
Selection of Lactic Acid Bacteria with In Vitro Probiotic-Related Characteristics from the Cactus Pilosocereus gounellei (A. Weber ex. K. Schum.) Bly. ex Rowl


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Preliminary Identification of LAB
2.2. Evaluation of Safety Characteristics
2.2.1. Hemolytic Activity
2.2.2. Mucinolytic Activity
2.2.3. Evaluation of Antibiotic Susceptibility
2.3. Evaluation of In Vitro Physiological Functionality Properties
2.3.1. Acid and Bile Salt Tolerance
2.3.2. Cell Surface Hydrophobicity
2.3.3. Autoaggregation and Coaggregation Capacity
2.3.4. Evaluation of the Antagonistic Activity against Pathogens
2.4. Evaluation of In Vitro Technological Properties
2.4.1. Proteolytic Activity
2.4.2. Diacetyl Production
2.4.3. Exopolysaccharide Production
2.4.4. Tolerance to Sodium Chloride
2.4.5. Ability to Grow at Different Temperatures
2.5. Selection of Isolates with the Most Promising In Vitro Characteristics for Probiotic Using Multivariate Analyze
2.6. Survival of Selected Isolates in Different Food Matrices
2.7. Identification of Selected Isolates by Sequence Analysis of 16S-rRNA Gene
2.8. Statistical Analysis
3. Results
3.1. Isolation and Preliminary Identification of Isolates
3.2. Safety Analysis
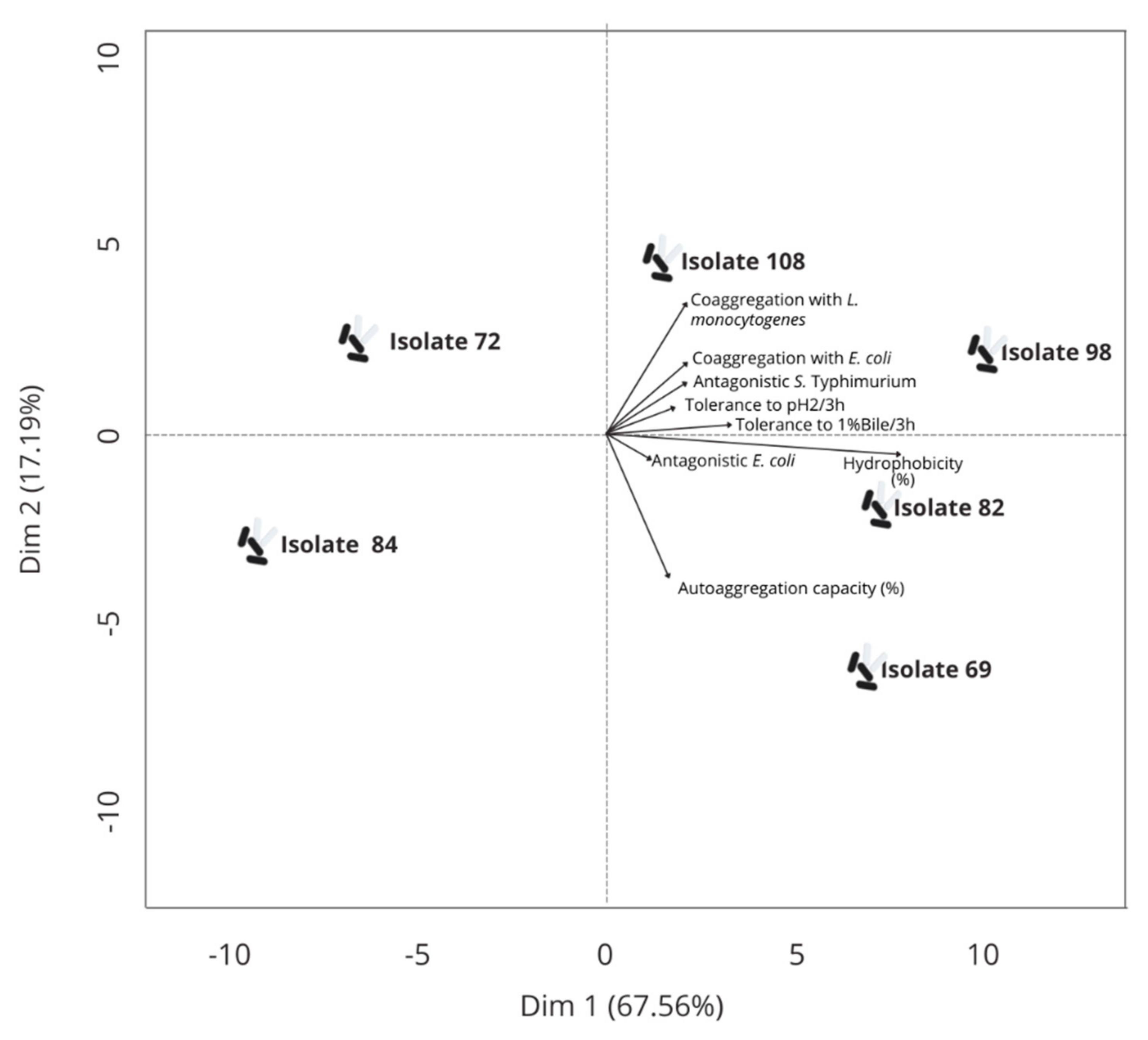
3.3. Selection of LAB Isolates Based on In Vitro Physiological Functionality Characteristics
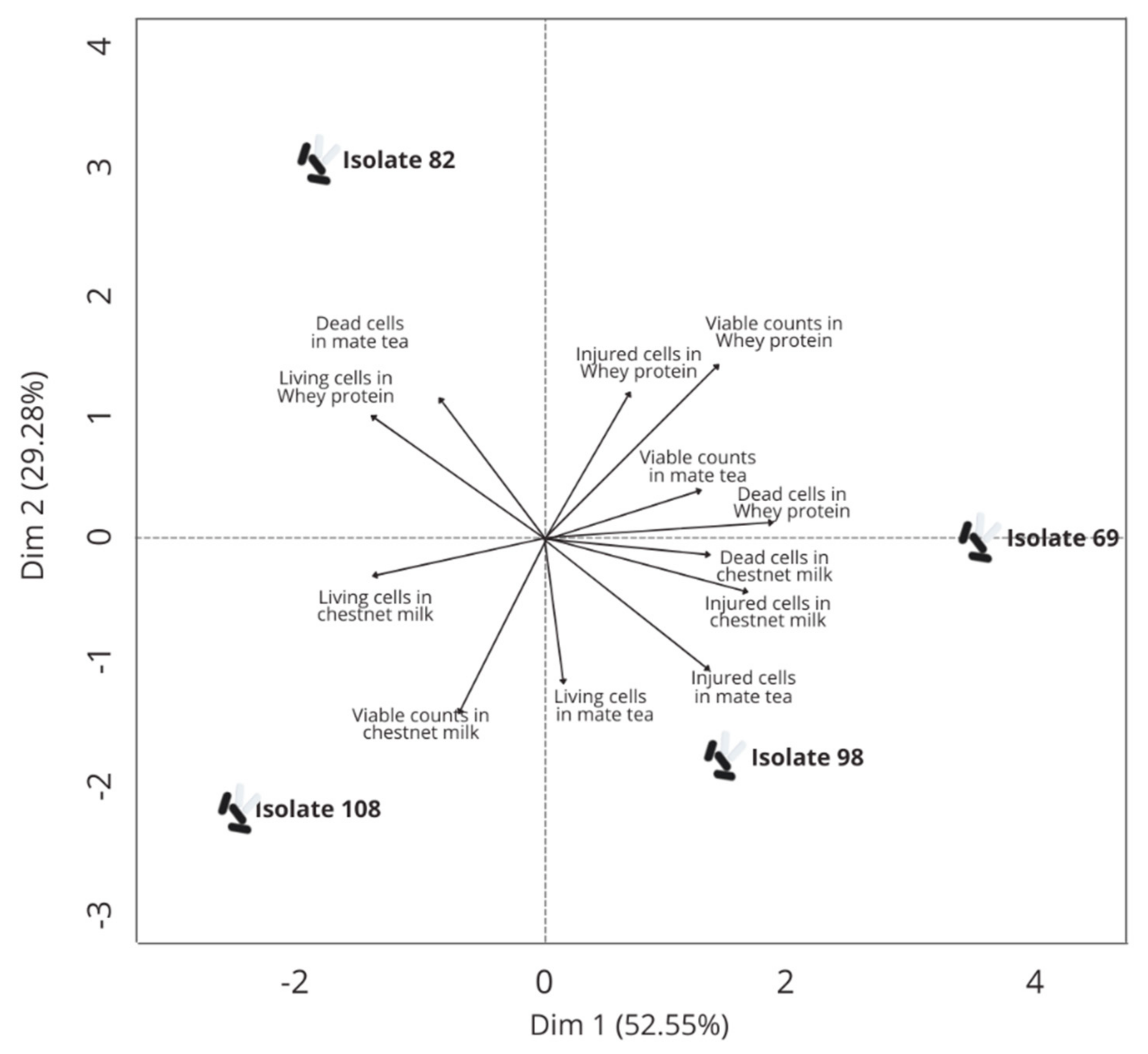
3.4. Technological Properties and Survival of Selected Isolates in Different Food Matrices
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, A.; Sharma, N. Probiotic Potential of Lactic Acid Bacteria Ch-2 Isolated from Chuli Characterization of Potential Probiotic Lactic Acid Bacteria- Pediococcus acidilactici Ch-2 Isolated from Chuli- A Traditional Apricot Product of Himalayan Region for the Production of Novel Bioactive Compounds with Special Therapeutic Properties. J. Food Microbiol. Saf. Hyg. 2017, 2, 1–11. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Bielecka, M. Probiotics in Food. Chem. Funct. Prop. Food Compon. 2006, 413–426. [Google Scholar] [CrossRef]
- Kakelar, H.M.; Barzegari, A.; Hanifian, S.; Barar, J.; Omidi, Y. Isolation and molecular identification of Lactobacillus with probiotic potential from abomasums driven rennet. Food Chem. 2019, 272, 709–714. [Google Scholar] [CrossRef]
