
Volatile Compounds in Pulses: A Review




Abstract
:1. Introduction
2. Origins of Volatile Compounds in Pulses
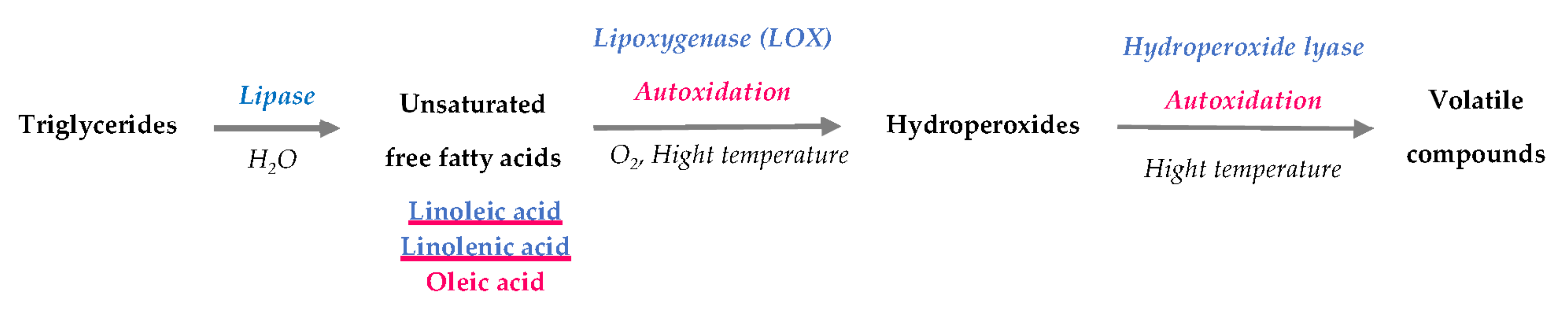
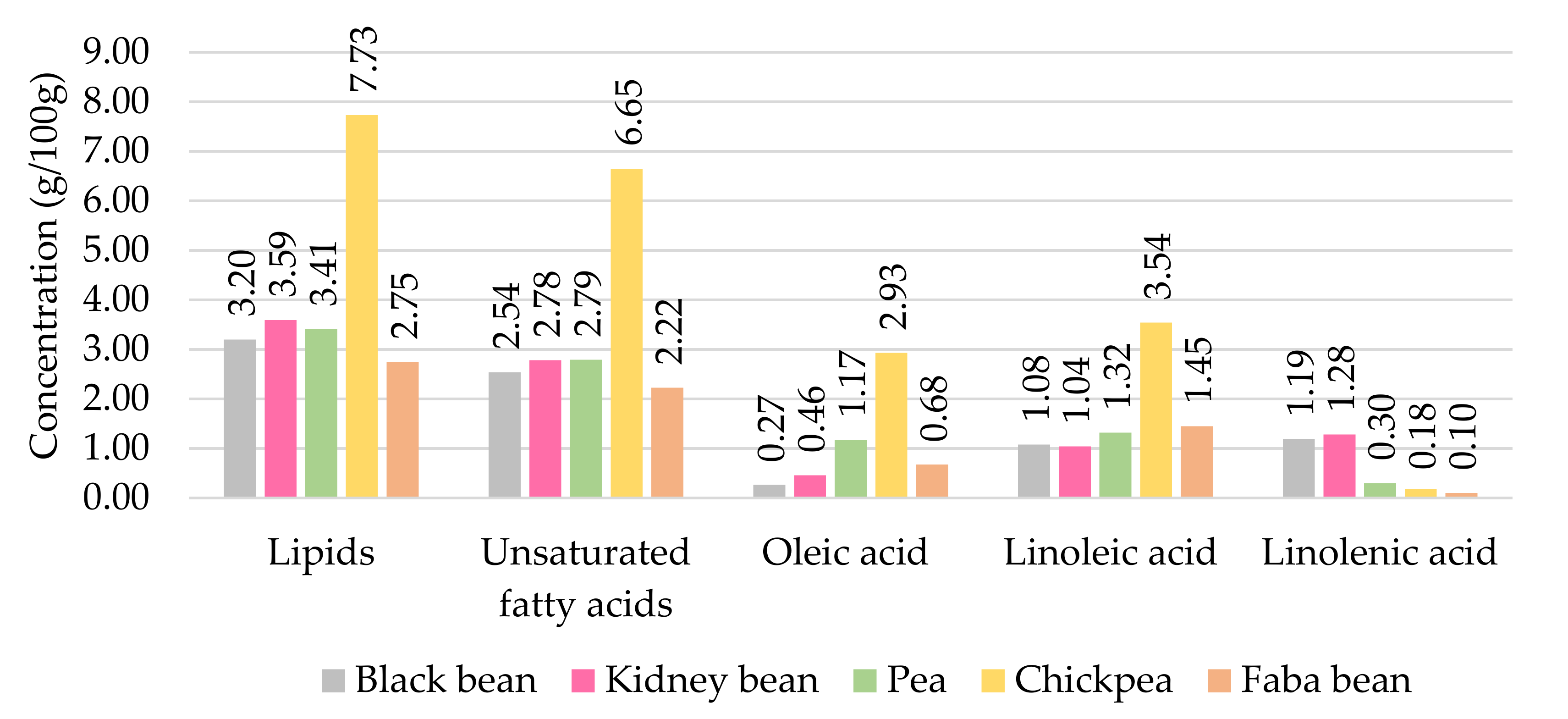
2.1. Oxidation of Unsaturated Free Fatty Acids
2.1.1. Triglyceride Hydrolysis
2.1.2. Oxidation of Free Fatty Acids
- Initiation: RH → R• + H•
- Propagation: R• + O2 → ROO•ROO• + RH → ROOH + R•
- Termination: ROO• + ROO• → Non-radical products
2.1.3. Degradation of Hydroperoxides and Formation of Volatile Compounds (Secondary Products)
2.2. Degradation of Free Amino Acids
2.2.1. Biodegradation
2.2.2. Degradation by Microorganisms
2.2.3. Maillard Reactions
2.3. Degradation of Carotenoids
3. Extraction, Separation, Identification, and Semi-Quantification Methods
4. Identification and Quantification of Volatile Compounds in Pulses
4.1. Aromatic Hydrocarbons
4.2. Aldehydes
4.3. Alkanes and Alkenes
4.4. Alcohols
4.5. Ketones
4.6. Acids
4.7. Esters
4.8. Pyrazines
4.9. Terpenes
4.10. Furans
4.11. Lactones
4.12. Other Volatiles
5. Odour-Active Compounds in Pulses
6. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pedrosa, M.M.; Guillamón, E.; Arribas, C. Autoclaved and Extruded Legumes as a Source of Bioactive Phytochemicals: A Review. Foods 2021, 10, 379. [Google Scholar] [CrossRef]
- Shevkani, K.; Singh, N.; Chen, Y.; Kaur, A.; Yu, L. Pulse Proteins: Secondary Structure, Functionality and Applications. J. Food Sci. Technol. 2019, 56, 2787–2798. [Google Scholar] [CrossRef] [PubMed]
- Voisin, A.-S.; Guéguen, J.; Huyghe, C.; Jeuffroy, M.-H.; Magrini, M.-B.; Meynard, J.-M.; Mougel, C.; Pellerin, S.; Pelzer, E. Legumes for Feed, Food, Biomaterials and Bioenergy in Europe: A Review. Agron. Sustain. Dev. 2014, 34, 361–380. [Google Scholar] [CrossRef]
- Singh, N. Pulses: An Overview. J. Food Sci. Technol. 2017, 54, 853–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bresciani, A.; Marti, A. Using Pulses in Baked Products: Lights, Shadows, and Potential Solutions. Foods 2019, 8, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roland, W.S.U.; Pouvreau, L.; Curran, J.; van de Velde, F.; de Kok, P.M.T. Flavor Aspects of Pulse Ingredients. Cereal Chem. J. 2017, 94, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Briand, L.; Salles, C. Taste perception and integration. In Flavor From Food to Behaviors, Wellbeing and Health; Etiévant, P., Guichard, E., Salles, C., Voilley, A., Eds.; Woodhead Publishing, Elsevier Ltd.: Duxford, UK, 2016; pp. 101–119. ISBN 978-0-08-100295-7. [Google Scholar]
- Vincenti, S.; Mariani, M.; Alberti, J.-C.; Jacopini, S.; Brunini-Bronzini de Caraffa, V.; Berti, L.; Maury, J. Biocatalytic Synthesis of Natural Green Leaf Volatiles Using the Lipoxygenase Metabolic Pathway. Catalysts 2019, 9, 873. [Google Scholar] [CrossRef] [Green Version]
- Guichard, E.; Genot, C.; Voilley, A.; Brachais, L.; Briand, L.; Brossard, C.; Cayot, N.N.; Chalier, P.; Deleris, I.; Doublier, J.L.; et al. Texture et Flaveur Des. Aliments: Vers Une Conception Maîtrisée; Educagri Editions: Dijon, France, 2012; ISBN 978-2-84444-879-8. [Google Scholar]
- López, M.A.; Vicente, J.; Kulasekaran, S.; Vellosillo, T.; Martínez, M.; Irigoyen, M.L.; Cascón, T.; Bannenberg, G.; Hamberg, M.; Castresana, C. Antagonistic Role of 9-Lipoxygenase-Derived Oxylipins and Ethylene in the Control of Oxidative Stress, Lipid Peroxidation and Plant Defence. Plant. J. 2011, 67, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Azarnia, S.; Boye, J.I.; Warkentin, T.; Malcolmson, L.; Sabik, H.; Bellido, A.S. Volatile Flavour Profile Changes in Selected Field Pea Cultivars as Affected by Crop Year and Processing. Food Chem. 2011, 124, 326–335. [Google Scholar] [CrossRef]
- Sessa, D.J.; Rackis, J.J. Lipid-Derived Flavors of Legume Protein Products. J. Am. Oil Chem. Soc. 1977, 54, 468–473. [Google Scholar] [CrossRef]
- Dundas, D.G.A.; Henderson, H.M.; Eskin, N.A.M. Lipase from Vicia faba Minor. Food Chem. 1978, 3, 171–178. [Google Scholar] [CrossRef]
- Frankel, E.N. Volatile Lipid Oxidation Products. Prog. Lipid Res. 1983, 22, 1–33. [Google Scholar] [CrossRef]
- Yang, Z.; Piironen, V.; Lampi, A.-M. Lipid-Modifying Enzymes in Oat and Faba Bean. Food Res. Int. 2017, 100, 335–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattee, H.E.; Salunkhe, D.K.; Sathe, S.K.; Reddy, N.R. Legume Lipids. Crit. Rev. Food Sci. Nutr. 1982, 17, 97–139. [Google Scholar] [CrossRef] [PubMed]
- Khrisanapant, P.; Kebede, B.; Leong, S.Y.; Oey, I. A Comprehensive Characterisation of Volatile and Fatty Acid Profiles of Legume Seeds. Foods 2019, 8, 651. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Saiki, M.; Yoshida, N.; Tomiyama, Y.; Mizushina, Y. Fatty Acid Distribution in Triacylglycerols and Phospholipids of Broad Beans (Vicia faba). Food Chem. 2009, 112, 924–928. [Google Scholar] [CrossRef]
- Peterson, D.M. Lipase Activity and Lipid Metabolism During Oat Malting. Cereal Chem. J. 1999, 76, 159–163. [Google Scholar] [CrossRef]
- Vliegenthart, J.F.G.; Veldink, G.A.; Boldingh, J. Recent Progress in the Study on the Mechanism of Action of Soybean Lipoxygenase. J. Agric. Food Chem. 1979, 27, 623–626. [Google Scholar] [CrossRef] [Green Version]
- Frankel, E.N. Lipid Oxidation. Prog. Lipid Res. 1980, 19, 1–22. [Google Scholar] [CrossRef]
- Yoon, S.; Klein, B.P. Some Properties of Pea Lipoxygenase Isoenzymes. J. Agric. Food Chem. 1979, 27, 955–962. [Google Scholar] [CrossRef]
- Sanz, L.C.; Perez, A.G.; Olias, J.M. Purification and Catalytic Properties of Chickpea Lipoxygenases. Phytochemistry 1992, 31, 2967–2972. [Google Scholar] [CrossRef]
- Clemente, A.; Olías, R.; Olías, J.M. Purification and Characterization of Broad Bean Lipoxygenase Isoenzymes. J. Agric. Food Chem. 2000, 48, 1070–1075. [Google Scholar] [CrossRef]
- Murray, K.E.; Shipton, J.; Whitfield, F.B.; Last, J.H. The Volatiles of Off-Flavoured Unblanched Green Peas (Pisum sativum). J. Sci. Food Agric. 1976, 27, 1093–1107. [Google Scholar] [CrossRef]
