. Appl. Biochem. Biotechnol. 1990, 24–25, 1–14. [Google Scholar] [CrossRef]
- Sluiter, A.; Hyman, D.; Payne, C.; Wolfe, J. Determination of Insoluble Solids in Pretreated Biomass Material; National Renewable Energy Laboratory: Golden, CO, USA, 2008. [Google Scholar]
- Singleton, V.L.; Orhofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by measure of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Wang, Z.; Wu, G.; Jönsson, L.J. Effects of impregnation of softwood with sulfuric acid and sulfur dioxide on chemical and physical characteristics, enzymatic digestibility, and fermentability. Bioresour. Technol. 2018, 247, 200–208. [Google Scholar] [CrossRef]
- Du, B.; Sharma, L.N.; Becker, C.; Chen, S.-F.; Mowery, R.A.; Van Walsum, G.P.; Chambliss, C.K. Effect of varying feedstock-pretreatment chemistry combinations on the formation and accumulation of potentially inhibitory degradation products in biomass hydrolysates. Biotechnol. Bioeng. 2010, 107, 430–440. [Google Scholar] [CrossRef]
- Larsson, S.; Palmqvist, E.; Hahn-Hägerdal, B.; Tengborg, C.; Stenberg, K.; Zacchi, G.; Nilvebrant, N.-O. The generation of fermentation inhibitors during dilute acid hydrolysis of softwood. Enzym. Microb. Technol. 1999, 24, 151–159. [Google Scholar] [CrossRef]
- Wang, L.-Q.; Cai, L.-Y.; Ma, Y.-L. Study on inhibitors from acid pretreatment of corn stalk on ethanol fermentation by alcohol yeast. RSC Adv. 2020, 10, 38409–38415. [Google Scholar] [CrossRef]
- Guo, X.; Cavka, A.; Jönsson, L.J.; Hong, F. Comparison of methods for detoxification of spruce hydrolysate for bacterial cel-lulose production. Microb. Cell Fact. 2013, 12, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralph, J.; Lapierre, C.; Boerjan, W. Lignin structure and its engineering. Curr. Opin. Biotechnol. 2019, 56, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Veguta, V.L.; Stevanic, J.S.; Lindström, M.; Salmén, L. Thermal and alkali stability of sodium dithionite studied using ATR-FTIR spectroscopy. BioResources 2017, 12, 2496–2506. [Google Scholar]