- Di Cagno, R.; Coda, R.; De Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Verón, H.E.; Di Risio, H.D.; Isla, M.I.; Torres, S. Isolation and selection of potential probiotic lactic acid bacteria from Opuntia ficus-indica fruits that grow in Northwest Argentina. LWT 2017, 84, 231–240. [Google Scholar] [CrossRef]
- de Carvalho, P.O.A.A.; Guerra, G.C.B.; Borges, G.D.S.C.; Bezerril, F.F.; Sampaio, K.B.; Ribeiro, T.S.; Pacheco, M.T.B.; Milani, R.F.; Goldbeck, R.; Ávila, P.F.; et al. Nutritional potential and bioactive compounds of xique-xique juice: An unconventional food plant from Semiarid Brazilian. J. Food Process. Preserv. 2021, 45, e15265. [Google Scholar] [CrossRef]
- Bezerril, F.F.; Souza, M.D.F.V.D.; Lima, M.D.S.; Pacheco, M.T.B.; de Carvalho, P.O.A.A.; Sampaio, K.B.; de Sousa, Y.R.F.; Milani, R.F.; Goldbeck, R.; Borges, G.D.S.C.; et al. Physicochemical characteristics and bioactive compounds of the Xique-xique (Pilosocereus gounellei) cactus from Caatinga Brazilian: Are they nutritive and functional? J. Food Meas. Charact. 2021, 15, 3284–3297. [Google Scholar] [CrossRef]
- Ribeiro, T.S.; Sampaio, K.B.; Menezes, F.N.D.D.; de Assis, P.O.A.; Lima, M.D.S.; de Oliveira, M.E.G.; de Souza, E.L.; Queiroga, R.D.C.R.D.E. In vitro evaluation of potential prebiotic effects of a freeze-dried juice from Pilosocereus gounellei (A. Weber ex K. Schum. Bly. Ex Rowl) cladodes, an unconventional edible plant from Caatinga biome. 3 Biotech 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.A.; Dantas, R.L.; Véras, C.D.S.; Alves, R.E.; Silva, S.D.M. Physical and physicochemical characteristics, bioactive compounds and antioxidant activity of xiquexique (Pilosocereus gounellei (A. Weber ex K. Schum.)) fruit. Semin. Ciênc. Agrár. 2018, 39, 1969–1980. [Google Scholar] [CrossRef]
- Tangyu, M.; Muller, J.; Bolten, C.J.; Wittmann, C. Fermentation of plant-based milk alternatives for improved flavour and nutritional value. Appl. Microbiol. Biotechnol. 2019, 103, 9263–9275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimentel, T.C.; da Costa, W.K.A.; Barão, C.E.; Rosset, M.; Magnani, M. Vegan probiotic products: A modern tendency or the newest challenge in functional foods. Food Res. Int. 2021, 140, 110033. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Koley, H.; Dutta, S.; Bhowal, J. Hepatoprotective effects of synbiotic soy yogurt on mice fed a high-cholesterol diet. Nutrition 2019, 1, 36–44. [Google Scholar] [CrossRef]