- Spinnler, H.-E. Flavors from amino acids. In Food Flavors, Chemical & Functional Properties of Food Components; Jelen, H., Ed.; CRC Press: Boca Raton, FL, USA, 2011; Volume 20116950, pp. 121–136. ISBN 978-1-4398-1491-8. [Google Scholar]
- Jackson, P.; Boulter, D.; Thurman, D.A. A Comparison of Some Properties of Vicilin and Legumin Isolated from Seeds of Pisum sativum, Vicia faba and Cicer arietinum. N. Phytol. 1969, 68, 25–33. [Google Scholar] [CrossRef]
- Bader, S.; Czerny, M.; Eisner, P.; Buettner, A. Characterisation of Odour-Active Compounds in Lupin Flour. J. Sci. Food Agric. 2009, 89, 2421–2427. [Google Scholar] [CrossRef]
- Czerny, M.; Schieberle, P. Important Aroma Compounds in Freshly Ground Wholemeal and White Wheat Flour Identification and Quantitative Changes during Sourdough Fermentation. J. Agric. Food Chem. 2002, 50, 6835–6840. [Google Scholar] [CrossRef] [PubMed]
- Gallois, A. Les Pyrazines Présentes Dans Les Aliments—État Actuel de Nos Connaissances. Sci. Aliment. 1984, 4, 145–166. [Google Scholar]
- Belitz, H.-D.; Grosch, W. Lipids. In Food Chemistry; Belitz, H.-D., Grosch, W., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 152–236. ISBN 978-3-662-07281-3. [Google Scholar]
- van Boekel, M.A.J.S. Formation of Flavour Compounds in the Maillard Reaction. Biotechnol. Adv. 2006, 24, 230–233. [Google Scholar] [CrossRef] [PubMed]
- Moussou, N.; Corzo-Martínez, M.; Sanz, M.L.; Zaidi, F.; Montilla, A.; Villamiel, M. Assessment of Maillard Reaction Evolution, Prebiotic Carbohydrates, Antioxidant Activity and α-Amylase Inhibition in Pulse Flours. J. Food Sci. Technol. 2017, 54, 890–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, K.E.; Whitfield, F.B. The Occurrence of 3-Alkyl-2-Methoxypyrazines in Raw Vegetables. J. Sci. Food Agric. 1975, 26, 973–986. [Google Scholar] [CrossRef]
- Jakobsen, H.B.; Hansen, M.; Christensen, M.R.; Brockhoff, P.B.; Olsen, C.E. Aroma Volatiles of Blanched Green Peas (Pisum sativum, L.). J. Agric. Food Chem. 1998, 46, 3727–3734. [Google Scholar] [CrossRef]
- Rizzi, G.P. Formation of Methoxypyrazines in Reactions of 2(1H)-Pyrazinones with Naturally Occurring Methylating Agents. J. Agric. Food Chem. 1990, 38, 1941–1944. [Google Scholar] [CrossRef]
- Akkad, R.; Kharraz, E.; Han, J.; House, J.D.; Curtis, J.M. Characterisation of the Volatile Flavour Compounds in Low and High Tannin Faba Beans (Vicia faba Var. minor) Grown in Alberta, Canada. Food Res. Int. 2019, 120, 285–294. [Google Scholar] [CrossRef]
- Bühler, J.M.; Dekkers, B.L.; Bruins, M.E.; van der Goot, A.J. Modifying Faba Bean Protein Concentrate Using Dry Heat to Increase Water Holding Capacity. Foods 2020, 9, 1077. [Google Scholar] [CrossRef] [PubMed]
- Maccarrone, M.; van Aarle, P.G.M.; Veldink, G.A.; Vliegenthart, J.F.G. In Vitro Oxygenation of Soybean Biomembranes by Lipoxygenase-2. Biochim. Biophys. Acta (BBA) Biomembr. 1994, 1190, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Oomah, B.D.; Liang, L.S.Y.; Balasubramanian, P. Volatile Compounds of Dry Beans (Phaseolus vulgaris, L.). Plant. Foods Hum. Nutr. 2007, 62, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Oomah, B.D.; Razafindrainibe, M.; Drover, J.C.G. Headspace Volatile Components of Canadian Grown Low-Tannin Faba Bean (Vicia faba, L.) Genotypes. J. Sci. Food Agric. 2014, 94, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Murat, C.; Bard, M.-H.; Dhalleine, C.; Cayot, N. Characterisation of Odour Active Compounds along Extraction Process from Pea Flour to Pea Protein Extract. Food Res. Int. 2013, 53, 31–41. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, L.; Zhang, X.; Wang, M.; Liu, H.; Zhu, Y. Nutritional Components, Volatile Constituents and Antioxidant Activities of 6 Chickpea Species. Food Biosci. 2021, 41, 100964. [Google Scholar] [CrossRef]
- Wang, Y.; Guldiken, B.; Tulbek, M.; House, J.D.; Nickerson, M. Impact of Alcohol Washing on the Flavour Profiles, Functionality and Protein Quality of Air Classified Pea Protein Enriched Flour. Food Res. Int. 2020, 132, 109085. [Google Scholar] [CrossRef] [PubMed]
- Akkad, R.; Kharraz, E.; Han, J.; House, J.D.; Curtis, J.M. The Effect of Short-Term Storage Temperature on the Key Headspace Volatile Compounds Observed in Canadian Faba Bean Flour. Food Sci. Technol. Int. 2021, 1082013221998843. [Google Scholar] [CrossRef] [PubMed]
- Aulakh, J.S.; Malik, A.K.; Kaur, V.; Schmitt-Kopplin, P. A Review on Solid Phase Micro Extraction—High Performance Liquid Chromatography (SPME-HPLC) Analysis of Pesticides. Crit. Rev. Anal. Chem. 2005, 35, 71–85. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Panigrahi, S. Solid-Phase Microextraction (SPME) Techniques for Quality Characterization of Food Products: A Review. Food Bioprocess. Technol. 2011, 4, 1–26. [Google Scholar] [CrossRef]
- Murat, C.; Gourrat, K.; Jerosch, H.; Cayot, N. Analytical Comparison and Sensory Representativity of SAFE, SPME, and Purge and Trap Extracts of Volatile Compounds from Pea Flour. Food Chem. 2012, 135, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Engel, W.; Bahr, W.; Schieberle, P. Solvent Assisted Flavour Evaporation—A New and Versatile Technique for the Careful and Direct Isolation of Aroma Compounds from Complex Food Matrices. Eur. Food Res. Technol. 1999, 209, 237–241. [Google Scholar] [CrossRef]
- Burdock, G.A. Handbook of Flavor Ingredients, 6th ed.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2010; ISBN 978-1-4200-9077-2. [Google Scholar]
- Azarnia, S.; Boye, J.I.; Warkentin, T.; Malcolmson, L. Changes in Volatile Flavour Compounds in Field Pea Cultivars as Affected by Storage Conditions. Int. J. Food Sci. Technol. 2011, 46, 2408–2419. [Google Scholar] [CrossRef]
- Turco, I.; Ferretti, G.; Bacchetti, T. Review of the Health Benefits of Faba Bean (Vicia faba, L.) Polyphenols. J. Food Nutr. Res. 2016, 55, 11. [Google Scholar]