- Patel, A.K.; Singhania, R.R.; Sim, S.J.; Pandey, A. Thermostable cellulases: Current status and perspectives. Bioresour. Technol. 2019, 279, 385–392. [Google Scholar] [CrossRef]
- Favaro, L.; Jansen, T.; Van Zyl, W.H. Exploring industrial and natural Saccharomyces cerevisiae strains for the bio-based economy from biomass: The case of bioethanol. Crit. Rev. Biotechnol. 2019, 39, 800–816. [Google Scholar] [CrossRef]
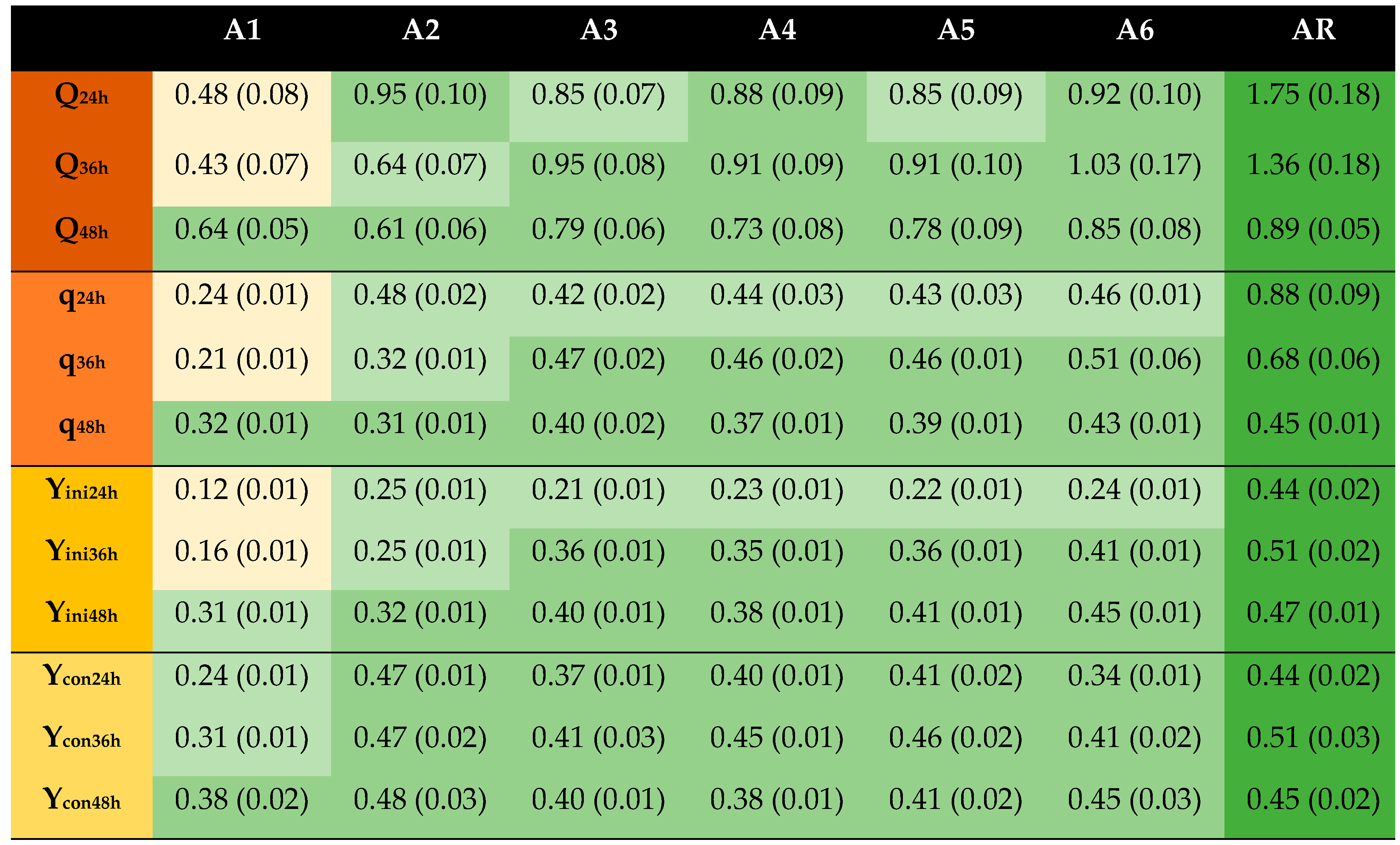
 , reference;
, reference;  , ≥67% of reference;
, ≥67% of reference;  , <67% to ≥33% of reference;
, <67% to ≥33% of reference;  , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g biomass × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, reference; , ≥67% of reference; , <67% to ≥33% of reference; , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g biomass × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g biomass × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, reference; , ≥67% of reference; , <67% to ≥33% of reference; , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g biomass × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