- Garcia, E.F.; de Oliveira Araújo, A.; Luciano, W.A.; de Albuquerque, T.M.R.; de Oliveira Arcanjo, N.M.; Madruga Lima, M.S.; Magnani, M.; Saarela, M.; de Souza, E.L. The performance of five fruit-derived and freeze-dried potentially probiotic Lactobacillus strains in apple, orange, and grape juices. J. Sci. Food Agric. 2018, 98, 5000–5010. [Google Scholar] [CrossRef]
- Amorim, J.C.; Piccoli, R.H.; Duarte, W.F. Probiotic potential of yeasts isolated from pineapple and their use in the elaboration of potentially functional fermented beverages. Food Res. Int. 2018, 107, 518–527. [Google Scholar] [CrossRef]
- American Public Health (APHA). Compendium for the Microbiological Examination of Foods; American Public Health: Washington, DC, USA, 2015. [Google Scholar]
- Garcia, E.F.; Luciano, W.A.; Xavier, D.E.; da Costa, W.C.; de Sousa Oliveira, K.; Franco, O.L.; Morais Júnior, M.A.; Lucena, B.T.L.; Picão, R.C.; Magnani, M.; et al. Identification of Lactic Acid Bacteria in Fruit Pulp Processing Byproducts and Potential Probiotic Properties of Selected Lactobacillus Strains. Front. Microbiol. 2016, 7, 1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.G.; Tsakalidou, E.; Nychas, G.-J.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rodríguez-Aparicio, L.; Rúa, J.; Martínez-Blanco, H.; Navasa, N.; García-Armesto, M.R.; Ferrero, M.Á. In vitro evaluation of physiological probiotic properties of different lactic acid bacteria strains of dairy and human origin. J. Funct. Foods 2012, 4, 531–541. [Google Scholar] [CrossRef]
- Awasti, N.; Tomar, S.; Pophaly, S.D.; Lule, V.K.; Singh, T.P.; Anand, S. Probiotic and functional characterization of bifidobacteria of Indian human origin. J. Appl. Microbiol. 2016, 120, 1021–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI (Clinical and Laboratory Standards Institute). Performance standards for antimicrobial susceptibility testing. Clin. Microbiol. Newsl. 2019, 49. [Google Scholar] [CrossRef]
- De Albuquerque, T.M.R.; Garcia, E.F.; Araújo, A.D.O.; Magnani, M.; Saarela, M.; De Souza, E.L. In Vitro Characterization of Lactobacillus Strains Isolated from Fruit Processing By-Products as Potential Probiotics. Probiotics Antimicrob. Proteins 2017, 10, 704–716. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, K.M.O.; Vieira, A.D.S.; Rocha, C.R.C.; do Nascimento, J.C.F.; de Souza Lopes, A.C.; Bruno, L.M.; Carvalho, J.D.G.; Franco, B.D.G.M.; Todorov, S.D. Brazilian artisanal cheeses as a source of beneficial Enterococcus faecium strains: Characterization of the bacteriocinogenic potential. Ann. Microbiol. 2014, 64, 1463–1471. [Google Scholar] [CrossRef]