- Cabrera, A.; Martin, A. Genetics of Tannin Content and Its Relationship with Flower and Testa Colours in Vicia Faba. J. Agric. Sci. 1989, 113, 93–98. [Google Scholar] [CrossRef]
- Kaczmarska, K.T.; Chandra-Hioe, M.V.; Frank, D.; Arcot, J. Aroma Characteristics of Lupin and Soybean after Germination and Effect of Fermentation on Lupin Aroma. LWT 2018, 87, 225–233. [Google Scholar] [CrossRef]
- Czerny, M.; Christlbauer, M.; Christlbauer, M.; Fischer, A.; Granvogl, M.; Hammer, M.; Hartl, C.; Hernandez, N.M.; Schieberle, P. Re-Investigation on Odour Thresholds of Key Food Aroma Compounds and Development of an Aroma Language Based on Odour Qualities of Defined Aqueous Odorant Solutions. Eur. Food Res. Technol. 2008, 228, 265–273. [Google Scholar] [CrossRef]
- Shi, Y.; Mandal, R.; Singh, A.; Singh, A.P. Legume Lipoxygenase: Strategies for Application in Food Industry. Legume Sci. 2020, 2, e44. [Google Scholar] [CrossRef]
- Wang, J.; Kuang, H.; Zhang, Z.; Yang, Y.; Yan, L.; Zhang, M.; Song, S.; Guan, Y. Generation of Seed Lipoxygenase-Free Soybean Using CRISPR-Cas9. Crop. J. 2020, 8, 432–439. [Google Scholar] [CrossRef]
- Gürbüz, G.; Liu, C.; Jiang, Z.; Pulkkinen, M.; Piironen, V.; Sontag-Strohm, T.; Heinonen, M. Protein-Lipid Co-Oxidation in Emulsions Stabilized by Microwave-Treated and Conventional Thermal-Treated Faba Bean Proteins. Food Sci. Nutr. 2018, 6, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Pulkkinen, M.; Wang, Y.; Lampi, A.-M.; Stoddard, F.L.; Salovaara, H.; Piironen, V.; Sontag-Strohm, T. Faba Bean Flavour and Technological Property Improvement by Thermal Pre-Treatments. LWT Food Sci. Technol. 2016, 68, 295–305. [Google Scholar] [CrossRef]
- Schindler, S.; Wittig, M.; Zelena, K.; Krings, U.; Bez, J.; Eisner, P.; Berger, R.G. Lactic Fermentation to Improve the Aroma of Protein Extracts of Sweet Lupin (Lupinus angustifolius). Food Chem. 2011, 128, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Schindler, S.; Zelena, K.; Krings, U.; Bez, J.; Eisner, P.; Berger, R.G. Improvement of the Aroma of Pea (Pisum Sativum) Protein Extracts by Lactic Acid Fermentation. Food Biotechnol. 2012, 26, 58–74. [Google Scholar] [CrossRef]
- El Youssef, C.; Bonnarme, P.; Fraud, S.; Péron, A.-C.; Helinck, S.; Landaud, S. Sensory Improvement of a Pea Protein-Based Product Using Microbial Co-Cultures of Lactic Acid Bacteria and Yeasts. Foods 2020, 9, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas-Danguin, T.; Barba, C.; Salles, C.; Guichard, E. Perception of mixtures of odorants and tastants: Sensory and analytical points of view. In Flavour; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016; pp. 319–340. ISBN 978-1-118-92938-4. [Google Scholar]
- Thomas-Danguin, T.; Sinding, C.; Tournier, C.; Saint-Eve, A. 5—Multimodal interactions. In Flavor From Food to Behaviors, Wellbeing and Health; Etiévant, P., Guichard, E., Salles, C., Voilley, A., Eds.; Woodhead Publishing, Elsevier Ltd.: Duxford, UK, 2016; pp. 121–141. ISBN 978-0-08-100295-7. [Google Scholar]



{kind=link}
{kind=link}
| Amino Acids | Aldehydes | Alcohols | Acids |
|---|---|---|---|
| Alanine | Acetaldehyde | Ethanol * | Acetic acid * |
| Phenylalanine | Phenylacetaldehyde * | 2-Phenylethanol * | Phenylacetic acid |
| Phenylalanine | Benzaldehyde * | Benzyl alcohol * | Benzoic acid |
| Valine | 2-Methylpropanal * | 2-Methylpropanol * | 2-Methylpropanoic acid |
| Methionine | 3-Methylthiopropanal * | 3-Methylthiopropanol | 3-Methylthiopropanoic acid |
| Isoleucine | 2-Methylbutanal * | 2-Methylbutanol * | 2-Methylbutanoic acid * |
| Leucine | 3-Methylbutanal * | 3-Methylbutanol * | 3-Methylbutanoic acid * |
| Code | Pulses | Cultivar | Year | Location | Storage | Seed Transformation | Extraction | Separation and Identification | References | |
|---|---|---|---|---|---|---|---|---|---|---|
| Black bean | Black bean (Phaseolus vulgaris L.) | AC Harblack | 2005 | Morden, Canada | Dry room (23 °C, 15–20% RH) (whole). | Ground in flour (coffee mill) (whole). | HS-SPME: 10 g in a 125-mL Erlenmeyer flask capped, DVB/CAR/PDMS Stable Flex SPME fibre at 50 °C for 1 h. | GC-MS: desorption at 250 °C for 2 min, Supelcowax 10 polar column, started at 40/1/70 °C, then 70/5/200 °C and 200/50/250 °C. | [40] | |
| CDC Rio | ||||||||||
| Onyx | ||||||||||
| Pinto bean | Pinto bean (Phaseolus vulgaris L.) | AC Pintoba | ||||||||
| Maverick | ||||||||||
| Dark red kidney bean | Dark red kidney bean (Phaseolus vulgaris L.) | ROG 802 | ||||||||
| Redhawk | ||||||||||
| Whole pea | Pea (Pisum sativum L.) | Eclipse (Yellow field pea) | 2005 | Near Saskatoon, Canada | 4 °C (whole). | Ground in flour (whole). | HS-SPME: 3 g, CAR/PDMS SPME Fibre at 50 °C for 30 min. | GC-MS: desorption at 300 °C for 3 min, VF-5MS capillary column, started at 35/6/80 °C and 80/20/280 °C. | [11] | |
| 2006 | ||||||||||
| 2007 | ||||||||||
| Dehulled pea | - | Before 2013 | - | In a glass bottle, −18 °C. | Ground in flour (dehulled). | SAFE: 20 g in 100 mL of water, 2 h at 30 °C and 10–2 mbar. Liquid-liquid separation with 3 × 10 mL of CH2Cl2. Concentration using Kuderna Danish apparatus, 70 °C. | GC-MS: ZB1.MS non-polar column, injection of 2 µL, started at 50/4/160 °C, then 160/15/320 °C. | [42] | ||
| Pea protein | Protein isolate, Nutralys® (dehulled, wet process). | |||||||||
| - | Before 2020 | - | - | Protein concentrate (dehulled, dry process). | HS-SPME: 1.5 g was dissolved into saturated NaCl solution for 1 h at 20 °C, then transferred into a bottle and incubated at 50 °C in an ultrasonic bath, insertion of the DVB/CAR/PDMS Stable Flex SPME fibre at 50 °C for 20 min. | GC-MS: desorption at 250 °C for 3 min, DB-5MS column, started at 40/5/70 °C, then 70/10/200 °C and 200/50/250 °C. | [44] | |||