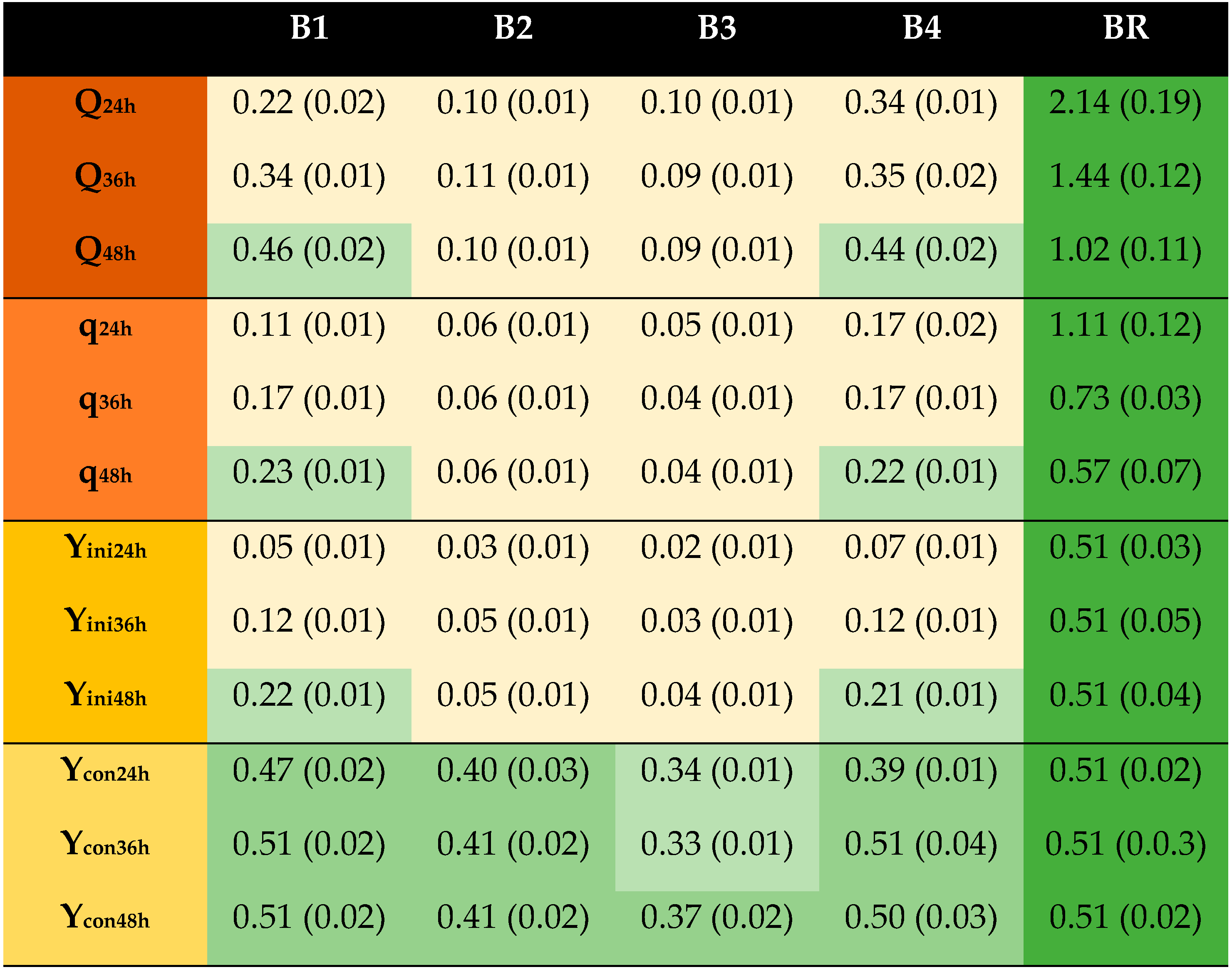
 , reference;
, reference;  , ≥67% of reference;
, ≥67% of reference;  , <67% to ≥33% of reference;
, <67% to ≥33% of reference;  , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, reference; , ≥67% of reference; , <67% to ≥33% of reference; , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, reference; , ≥67% of reference; , <67% to ≥33% of reference; , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
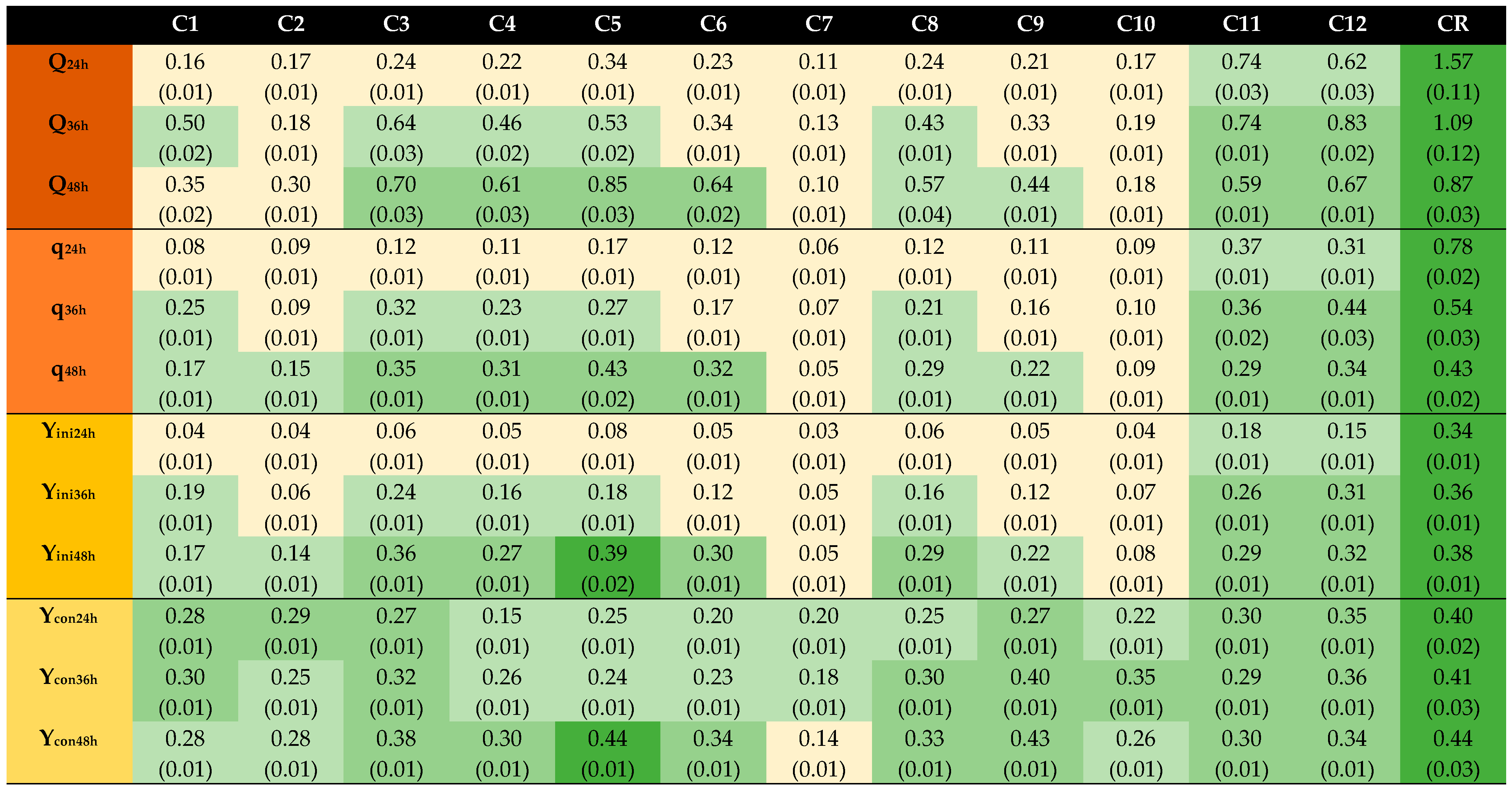
 , reference;
, reference;  , ≥67% of reference;
, ≥67% of reference;  , <67% to ≥33% of reference;
, <67% to ≥33% of reference;  , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, reference; , ≥67% of reference; , <67% to ≥33% of reference; , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.
, reference; , ≥67% of reference; , <67% to ≥33% of reference; , <33% of reference. Q, g EtOH/(L × h); q, g EtOH/(g × h); Yini, g EtOH/g initial glucose; Ycon, g EtOH/g consumed glucose.