- Todorov, S.D.; Botes, M.; Guigas, C.; Schillinger, U.; Wiid, I.; Wachsman, M.B.; Holzapfel, W.H.; Dicks, L.M.T. Boza, a natural source of probiotic lactic acid bacteria. J. Appl. Microbiol. 2008, 104, 465–477. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.; Stanton, C.; Ross, P.; Dapkevicius, M.; Silva, C. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Bujňáková, D.; Straková, E. Safety, probiotic and technological properties of Lactobacilli isolated from unpasteurised ovine and caprine cheeses. Ann. Microbiol. 2017, 67, 813–826. [Google Scholar] [CrossRef]
- Reale, A.; Di Renzo, T.; Rossi, F.; Zotta, T.; Iacumin, L.; Preziuso, M.; Parente, E.; Sorrentino, E.; Coppola, R. Tolerance of Lactobacillus casei, Lactobacillus paracasei and Lactobacillus rhamnosus strains to stress factors encountered in food processing and in the gastro-intestinal tract. LWT 2015, 60, 721–728. [Google Scholar] [CrossRef]
- Paparella, A.; Taccogna, L.; Aguzzi, I.; Lopez, C.C.; Serio, A.; Marsilio, F.; Suzzi, G. Flow cytometric assessment of the antimicrobial activity of essential oils against Listeria monocytogenes. Food Control. 2008, 19, 1174–1182. [Google Scholar] [CrossRef]
- Pedrosa, G.T.D.S.; de Souza, E.L.; de Melo, A.N.F.; Almeida, E.T.D.C.; Guedes, J.P.D.S.; de Carvalho, R.J.; Pagán, R.; Magnani, M. Physiological alterations involved in inactivation of autochthonous spoilage bacteria in orange juice caused by Citrus essential oils and mild heat. Int. J. Food Microbiol. 2020, 334, 108837. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qian, P.Y. Conservative Fragments in Bacterial 16S rRNA Genes and Primer Design for 16S Ribosomal DNA Amplicons in Metagenomic Studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 15 July 2021).