| Chickpea | Chickpea (Cicer arietinum) | Kabuli (Benying-1) | 2018 | Urumqi, China | −18 °C for a maximum of 3 weeks (whole). −20 °C for a maximum of 1 week (powder). | Dried using sunlight before storage (whole). Ground to a fine powder (80 mesh, mill). | HS-SPME: 1.5 g was dispersed in water and 5 mL was placed in a 20-mL headspace sampling vial and capped, PDMS/DVB fibre at 60 °C for 60 min. | GC-MS: desorption at 250 °C for 5 min, PEG 20 M column, started at 35/5/130 °C and 130/9/200 °C. | [43] | |
| Desi (YZ-364) | ||||||||||
| Faba bean Tannin | Faba bean (Vicia faba L. minor) | High tannin | 2016 | Alberta, Canada | In freezer bags (polypropylene), 22 °C and 18% RH in a dark and solvent-free room (flour). | Ground in flour (impact mill) (whole). | HS-SPME: 2 g was pre-incubated at 50 °C for 5 min, DVB/CAR/PDMS Stable Flex SPME fibre at 50 °C for 1 h. | GC-MS: desorption at 250 °C for 60 s, DB-17 mod polarity column, 40/5/200 °C. | [37] | |
| Low tannin | ||||||||||
| Faba bean Location | Low tannin (13 cultivars) | 2009 | Barrhead, Canada | - | Ground in flour (coffee mill) (whole). | HS-SPME: 10 g in a 125-mL Erlenmeyer flask capped, DVB/CAR/PDMS Stable Flex SPME fibre at 50 °C for 1 h. | GC-MS: desorption at 250 °C for 2 min, Supelcowax 10 polar column, started at 40/1/70 °C, then 70/5/200 °C and 200/50/250 °C. | [41] | ||
| Namao, Canada | ||||||||||
| Faba bean Storage | High tannin | 2016 | Alberta, Canada | No storage. | Ground in flour (micro-mill) with a water-cooled system to protect from overheating (whole). | HS-SPME: 2 g was pre-incubated at 50 °C for 5 min, DVB/CAR/PDMS Stable Flex SPME fibre at 50 °C for 1 h. | GC-MS: desorption at 250 °C for 60 s, DB-17 mod polarity column, 40/5/200 °C. | [45] | ||
| In bags (PE), in a dark and solvent-free room, 60 days (flour). | 22 °C, 19% RH. | |||||||||
| 4 °C, 9% RH. | ||||||||||
| −21 °C. | ||||||||||
| Aromatic Hydrocarbons | CAS | Origin (s) | Black Bean | Pinto Bean | Dark Red Kidney Bean | Pea | Chickpea | Faba Bean | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Tannin | Location | Storage | |||||||
| Toluene | 108-88-3 | FFA 1,2 | 0.00–0.86 | 0.96–1.86 | 0.00–0.73 | 1.20–2.40 | Coelution | 0.26–0.37 | 0.88–0.96 | 0.41–3.12 | |
| m-Ethyltoluene | 620-14-4 | 0.09–0.11 | 0.00–0.28 | ||||||||
| Benzene | 71-43-2 | FFA 4 | 0.00–0.50 | 0.09–0.11 | |||||||
| Ethylbenzene | 100-41-4 | FFA 1,2 | 0.00–0.45 | 0.00–0.44 | 0.30–0.80 | 0.12–0.17 | 1.19–1.28 | 0.19–0.73 | |||
| 1,2,3-Trimethylbenzene | 526-73-8 | FFA 2 | 0.00–0.54 | 0.54–1.03 | 0.12–0.13 | 0.00–0.21 | |||||
| 1,3,5-Trimethylbenzene | 108-67-8 | FFA 2 | 0.00–0.77 | 0.81–1.84 | 0.00–0.19 | ||||||
| Propylbenzene | 103-65-1 | 0.00–0.35 | 0.00–0.40 | 0.00–0.19 | |||||||
| Cumene | 98-82-8 | FFA 2 | 0.70–1.13 | 0.84–0.86 | 1.11–1.69 | 0.30–0.60 | 0.42–0.46 | ||||
| p-Xylene | 106-42-3 | FFA 1 | 1.15–1.50 | 0.00–1.20 | 0.00–1.17 | 0.40–1.00 | 0.27–0.38 | 0.00–0.70 | |||
| o-Xylene | 95-47-6 | FFA 1,2,4 | 0.95–1.16 | 1.14–1.73 | 1.13–1.45 | Coelution | |||||
| m-Xylene | 108-38-3 | FFA 1 | 1.25 | ||||||||
| 4-Ethyl-m-xylene | 874-41-9 | 0.05–0.07 | 0.00–0.64 | ||||||||
| p-Cymene | 99-87-6 | Trace | 0.32–0.33 | ||||||||
| Styrene | 100-42-5 | FFA 1,2,3; N 2,3 | 32.55–45.47 | 30.88–31.76 | 38.75–47.57 | 0.70–2.20 | 0.00–0.60 | 11.36–13.79 | 0.00–1.90 | ||
| α-Methylstyrene | 98-83-9 | 0.00–0.43 | 0.00–0.38 | 0.00–0.49 | |||||||
| Total | 38.51–49.28 | 38.24–38.51 | 43.41–50.72 | 3.50–6.70 | 1.25 | 0.00–0.60 | 1.07–1.27 | 14.15–16.89 | 0.68–7.75 | ||
| Aldehydes | CAS | Origin (s) | Black Bean | Pinto Bean | Dark Red Kidney Bean | Pea | Chickpea | Faba Bean | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Proteins | Tannin | Location | Storage | |||||||
| Benzaldehyde | 100-52-7 | FFA 2; AA 5,7,11 | 2.36–3.64 | 2.58–2.60 | 3.46–4.04 | 0.00–2.11 | 0.00–2.19 | 0.36–0.45 | 2.09–2.10 | 0.46–0.85 | ||
| 4-Ethylbenzaldehyde | 53951-50-1 | 0.00–0.54 | ||||||||||
| Phenylacetaldehyde | 122-78-1 | AA 3,5,7,11 | 0.00–0.67 | 0.14–0.17 | 0.21–0.39 | |||||||
| Vanillin | 121-33-5 | 0.16 | 0.00–0.08 | |||||||||
| 2-Methylpropanal | 78-84-2 | AA 11 | 0.49–0.51 | |||||||||
| 3-Methylthiopropanal | 3268-49-3 | AA 3,11 | 0.00–0.03 | |||||||||
| 2-Methylbutanal | 96-17-3 | AA 5,8,11 | 0.03–0.49 | 0.31–0.48 | 0.00–0.13 | |||||||
| 3-Methylbutanal | 590-86-3 | AA 5,7,11 | 0.00–1.00 | 0.00–0.10 | 1.04–1.17 | 0.36–0.59 | ||||||
| Pentanal | 110-62-3 | FFA 2,9,10 | 0.00–0.59 | 0.60–0.79 | 1.03–1.11 | 0.00–0.73 | 0.96–1.21 | |||||
| Furfural | 98-01-1 | 0.00–0.05 | ||||||||||
| Hexanal | 66-25-1 | FFA 1,2,3,4,6,7,8,9,10 | 12.76–16.71 | 9.77–11.27 | 15.88–18.6 | 1.50–6.10 | 0.93 | 27.22–54.12 | 10.28–13.85 | 40.78–40.88 | 1.29–25.07 | |
| 2-Ethylhexanal | 123-05-7 | 0.00–0.16 | ||||||||||
| (E)-2-Hexenal | 6728-26-3 | FFA 1,2,4,8,9,10 | 0.00–1.60 | 0.00–1.67 | 0.00–1.75 | 0.00–0.19 | ||||||
| Heptanal | 111-71-7 | FFA 2,3,7,8,9,10 | 0.00–0.75 | 0.00–0.82 | 0.00–0.95 | 1.13–5.31 | 1.04–1.14 | 1.10–1.14 | 0.00–1.91 | |||
| (E)-2-Heptenal | 18829-55-5 | FFA 1,2,4,8,9,10 | 1.52–1.75 | 1.54–2.01 | 1.52–1.88 | 0.00–2.60 | 0.00–0.58 | 1.79–1.97 | ||||
| (E,E)-2,4-Heptadienal | 4313-03-5 | FFA 2,8,9,10 | 0.57–0.66 | 0.57–0.59 | 0.00–0.71 | Trace | 0.00−Coelution | |||||
| Octanal | 124-13-0 | FFA 2,7,8,9,10 | 0.00–0.62 | 0.00–0.68 | 0.00–0.58 | 1.35–1.76 | 2.07–2.40 | 1.15–1.29 | 0.08–2.22 | |||
| (E)-2-Octenal | 2548-87-0 | FFA 1,3,7,8,9,10 | 3.00–13.10 | 0.00–0.10 | 2.13–2.16 | 0.23–0.23 | 0.48–0.51 | 0.19–0.45 | ||||
| Nonanal | 124-19-6 | FFA 2,4,6,7,8,9,10 | 2.11–3.08 | 2.42–2.80 | 2.13–3.60 | 1.30 | 3.12–5.24 | 3.70–4.60 | 9.8–12.31 | 7.99–10.54 | 3.54–36.29 | |
| (E)-2-Nonenal | 18829-56-6 | FFA 2,7,9,10 | 0.00–0.67 | 0.00–0.58 | 0.20–0.26 | 0.14–2.52 | ||||||
| 4-Oxononanal | 74327-29-0 | 0.72–0.91 | ||||||||||
| (E,E)-2,4-Nonadienal | 5910-87-2 | FFA 7,9,10 | 0.00–0.44 | |||||||||
| Decanal | 112-31-2 | FFA 2,7,10 | 0.81–1.59 | 0.00–1.06 | 0.72–0.99 | Coelution | Coelution−1.33 | 1.28–1.33 | 2.86–3.26 | 1.15–1.41 | 0.37–6.16 | |
| (Z)-2-Decenal | 2497-25-8 | FFA 7,10 | 0.79–1.15 | |||||||||
| (E,E)-2,4-Decadienal | 25152-84-5 | FFA 3,7,9,10 | Trace | |||||||||