{kind=link}
{kind=link}
{kind=link}
| Codification | Na2S2O4 (mM) | T (°C) | Time (min) | pH |
|---|---|---|---|---|
| A1 | - | 23 | 10 | 5.5 |
| A2 | 5 | 23 | 10 | 2.5 |
| A3 | 5 | 23 | 10 | 5.5 |
| A4 | 5 | 23 | 10 | 8.5 |
| A5 | 5 a | 23 | 10 | 5.5 |
| A6 | 5 | 75 | 10 | 5.5 |
| AR | Glucose reference | |||
| B1 | 5 | 75 | 10 | 5.5 |
| B2 | - | 75 | 10 | 5.5 |
| B3 | - | 23 | 10 | 5.5 |
| B4 | 5 | 23 | 10 | 5.5 |
| BR | Glucose reference | |||
| C1 | 5 | 50 | 10 | 5.5 |
| C2 | 8.7 | 50 | 10 | 5.5 |
| C3 | 12.5 | 50 | 10 | 5.5 |
| C4 | 5 | 75 | 10 | 5.5 |
| C5 | 8.7 | 75 | 10 | 5.5 |
| C6 | 12.5 | 75 | 10 | 5.5 |
| C7 | 5 | 95 | 10 | 5.5 |
| C8 | 8.7 | 95 | 10 | 5.5 |
| C9 | 12.5 | 95 | 10 | 5.5 |
| C10 | 5 | 110 | 10 | 5.5 |
| C11 | 8.7 | 110 | 10 | 5.5 |
| C12 | 12.5 | 110 | 10 | 5.5 |
| CR | Glucose reference | |||
| Substance | Concentration |
|---|---|
| Aliphatic aldehydes (mM) | |
| Acetaldehyde | 1.7 (0.2) |
| Formaldehyde | 6.1 (0.4) |
| Aliphatic carboxylic acids (mM) | |
| Acetic acid | 95 (3) |
| Formic acid | 21 (2) |
| Levulinic acid | 19 (2) |
| Benzoquinones (μM) | |
| p-Benzoquinone | ND b |
| Furan aldehydes (mM) | |
| Furfural | 19.3 (0.9) |
| HMF | 20.1 (0.9) |
| Group analyses | |
| TAC (AU) c | 0.26 (0.01) |
| TCAC (mM) d | 153 (2) |
| Total phenolics (g/L) e | 1.6 (0.1) |
| Phenols (μM) | |
| Acetovanillone | 9.8 (1.0) |
| Coniferyl aldehyde | 121 (10) |
| p-Coumaraldehyde | 1.6 (0.1) |
| 4-Hydroxybenzaldehyde | 35.1 (9.4) |
| Syringaldehyde | 0.12 (0.01) |
| Vanillin | 430 (120) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilanidis, D.; Stagge, S.; Alriksson, B.; Jönsson, L.J. Factors Affecting Detoxification of Softwood Enzymatic Hydrolysates Using Sodium Dithionite. Processes 2021, 9, 887. https://doi.org/10.3390/pr9050887
Ilanidis D, Stagge S, Alriksson B, Jönsson LJ. Factors Affecting Detoxification of Softwood Enzymatic Hydrolysates Using Sodium Dithionite. Processes. 2021; 9(5):887. https://doi.org/10.3390/pr9050887
Chicago/Turabian StyleIlanidis, Dimitrios, Stefan Stagge, Björn Alriksson, and Leif J. Jönsson. 2021. "Factors Affecting Detoxification of Softwood Enzymatic Hydrolysates Using Sodium Dithionite" Processes 9, no. 5: 887. https://doi.org/10.3390/pr9050887
APA StyleIlanidis, D., Stagge, S., Alriksson, B., & Jönsson, L. J. (2021). Factors Affecting Detoxification of Softwood Enzymatic Hydrolysates Using Sodium Dithionite. Processes, 9(5), 887. https://doi.org/10.3390/pr9050887