- Doan, N.; Van Hoorde, K.; Cnockaert, M.; De Brandt, E.; Aerts, M.; Le Thanh, B.; Vandamme, P. Validation of MALDI-TOF MS for rapid classification and identification of lactic acid bacteria, with a focus on isolates from traditional fermented foods in Northern Vietnam. Lett. Appl. Microbiol. 2012, 55, 265–273. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Vincentini, O.; Lanera, A.; Cavoski, I.; Gobbetti, M. Exploitation of Leuconostoc mesenteroides strains to improve shelf life, rheological, sensory and functional features of prickly pear (Opuntia ficusindica L.) fruit puree. Food Microbiol. 2016, 59, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.A.; Santos, E.M.; Araújo, G.G.L.; Oliveira, J.S.; Pinho, R.M.A.; Zanine, A.D.M.; Souza, A.F.N.; Macedo, A.J.S.; Neto, J.M.C.; Nascimento, T.V.C. Isolation and identification of lactic acid bacteria in fresh plants and in silage from Opuntia and their effects on the fermentation and aerobic stability of silage. J. Agric. Sci. 2019, 157, 684–692. [Google Scholar] [CrossRef]
- Zhou, J.; Gopal, P.; Gill, H. Potential probiotic lactic acid bacteria Lactobacillus rhamnosus (HN001), Lactobacillus acidophilus (HN017) and Bifidobacterium lactis (HN019) do not degrade gastric mucin in vitro. Int. J. Food Microbiol. 2001, 63, 81–90. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA—Opinion of the Scientific Committee. EFSA J. 2007, 5, 587. [CrossRef]
- Akman, P.K.; Ozulku, G.; Tornuk, F.; Yetim, H. Potential probiotic lactic acid bacteria isolated from fermented gilaburu and shalgam beverages. LWT 2021, 149, 111705. [Google Scholar] [CrossRef]
- Speranza, B.; Campaniello, D.; Petruzzi, L.; Altieri, C.; Sinigaglia, M.; Bevilacqua, A.; Corbo, M.R. The Inoculation of Probiotics In Vivo Is a Challenge: Strategies to Improve Their Survival, to Avoid Unpleasant Changes, or to Enhance Their Performances in Beverages. Beverages 2020, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Bezerra, T.K.A.; de Araujo, A.R.R.; do Nascimento, E.S.; de Matos Paz, J.E.; Gadelha, C.A.; Gadelha, T.S.; Pacheco, M.T.B.; Queiroga, R.C.R.E.; Oliveira, M.E.G.; Madruga, M.S. Proteolysis in goat “coalho” cheese supplemented with probiotic lactic acid bacteria. Food Chem. 2016, 196, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Gregoret, V.; Perezlindo, M.; Vinderola, G.; Reinheimer, J.; Binetti, A. A comprehensive approach to determine the probiotic potential of human-derived Lactobacillus for industrial use. Food Microbiol. 2013, 34, 19–28. [Google Scholar] [CrossRef]
- Gomand, F.; Borges, F.; Burgain, J.; Guerin, J.; Revol-Junelles, A.-M.; Gaiani, C. Food Matrix Design for Effective Lactic Acid Bacteria Delivery. Annu. Rev. Food Sci. Technol. 2019, 10, 285–310. [Google Scholar] [CrossRef]