| Undecanal | 112-44-7 | 0.22–0.26 | 0.00–1.61 | |||||||||
| Dodecanal | 112-54-9 | 0.00–0.89 | ||||||||||
| Tetradecanal | 124-25-4 | 0.63 | 0.26–0.55 | 0.00–0.39 | ||||||||
| Total | 24.42–28.01 | 18.19–23.57 | 25.77–33.33 | 10.70–17.70 | 3.02 | 40.21–63.66 | 12.41–12.91 | 29.14–36.39 | 58.05–60.58 | 9.19–79.03 | ||
| Alkanes/Alkenes | CAS | Origin (s) | Black Bean | Pinto Bean | Dark Red Kidney Bean | Pea | Chickpea | Faba Bean | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Proteins | Tannin | Location | Storage | |||||||
| Trichloromethane | 67-66-3 | N 1 | 0.00–0.50 | |||||||||
| Octylcyclopropane | 1472-09-9 | 0.00–0.96 | 0.00–1.28 | 0.00–2.04 | ||||||||
| Pentane | 109-66-0 | FFA 4 | 0.10–0.10 | |||||||||
| Hexane | 110-54-3 | FFA 4 | 0.74–1.54 | 1.14–1.72 | 0.83–0.86 | |||||||
| 3-Methylhexane | 589-34-4 | 0.00–2.50 | ||||||||||
| Butylcyclohexane | 1678-93-9 | 0.00–0.49 | 0.00–0.51 | |||||||||
| Heptane | 142-82-5 | FFA 3 | 0.54–1.15 | 0.94–1.07 | 0.57–0.76 | 0.55–0.56 | ||||||
| Octane | 111-65-9 | FFA 4 | 1.74–2.66 | 0.00–1.74 | 1.89–3.64 | 1.79–1.96 | ||||||
| 2,6-Dimethyloctane | 2051-30-1 | 0.00–0.48 | 0.46–0.83 | |||||||||
| Nonane | 111-84-2 | FFA 4 | 0.75–0.95 | 1.00–1.37 | 0.77–1.17 | 0.16–0.17 | 0.34–0.36 | |||||
| 3-Methylnonane | 5911-04-6 | 0.00–0.52 | 0.56–0.80 | |||||||||
| 4-Methylnonane | 17301-94-9 | 0.00–0.94 | 0.95–1.64 | |||||||||
| 3,7-Dimethylnonane | 17302-32-8 | 0.18–0.19 | ||||||||||
| Decane | 124-18-5 | FFA 2,4 | 2.66–4.27 | 4.78–6.55 | 1.60–1.86 | 0.67–0.70 | 0.95–3.88 | |||||
| 4-Methyldecane | 2847-72-5 | 0.00–0.75 | 0.64–1.34 | |||||||||
| 2,4-Dimethyldecane | 2801-84-5 | 0.00–0.32 | ||||||||||
| 3,7-Dimethyl-decane | 17312-54-8 | 0.00–0.31 | ||||||||||
| Undecane | 1120-21-4 | FFA 1,4 | 0.00–0.87 | 1.06–1.51 | 1.80–2.60 | 0.15–0.15 | 1.36–1.52 | |||||
| Dodecane | 112-40-3 | 0.00–1.12 | 1.22–1.27 | 0.80–3.60 | 0.57–0.62 | 0.80–3.22 | ||||||
| 2,4-Dimethyldodecane | 6117–99–3 | FFA 1 | 0.09–0.09 | 0.00–0.33 | ||||||||
| 5,8-Diethyldodecane | 24251-86-3 | 0.04–0.06 | ||||||||||
| 2,6,10-Trimethyldodecane | 3891-98-3 | 0.00–0.79 | 0.64–1.27 | |||||||||
| Tridecane | 629-50-5 | 0.90–4.30 | 0.06–0.08 | 0.00–0.15 | ||||||||
| 2-Methyltridecane | 1560-96-9 | FFA 1 | 0.05–0.05 | 0.00–0.14 | ||||||||
| 3-Methyltridecane | 6418-41-3 | FFA 1 | 0.12–0.15 | 0.00–0.42 | ||||||||
| 2,2-Dimethyltridecane | 61869-04-3 | 0.13–0.15 | ||||||||||
| Tetradecane | 629-59-4 | 0.00–1.10 | Trace | 0.00–1.10 | 1.30–1.90 | 0.48–1.34 | ||||||
| Pentadecane | 629-62-9 | 0.00–1.57 | 0.19–0.21 | 0.00–0.64 | ||||||||
| 3-Methylpentadecane | 2882-96-4 | 0.15–0.26 | 0.00–0.08 | |||||||||
| Hexadecane | 544-76-3 | 0.00–2.75 | 0.24–0.26 | 0.00–0.35 | ||||||||
| Heptadecane | 629-78-7 | 1.27 | 0.07–0.10 | |||||||||
| Nonadecane | 629-92-5 | 0.02–0.06 | 0.46–0.99 | |||||||||
| Tetracosane | 646-31-1 | 0.00–0.57 | ||||||||||
| Total alkanes | 7.84–15.66 | 16.43–20.00 | 7.95–8.09 | 7.30–14.30 | 1.27 | 0.00–5.44 | 0.31–0.57 | 4.37–5.22 | 4.06–4.41 | 4.61–9.22 | ||
| (E)-5-(Pentyloxy)-2-pentene | 34061-80-8 | 0.00–0.69 | ||||||||||
| (Z)-1-Methoxy-3-hexene | 70220-06-3 | 0.00–1.91 | ||||||||||
| 3-Ethyl-2-methyl-1,3-hexadiene | 61142-36-7 | 0.00–0.05 | 0.37–0.42 | |||||||||
| 1-Tetradecene | 1120-36-1 | 5.74 | 0.00−Coelution | 0.07–0.08 | ||||||||
| Total alkenes | 5.74 | 0.05–0.69 | 0.00–1.91 | 0.07–0.08 | 0.37–0.42 | |||||||
| Alcohols | CAS | Origin (s) | Black Bean | Pinto Bean | Dark Red Kidney Bean | Pea | Chickpea | Faba Bean | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Proteins | Tannin | Location | Storage | |||||||
| Ethanol | 64-17-5 | AA 5 FFA 9,10,11 | 0.48–1.29 | 0.00–0.42 | ||||||||
| 2-Phenylethanol | 60-12-8 | AA 5,7,11 | 0.27–0.34 | 0.00–0.83 | ||||||||
| 2-Butoxyethanol | 111-76-2 | 0.95 | 0.00–0.88 | |||||||||
| 2-Phenoxyethanol | 122-99-6 | Coelution | 0.00−Coelution | 0.00–0.30 | ||||||||
| Propanol | 71-23-8 | AA 1 FFA 9,10 | 0.60–1.30 | 0.48–0.53 | 0.00–0.64 | |||||||
| 2-Propanol | 67-63-0 | 0.48–0.57 | 0.00–0.15 | |||||||||
| 1,2-Propanediol | 57-55-6 | 0.00–0.04 | ||||||||||
| 2-Methylpropanol | 78-83-1 | AA 9,11 | 0.00–0.50 | |||||||||
| 1-Methoxy-2-Propanol | 107-98-2 | 0.00–0.28 | ||||||||||
| 2-Phenyl-2-propanol | 617-94-7 | 0.38–0.41 | ||||||||||
| 1-[1-Methyl-2-(2-propenyloxy)ethoxy]-2-propanol | 55956-25-7 | 0.00–0.09 | ||||||||||
| Butanol | 71-36-3 | FFA 9,10 | 4.32–4.72 | |||||||||
| 2-Butanol | 78-92-2 | 1.29–1.38 | 0.00–1.90 | |||||||||
| 2-Methylbutanol | 137-32-6 | AA 5,8,9,11 | 0.06–0.34 | 3.52–4.80 | 0.38–5.48 | |||||||
| 3-Methylbutanol | 123-51-3 | AA 5,7,8,9,11 | 0.00–0.80 | 0.58–0.86 | 0.42 | 0.06–0.22 | 1.97–2.43 | 0.12–3.23 | ||||
| 3-Phenyl-2-butanol | 52089-32-4 | 0.00–1.23 | ||||||||||
| 2,3-Butanediol | 513-85-9 | 0.39–0.40 | 0.00–1.72 | |||||||||
| Pentanol | 71-41-0 | FFA 2,3,4,7,8,9,10 | 0.00–1.61 | 0.00–0.62 | 1.52 | 1.17–3.89 | 4.90–4.98 | 1.28–1.36 | 1.85–2.29 | 0.74–1.57 | ||
| 2-Pentanol | 6032-29-7 | FFA 9 | 0.23 | 0.18–0.27 | 0.00–0.46 | |||||||
| 3-Methyl-3-pentanol | 77-74-7 | 5.34 | 0.00–0.60 | |||||||||
| 1-Penten-3-ol | 616-25-1 | FFA 2,3,4,8,9,10 | 0.99–1.86 | 1.34–1.88 | 2.30–2.64 | 6.87 | 0.44–5.58 | |||||
| 2-Penten-1-ol | 20273-24-9 | 0.00–3.84 | ||||||||||
| 5-[3-(4-Methoxyphenyl)-2-oxaziridinyl]-1-pentanol | - | 0.33–0.90 | ||||||||||
| Phenol | 108–95–2 | 0.00–0.47 | 0.31–0.34 | 0.00−Coelution | 0.54–0.70 | |||||||
| Benzyl alcohol | 100-51-6 | AA 5,11 | 0.00–0.24 | 0.26–1.11 | 0.00−Coelution | 0.54–0.54 | 0.58–1.46 | |||||
| Hexanol | 111-27-3 | FFA 2,3,4,6,7,8,9,10 | 1.58–1.86 | 1.39–1.60 | 1.24–1.25 | 4.54 | 0.88–9.53 | 10.64–11.32 | 3.87–4.29 | 0.33–31.41 | ||
| 2-Ethylhexanol | 104-76-7 | 0.29–0.45 | 0.29–0.66 | 0.00–0.4 | 0.00–1.39 | 0.00–9.38 | ||||||
| 4-Ethylcyclohexanol | 4534-74-1 | 0.15–0.17 | ||||||||||
| 2,3-Dimethylcyclohexanol | 1502-24-5 | 0.00–0.20 | ||||||||||
| 1-Hexen-3-ol | 4798-44-1 | 0.29 | 0.00–0.14 | |||||||||
| (Z)-3-Hexen-1-ol | 928-96-1 | FFA 8,9 | 4.23 | 0.00–0.29 | ||||||||
| (Z)-4-Hexen-1-ol | 928-91-6 | 0.00–0.33 | ||||||||||
| Heptanol | 111-70-6 | FFA 8,9,10 | 0.00–1.20 | Coelution | Coelution−0.66 | 0.32–0.36 | 0.50–0.54 | 0.00–0.26 | ||||
| 2-Heptanol | 543-49-7 | FFA 9 | 0.00–0.40 | |||||||||
| 2-Methyl-3-heptanol | 18720-62-2 | 0.72 | ||||||||||
| 3-Methyl-2-heptanol | 31367-46-1 | 0.00–2.40 | ||||||||||
| 2-Hepten-4-ol | 4798-59-8 | 0.00–0.78 | ||||||||||
| Octanol | 111-87-5 | FFA 3,7,8,9,10 | 0.00–0.33 | 0.29–0.61 | 0.00–0.29 | 3.7–10.7 | 1.16 | 0.82–1.29 | 0.24–0.28 | 1.18–1.48 | 0.35–0.83 | |
| 3-Octanol | 589-98-0 | FFA1 | 0.00–1.3 | 0.00–0.42 | ||||||||
| 1-Octen-3-ol | 3391-86-4 | FFA 2,3,4,6,7,8,9,10 | 1.18–1.43 | 1.23–1.60 | 0.00–3.51 | 2.61 | 1.23–1.38 | 0.35–0.41 | 3.93–4.00 | 0.29–1.64 | ||
| (E)-2-Octen-1-ol | 18409-17-1 | FFA 10 | 0.00–0.96 | 0.37–0.41 | ||||||||
| (E,E)-3,5-Octadien-2-ol | 69668-82-2 | 0.10–0.14 | ||||||||||
| Nonanol | 143-08-8 | FFA 7 | 0.42–1.30 | 0.92–1.10 | 0.00–0.43 | 1.27 | 0.00–2.69 | 0.92–1.14 | 0.28–0.41 | 0.85–1.74 | ||
| Decanol | 112-30-1 | 0.22–0.29 | ||||||||||
| Undecanol | 112-42-5 | 6.18 | 0.00–0.09 | |||||||||
| 2-Pentadecyn-1-ol | 2834-00-6 | 0.00–0.55 | ||||||||||
| Total | 6.13–7.72 | 8.05–9.04 | 4.67–7.41 | 7.40–12.70 | 36.33 | 8.22–30.70 | 11.22–13.01 | 24.21–27.63 | 13.00–13.27 | 4.34–53.01 | ||
| Ketones | CAS | Origin (s) | Black Bean | Pinto Bean | Dark Red Kidney Bean | Pea | Chickpea | Faba Bean | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Proteins | Tannin | Location | Storage | |||||||
| Acetophenone | 98-86-2 | AA 5; FFA 6 | 0.63–0.98 | 0.53–0.77 | 0.00–0.82 | 0.33–0.36 | 0.00–0.50 | |||||
| p-Isopropylacetophenone | 645-13–6 | 0.00–0.73 | 0.15–0.57 | |||||||||
| p-Acetylacetophenone | 1009-61-6 | 0.19–0.29 | ||||||||||
| Acetone | 67-64-1 | FFA 5 | 0.66–1.47 | 0.85–1.12 | 1.16–1.75 | 1.99–2.46 | 1.00–1.02 | 0.41–0.92 | ||||
| Butanone | 78-93-3 | FFA 1,2,6 | 0.00–0.41 | 0.00–0.38 | 0.00–0.50 | 0.00–0.97 | 3.20–3.43 | 0.58–0.67 | 0.79–4.82 | |||
| 3-Hydroxy-3-methyl-2-butanone | 115-22-0 | 0.00−Coelution | ||||||||||
| Pentanone | 107-87-9 | FFA 1,6 | 0.00–1.20 | |||||||||
| 3-Pentanone | 96-22-0 | 0.87 | ||||||||||
| 2,3-Pentanedione | 600-14-6 | FFA 6 | 0.42 | 0.00–1.05 | ||||||||
| Hexanone | 591-78-6 | FFA 6 | 0.03–0.04 | |||||||||
| Cyclohexanone | 108-94-1 | 0.00–0.15 | ||||||||||
| 5-Hexen-2-one | 109-49-9 | 3.29 | 0.00–1.75 | |||||||||
| 6-Methyl-5-hepten-2-one | 110-93-0 | FFA 2; CAR 4 | 1.97–2.45 | 0.94–1.76 | 1.27–2.44 | 0.82–0.87 | 1.08–1.50 | 1.06–1.09 | 0.00–1.27 | |||
| Heptanone | 110-43-0 | FFA 3,6 | 0.17 | 0.25–1.03 | 0.27–0.32 | 0.46–0.48 | 0.24–0.35 | |||||
| 2-Methyl-3-heptanone | 13019-20-0 | 6.85 | 0.00–6.50 | |||||||||
| Isobutyl-2-heptenone | - | 0.00–0.17 | ||||||||||
| Octanone | 111-13-7 | FFA 6 | 0.41 | |||||||||
| 3-Octanone | 106-68-3 | FFA 4,6 | Trace | |||||||||
| 3-Octen-2-one | 1669-44-9 | FFA 5 | 0.28–0.83 | 0.69–0.86 | ||||||||
| 2,3-Octanedione | 585-25-1 | Trace | 0.00–1.34 | 1.71–2.00 | 0.52–0.53 | |||||||
| (E,E)-3,5-Octadien-2-one | 30086-02-3 | FFA 2,5 | 1.14–1.98 | 1.31–2.41 | 1.42–1.46 | 0.00–8.00 | 0.20–0.28 | 0.16–0.20 | ||||
| Nonanone | 821-55-6 | 0.26 | ||||||||||
| Decanone | 693-54-9 | 0.00–0.35 | ||||||||||
| 1,6-Dioxacyclododecane-7,12-dione | 777-95-7 | 0.00–0.72 | ||||||||||
| Undecanone | 112-12-9 | Trace | 0.00–0.18 | |||||||||
| 2-Butyl-1,3,2-dioxaborinan-4-one | 33823-94-8 | 0.00–0.37 | ||||||||||
| Total | 4.95–6.90 | 4.74–5.38 | 4.73–5.62 | 0.50–1.20 | 12.01 | 1.69–21.96 | 3.24–3.25 | 7.59–8.42 | 4.48–4.53 | 3.72–7.46 | ||
| Acids | CAS | Origin (s) | Pea | Chickpea | Faba Bean | ||
|---|---|---|---|---|---|---|---|
| Dehulled | Proteins | Tannin | Storage | ||||
| Acetic acid | 64-19-7 | AA 2,3,4 | 3.10–3.90 | 1.84–2.35 | 0.49–1.94 | ||
| 2-Methylbutanoic acid | 116-53-0 | N 1; AA 1,2,4 | 0.00–0.22 | 0.13 | 0.00–0.26 | ||
| 3-Methylbutanoic acid | 503-74-2 | AA 2,3,4 | 0.42 | 0.00–0.41 | 8.79–12.37 | 2.55–10.36 | |
| Pentanoic acid | 109-52-4 | 0.48 | |||||
| Hexanoic acid | 142-62-1 | FFA 1 | 0.73 | 0.00−Coelution | 1.06–1.98 | 0.57–1.54 | |
| 2-Ethyl hexanoic acid | 149-57-5 | 0.51 | 0.00–0.43 | ||||
| Heptanoic acid | 111-14-8 | 1.46 | |||||
| Octanoic acid | 124-07-2 | N 1; FFA 3 | 1.54 | 0.00−Coelution | 0.00–0.24 | ||
| Nonanoic acid | 112-05-0 | FFA 3 | 1.92 | 0.00–1.40 | 0.00–0.58 | ||
| Decanoic acid | 334-48-5 | 0.00–0.89 | |||||
| Palmitic acid | 57-10-3 | 12.00–15.00 | |||||
| Oleic acid | 112-80-1 | 11.00–24.00 | |||||
| Total | 7.07 | 0.63–1.84 | 32.77–40.16 | 11.33–16.39 | 3.24–13.15 | ||
| Esters | CAS | Origin (s) | Black bean | Pinto Bean | Dark Red Kidney Bean | Pea | Chickpea | Faba Bean | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Proteins | Tannin | Storage | |||||||
| Ethyl ethanoate | 141-78-6 | FFA 3 | 0.00–1.20 | ||||||||
| 2-Ethylhexyl ethanoate | 103-09-3 | 0.00–0.79 | |||||||||
| Butyl ethanoate | 123-86-4 | 0.00–0.25 | |||||||||
| Hexyl ethanoate | 142-92-7 | FFA 2 | 0.00–2.96 | ||||||||
| Ethyl cyanoacetate | 105-56-6 | 0.00–0.39 | |||||||||
| Decyl bromoacetate | 5436-93-1 | 0.00–0.36 | 0.00–0.32 | 0.00–0.34 | |||||||
| 1,2-Dihydro-2-naphtalenylacetate | - | 0.80–1.06 | |||||||||
| Ethyl propanoate | 105-37-3 | Coelution | 0.00–17.19 | ||||||||
| Octyl pivalate | - | 0.00–0.76 | |||||||||
| Hexyl 2-methylbutanoate | 10032-15-2 | 0.00–3.54 | |||||||||
| Ethyl butanoate | 105-54-4 | 0.93 | 0.00–0.05 | 0.00–3.66 | |||||||
| 2-Ethylhexyl butanoate | 25415-84-3 | 0.00–0.92 | |||||||||
| Methyl isovalerate | 556-24-1 | AA 1 | 0.00–2.11 | ||||||||
| Isoamyl isovalerate | 659-70-1 | 0.00–0.51 | |||||||||
| Butyl hexanoate | 626-82-4 | 0.00–7.48 | |||||||||
| 2-Propenyl hexanoate | 123-68-2 | 0.82 | |||||||||
| Hexyl hexanoate | 6378-65-0 | 0.00–3.33 | |||||||||
| Octyl hexanoate | 4887-30-3 | 0.00–1.75 | |||||||||