- Gonzalez-Gil, F.; Diaz-Sanchez, S.; Pendleton, S.; Andino, A.; Zhang, N.; Yard, C.; Crilly, N.; Harte, F.; Hanning, I. Yerba mate enhances probiotic bacteria growth in vitro but as a feed additive does not reduce Salmonella Enteritidis colonization in vivo. Poult. Sci. 2014, 93, 434–440. [Google Scholar] [CrossRef]
- International Standards Organization (ISO); International Dairy Federation (IDF). Milk and Milk Products—Starter Cultures, Probiotics and Fermented Products—Quantification of Lactic Acid Bacteria by Flow Cytometry; ISO: Geneva, Switzerland, 2015; ISO 193442015 IDF 232:20. [Google Scholar]
- Wilkinson, M.G. Flow cytometry as a potential method of measuring bacterial viability in probiotic products: A review. Trends Food Sci. Technol. 2018, 78, 1–10. [Google Scholar] [CrossRef]
- Truong, V.K.; Dupont, M.; Elbourne, A.; Gangadoo, S.; Pathirannahalage, P.R.; Cheeseman, S.; Chapman, J.; Cozzolino, D. From Academia to Reality Check: A Theoretical Framework on the Use of Chemometric in Food Sciences. Foods 2019, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Ye, P.; Wang, J.; Liu, M.; Li, P.; Gu, Q. Purification and characterization of a novel bacteriocin from Lactobacillus paracasei ZFM54. LWT 2021, 143, 111125. [Google Scholar] [CrossRef]
- Aspri, M.; Papademas, P.; Tsaltas, D. Review on Non-Dairy Probiotics and Their Use in Non-Dairy Based Products. Fermentation 2020, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Klein, G.; Pack, A.; Bonaparte, C.; Reuter, G. Taxonomy and physiology of probiotic lactic acid bacteria. Int. J. Food Microbiol. 1998, 41, 103–125. [Google Scholar] [CrossRef]
- Holzapfel, W.H.; Haberer, P.; Geisen, R.; Bjorkroth, J.; Schillinger, U. Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr. 2001, 73, 365s–373s. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Isolate | Chloramphenicol (30 μg/Disc) | Erythromycin (15 μg/Disc) | Tobramycin (10 μg/Disc) | Tetracycline (30 μg/Disc) | Clindamycin (2 μg/Disc) | Ampicillin (10 μg/Disc) | Vancomycin (30 μg/Disc) | Gentamycin (10 μg/Disc) | Streptomycin (300 μg/Disc) |
|---|---|---|---|---|---|---|---|---|---|
| 69 | S | S | S | S | S | S | S | S | S |
| 72 | S | S | S | S | S | S | S | S | S |
| 79 | S | S | S | S | R | S | S | S | S |