| Methyl salicylate | 119-36-8 | 0.04–0.05 | 0.00–0.50 | ||||||||
| 5-Isobutylnonane | 62185-53-9 | 0.00–3.10 | |||||||||
| 4-Dodecanoyloxybutyl dodecanoate | 624-07-7 | 0.00–0.77 | |||||||||
| Isopropyl myristate | 110-27-0 | 0.44–0.53 | |||||||||
| 3-Hydroxy-ethyl mandelate | - | 0.28–0.42 | 0.34–0.45 | 0.00–0.42 | |||||||
| Total | 0.42–0.66 | 0.45–0.66 | 0.00–0.77 | 0.00–1.20 | 1.75 | 1.75–17.25 | 1.50–5.59 | 0.04–0.05 | 0.00–24.15 | ||
| Pyrazines | CAS | Origin (s) | Pea | ||
|---|---|---|---|---|---|
| Whole | Dehulled | Protein | |||
| 2,3-Diethyl-5-methylpyrazine | 18138-04-0 | 0.00–1.30 | |||
| 2-Methyoxy-3-isopropyl(5or6)-methylpyrazine | 32021-41-3 | AA 1 | 2.62 | 0.00–0.13 | |
| 2-Methoxy-3-isobutylpyrazine | 24683-00-9 | AA 2 | Trace | ||
| Total | 0.00–1.30 | 2.62 | 0.00–0.13 | ||
| Terpenes | CAS | Origin (s) | Black Bean | Pinto Bean | Dark Red Kidney Bean | Pea | Faba Bean | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Proteins | Tannin | Storage | ||||||
| α-Pinene | 80-56-8 | CAR 1,2 | 1.37–2.29 | 2.11–3.92 | 0.75–1.07 | |||||
| Δ3-Carene | 13466-78-9 | CAR 1,2 | 0.00–0.50 | 0.00–0.48 | 0.30–0.70 | |||||
| Limonene | 138-86-3 | CAR 1,2 | 0.00–1.36 | 1.31–2.96 | 0.66 | 0.00–0.11 | 0.96 | 0.00–10.81 | ||
| γ-Terpinene | 99-85-4 | 0.00–0.79 | ||||||||
| Terpinolene | 586-62-9 | 0.00–0.12 | ||||||||
| Thujospsene-I3 | - | 0.00–0.09 | ||||||||
| (E)-β-Ionone | 79-77-6 | CAR 3 | 0.11–0.12 | |||||||
| β-Myrcene | 123-35-3 | CAR 2 | 0.00–0.60 | |||||||
| Geranylacetone | 689-67-8 | 0.00–0.69 | 0.00–0.30 | |||||||
| Elemol | 639-99-6 | 2.71 | ||||||||
| α-Muurolol | 19435-97-3 | 2.94 | ||||||||
| t-Muurolol | 19912-62-0 | 7.45 | ||||||||
| α-Cadinol | 481-34-5 | Coelution | ||||||||
| β-Eudesmol | 473-15-4 | Trace | ||||||||
| β-Linalool | 78-70-6 | 0.11–0.79 | ||||||||
| Menthol | 1490-04-6 | 0.28–0.45 | 0.00–0.60 | |||||||
| p-Menth-1,5-dien-8-ol | 1686-20-0 | Coelution | ||||||||
| Total | 2.06–4.77 | 3.91–6.89 | 1.06–1.07 | 0.30–0.70 | 13.77 | 0.00–0.11 | 1.35–1.63 | 0.11–12.73 | ||
| Furans | CAS | Origin (s) | Black Bean | Pinto Bean | Dark Red Kidney Bean | Pea | Faba Bean | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Proteins | Tannin | Location | Storage | ||||||
| 2-Methylfuran | 534-22-5 | FFA 3 | 0.00–1.80 | ||||||||
| 2-Ethylfuran | 3208-16-0 | FFA 1,5 | 0.00–0.64 | 0.62–0.74 | 0.00–0.58 | 0.00–3.90 | 0.29–0.29 | ||||
| 2-Acetylfuran | 1192-62-7 | Coelution | 0.00–Trace | ||||||||
| 2-Propylfuran | 4229-91-8 | 0.53–0.71 | 0.49–0.61 | 0.75–0.85 | |||||||
| 2-Propionylfuran | 3194-15-8 | 0.44–0.66 | 0.00–0.78 | 0.53–0.69 | |||||||
| 2-Pentylfuran | 3777-69-3 | FFA 2,3,4,5 | 0.00–0.91 | 0.60–0.78 | 1.72–1.91 | 0.61–1.43 | |||||
| Total | 1.37–1.64 | 1.23–2.02 | 1.43–1.97 | 0.00–5.70 | 0.00–0.91 | 0.60–0.78 | 2.02–2.21 | 0.61–1.43 | |||
| Lactones | CAS | Origin (s) | Pea | Chickpea | Faba Bean | ||
|---|---|---|---|---|---|---|---|
| Dehulled | Proteins | Tannin | Storage | ||||
| γ-Butyrolactone | 96-48-0 | 0.00–0.85 | 0.32–0.63 | 0.22–1.08 | |||
| 3-Methylbutyrolactone | 1676-49-8 | 0.45 | |||||
| 4-Methyl-4-vinylbutyrolactone | 1073-11-6 | 0.08–0.09 | |||||
| Pentolactone | 599-04-2 | 2.20–2.30 | |||||
| γ-Pentalactone | 108-29-2 | 1.01 | |||||
| δ-Caprolactone | 823-22-3 | N 2 | 0.24–0.26 | 0.00–0.82 | |||
| γ-Caprolactone | 695-06-7 | N 2 | 10.11 | 0.00–Coelution | 0.34–0.36 | 0.00–1.07 | |
| γ-Methyl-γ-caprolactone | 2865-82-9 | 1.16 | 0.00–1.63 | ||||
| 4-Hydroxy-2-hexenoic acid lactone | 2407-43-4 | 0.00–0.17 | |||||
| γ-Octalactone | 104-50-7 | FFA 1 | Trace | 0.00-Trace | |||
| γ-Nonalactone | 104-61-0 | FFA 1 | 1.31 | 0.00–0.33 | |||
| 4-Hydroxy-2-noneic acid lactone | 21963-26-8 | FFA 1 | 0.72 | 0.00–1.04 | |||
| δ-Undecalactone | 710-04-3 | N 2 | 0.00–0.47 | 0.19–0.20 | |||
| Total | 14.76 | 0.00–3.18 | 2.30–3.52 | 1.21–1.50 | 0.22–2.47 | ||
| Other Volatiles | CAS | Black Bean | Pinto Bean | Dark Red Kidney Bean | Pea | Chickpea | Faba Bean | |||
|---|---|---|---|---|---|---|---|---|---|---|
| Whole | Dehulled | Proteins | Tannin | Storage | ||||||
| Estragole | 140-67-0 | 0.00–0.16 | ||||||||
| Benzothiazole | 95-16-9 | 0.42 | 0.00–1.50 | 0.00–0.30 | ||||||
| 4,5-Dimethylimidazole | 2302-39-8 | 0.00–1.22 | ||||||||
| Dimethyl sulphide | 75-18-3 | 0.86–3.30 | ||||||||
| Dimethyl disulphide | 624-92-0 | 0.00–0.80 | ||||||||
| 3-Phenylindole | 1504-16-1 | 0.00–0.84 | 0.49–1.18 | 0.72–0.73 | ||||||
| Methoxy-phenyl-oxime | - | 0.43–0.88 | 0.00–0.62 | 0.00–0.72 | 3.39–3.60 | |||||
| 2,4-Dimethylbenzenamine | 95-68-1 | 0.00–0.26 | 0.00–0.25 | 0.00–0.35 | ||||||
| 2-(Trimethylsilylethynyl)pyridine | 86521-05-3 | 0.00–0.40 | ||||||||
| Total | 0.72–1.72 | 1.18–1.37 | 0.72–1.80 | 1.26–3.40 | 0.42 | 0.00–1.50 | 0.00–1.92 | 3.39–3.60 | 0.00–0.16 | |
| Chemical Classes | Volatiles | CAS | Origin (s) | Odour Threshold | Odour Descriptors | Pea | Lupin | ||
|---|---|---|---|---|---|---|---|---|---|
| Dehulled 1 (DM) | Protein Isolate 1 (DM) | Whole 2 | Dehulled 3 | ||||||
| Aldehydes | 4-Ethylbenzaldehyde | 53951-50-1 | 13 ppb 5 | Sweet, honey, hay, dry vegetable (pea isolate) | (ND) | %D 83 (1.7 µg/g) | |||
| Phenylacetaldehyde | 122-78-1 | AA | 4 ppb 5 | Floral (pea isolate) | (ND) | %D 56 (2.1 µg/g) | |||
| Vanillin | 121-33-5 | N 6 | 53 µg/L (water) 4 | Sweet, vanilla, caramel (peas, dehulled lupin) | %D 33 (0.1 µg/g) | %D 28 (0.3 µg/g) | FD 1024 | ||
| Ethylvanillin | 121-32-4 | N 6 | 100 ppb 5 | Vanilla (dehulled lupin) | FD 256 | ||||
| 3-Methylthiopropanal | 3268-49-3 | AA | 0.1 % (oil) 5 | Potato (pea isolate) | (ND) | %D 94 (0.1 µg/g) | |||
| Hexanal | 66-25-1 | FFA | 2.4 µg/L (water) 4 | Grass, floral, fruity (dehulled pea)/Fresh grass (pea isolate)/Fresh green bean, green (whole lupin) | %D 50 (0.6 µg/g) | %D 61 (83.0 µg/g) | OI 10 (135 µg/g) | ||
| Heptanal | 111-87-7 | FFA | Green vegetable (pea isolate) | (ND) | %D 28 (16.2 µg/g) | (NO–4 µg/g) | |||