| 80 | S | S | S | S | S | R | S | S | S |
| 82 | S | S | S | S | S | S | S | S | S |
| 84 | S | S | S | S | S | S | S | S | S |
| 98 | S | S | S | S | S | S | S | S | S |
| 108 | S | S | S | S | S | S | S | S | S |
| Isolate | Diacetyl Production * | Proteolytic ** | EPS Production (mg/L) | Capability to Grow at Different Temperatures (°C) ** | Tolerance to NaCl | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 35 | 35 | 37 | 40 | 45 | 1% | 2% | 3% | 4% | 5% | ||||
| 69 | ++ | + | 75.1 ± 1.5 d | + | + | + | + | + | 89.5 ± 0.9 a | 86.2 ± 0.6 b | 88.6 ± 0.3 c | 20.1 ± 0.1 c | 17.3 ± 0.3 c |
| 82 | ++ | + | 90.6 ± 0.9 a | + | + | + | + | + | 99.5 ± 0.3 a | 99.3 ± 0.2 a | 96.8 ± 0.4 a | 94.4 ± 0.6 a | 11.5 ± 0.4 d |
| 98 | ++ | + | 88.1 ± 0.4 b | + | + | + | + | + | 98.9 ± 0.1 a | 97.8 ± 0.9 a | 90.4 ± 0.1 b | 87.7 ± 0.1 b | 52.8 ± 0.1 a |
| 108 | ++ | + | 78.3 ± 0.3 c | + | + | + | + | + | 99.3 ± 0.7 a | 99.2 ± 0.3 a | 95.7 ± 0.7 a | 85.9 ± 0.7 b | 28.4 ± 0.1 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sampaio, K.B.; de Albuquerque, T.M.R.; Rodrigues, N.P.A.; de Oliveira, M.E.G.; de Souza, E.L. Selection of Lactic Acid Bacteria with In Vitro Probiotic-Related Characteristics from the Cactus Pilosocereus gounellei (A. Weber ex. K. Schum.) Bly. ex Rowl. Foods 2021, 10, 2960. https://doi.org/10.3390/foods10122960
Sampaio KB, de Albuquerque TMR, Rodrigues NPA, de Oliveira MEG, de Souza EL. Selection of Lactic Acid Bacteria with In Vitro Probiotic-Related Characteristics from the Cactus Pilosocereus gounellei (A. Weber ex. K. Schum.) Bly. ex Rowl. Foods. 2021; 10(12):2960. https://doi.org/10.3390/foods10122960
Chicago/Turabian StyleSampaio, Karoliny Brito, Thatyane Mariano Rodrigues de Albuquerque, Noádia Priscila Araújo Rodrigues, Maria Elieidy Gomes de Oliveira, and Evandro Leite de Souza. 2021. "Selection of Lactic Acid Bacteria with In Vitro Probiotic-Related Characteristics from the Cactus Pilosocereus gounellei (A. Weber ex. K. Schum.) Bly. ex Rowl" Foods 10, no. 12: 2960. https://doi.org/10.3390/foods10122960
APA StyleSampaio, K. B., de Albuquerque, T. M. R., Rodrigues, N. P. A., de Oliveira, M. E. G., & de Souza, E. L. (2021). Selection of Lactic Acid Bacteria with In Vitro Probiotic-Related Characteristics from the Cactus Pilosocereus gounellei (A. Weber ex. K. Schum.) Bly. ex Rowl. Foods, 10(12), 2960. https://doi.org/10.3390/foods10122960