| Octanal | 124-13-0 | FFA | 3.4 µg/L (water) 4 | Floral, sweet, beany (whole lupin) | OI 26 (3 µg/g) | ||||
| (E)-2-Octenal | 2548-87-0 | FFA | 3–6 ppb 5 | Pea, dry vegetable (pea isolate)/Fatty, green, cucumber (dehulled lupin) | (ND) | %D 94 (0.3 µg/g) | OI 98 (3 µg/g) | ||
| Nonanal | 124-19-6 | FFA | 2.8 µg/L (water) 4 | Solvent (dehulled pea) | %D 17 (0.9 µg/g) | (NO–9.5 µg/g) | |||
| (E)-2-Nonenal | 18829-56-6 | FFA | 0.19 µg/L (water) 4 | Beany, green (whole lupin)/Cardboard, fatty, green (dehulled lupin) | OI 49 (6 µg/g) | FD 256 | |||
| (Z)-2-Nonenal | 60784-31-8 | FFA 3 | 0.1 ppb 5 | Cardboard (dehulled lupin) | FD 32 | ||||
| (E,Z)-2,6-Nonadienal | 557-48-2 | FFA 3 | 45 µg/L (water) 4 | Sweet, fatty (whole lupin)/Cucumber, green (dehulled lupin) | OI 10 (7 µg/g) | FD 256 | |||
| (E,E,Z)-2,4,6-Nonatrienal | 100113-52-8 | FFA 3 | Nutty, oat-flake (dehulled lupin) | FD 256 | |||||
| (E)-4,5-Epoxy-(E)-decenal | 134454-31-2 | FFA 3 | 0.19 µg/L (water) 4 | Metallic (dehulled lupin) | FD 1024 | ||||
| (E,E)-2,4-Decadienal | 25152-84-5 | FFA | 0.038 µg/L (water) 4 | Peanut, grilled meat (peas) | %D 94 (Trace) | %D 83 (Coelution) | |||
| Alcohols | Pentanol | 71-41-0 | FFA | 1.6–70 ppm 5 | Grilled, dust (dehulled pea) | %D 28 (1.0 µg/g) | (NO–3.6 µg/g) | ||
| 1-Penten-3-ol | 616-25-1 | FFA | 400 ppb 5 | Beany, green (whole lupin) | (NO–4.6 µg/g) | (NO–1.4 µg/g) | OI 29 (26 µg/g) | ||
| 2-Methoxyphenol (guaiacol) | 90-05-1 | 3–31 ppb 5 | Sweet, phenolic (whole lupin) | OI 2 (4 µg/g) | |||||
| 2-Ethylhexanol | 104-76-7 | Floral (pea protein) | %D 56 (Coelution) | ||||||
| Octanol | 117-87-5 | FFA | 42–480 ppb 5 | Mushroom, vegetable (dehulled pea) | %D 83 (0.8 µg/g) | (NO–2.5 µg/g) | |||
| 1-Octen-3-ol | 3391-86-4 | FFA | 14 ppb 5 | Mushroom, vegetable (peas) | %D 94 (1.7 µg/g) | %D 78 (4.2 µg/g) | (NO – 78 µg/g) | ||
| Nonanol | 143-08-8 | FFA | 50–90 ppm 5 | Pea, vegetable, silt, earth (dehulled pea) | %D 78 (0.8 µg/g) | (ND) | |||
| Ketones | 2-Acetyl-1-pyrroline | 85213-22-5 | AA 3 | 99,000 µg/L (water) 4 | Popcorn (dehulled lupin) | FD 32 | |||
| Maltol | 118-71-8 | AA 3 | 35,000 µg/L (water) 4 | Caramel (dehulled lupin) | (ND) | (NO – Coelution) | FD 256 | ||
| Sotolone | 28664-35-9 | AA 6 | 2.1 µg/kg (cellulose) 4 | Spicy, savoury (dehulled lupin) | FD 256 | ||||
| 2,3-Butanedione | 431-03-8 | FFA 4 | 0.3–15 ppb 5 | Sweet, caramel, vanilla (whole lupin) | OI 17 (79 µg/g) | ||||
| 4-Methyl-2-pentanone | 108-10-1 | FFA 4 | 240–540 ppb 5 | Sweet, fruity, bubble gum (whole lupin) | OI 5 (0 µg/g) | ||||
| 2-Pentylfuran | 3777-69-3 | FFA | 6 ppb 5 | Oat, nutty (whole lupin) | OI 5 (172 µg/g) | ||||
| 1-Penten-3-one | 1629-58-9 | FFA 4 | 400 ppb 5 | Soil, green, herbal (whole lupin) | OI 28 (32 µg/g) | ||||
| 1-Octen-3-one | 4312-99-6 | FFA 3 | 0.016 µg/L (water) 5 | Mushroom, fungi (lupins) | OI 49 (2 µg/g) | FD 32 | |||
| 3-Octen-2-one | 1669-44-9 | FFA | 5–4 µg/L (water) 4 | Soil, earthy (whole lupin) | (ND) | (NO–2.5 µg/g) | OI 13 (2 µg/g) | ||
| 2,3-Octanedione | 585-25-1 | Mushroom, vegetable (peas) | %D 94 (Trace) | %D 78 (4.1 µg/g) | |||||
| (Z)-Octan-1,5-dien-3-one | 65213-86-7 | FFA 3 | Geranium, metallic (dehulled lupin) | FD 128 | |||||
| (E,E)-3,5-Octadien-2-one | 30086-02-3 | FFA | 0.15 ppm 5 | Vegetable, hay, earth (pea isolate) | (ND) | %D 100 (24.4 µg/g) | |||
| Undecanone | 112-12-9 | 7–82 ppb 5 | Floral, fruity (peas) | %D 22 (Trace) | %D 39 (0.6 µg/g) | ||||
| β-Ionone | 79-77-6 | CAR | 3.5 µg/L (water) 4 | Violet, flowery (dehulled lupin) | FD 512 | ||||
| Acids | Acetic acid | 64-19-7 | AA | 99,000 µg/L (water) 4 | Vinegar (dehulled lupin) | FD 32 | |||
| Phenylacetic acid | 103-82-2 | AA 7 | 6100 µg/L (water) 4 | Beeswax, honey (dehulled lupin) | FD 256 | ||||
| 2-Methylbutanoic acid | 116-53-0 | N, AA | 2200 µg/L (water) 4 | Sweaty, fruity, cheese (dehulled lupin) | (ND) | (ND) | FD 2048 | ||
| 3-Methylbutanoic acid | 503-74-2 | AA | 490 µg/L (water) 4 | Animal (dehulled pea)/Sweaty, fruity, cheese (dehulled lupin) | %D 22 (0.3 µg/g) | (ND) | FD 2048 | ||
| Pentanoic acid | 109-52-4 | AA 3 | 11,000 µg/L (water) 4 | Sweaty, fruity, cheese (dehulled lupin) | (NO–0.3 µg/g) | (ND) | FD 32 | ||
| Hexanoic acid | 142-62-1 | FFA | 93 ppb–10 ppm 5 | Faeces, meat broth, sewer (pea isolate) | (NO–0.5 µg/g) | %D 83 (Coelution) | |||
| Heptanoic acid | 111-14-8 | 640 ppb–10.4 ppm 5 | Animal, vomit (pea isolate) | (NO–1.0 µg/g) | %D 22 (ND) | ||||
| Lactones | γ-Octalactone | 104-50-7 | FFA | 6.5 g/L (water) 4 | Floral, anise, mint (peas)/ Coconut, sweet (dehulled lupin) | %D 17 (Trace) | %D 94 (Trace) | FD 64 | |
| γ-Nonalactone | 104-61-0 | FFA | 9.7 µg/L (water) 4 | Sweet, candies, coconut (dehulled pea)/Coconut, sweet (dehulled lupin) | %D 72 (0.9 µg/g) | (NO – 1.0 µg/g) | FD 256 | ||
| 4-Hydroxy-2-noneic acid lactone | 21963-26-8 | FFA | Mint, strawberry, chlorophyll (peas) | %D 50 (0.5 µg/g) | %D 56 (3.2 µg/g) | ||||
| γ-Decalactone | 706-14-9 | FFA 3 | 1–11 ppb 5 | Peach, fruity (dehulled lupin) | FD 32 | ||||
| Pyrazines | 3-Isobutyl-2-methoxypyrazine | 24683-00-9 | AA | 62 µg/L (water) 4 | Woody, fresh pea (whole lupin)/Green pepper, earthy (dehulled lupin) | OI 70 (9 µg/g) | FD 32 | ||
| 2-Sec-Butyl-3-methoxypyrazine | 24168-70-5 | AA 4 | 1 ppb 5 | Woody, green (whole lupin) | OI 54 (1 µg/g) | ||||
| 3-Isopropyl-2-methoxypyrazine | 25773-40-4 | AA 4 | 39 µg/L (water) 4 | Soil, fresh green, woody (whole lupin)/Pea, green pepper (dehulled lupin) | OI 53 (0 µg/g) | FD 256 | |||
| 5or6-Methyl-3-isopropyl-2-methoxypyrazine | 32021-41-3 | AA | 3–7 ppb 5 | Plastic, cardboard, hay (dehulled pea)/Hay, dry, vegetable (pea isolate) | %D 50 (1.8 µg/g) | %D 83 (0.4 µg/g) | |||
| 3-Ethyl-2,5-dimethylpyrazine | 13360-65-1 | Potato, baked (whole lupin) | OI 65 (1 µg/g) | ||||||
| Others | Benzothiazole | 95-16-9 | AA 6 | 80–450 ppb 5 | Grilled meat (peas) | %D 39 (0.3 µg/g) | %D 39 (4.6 µg/g) | ||
| Dimethyl trisulfide | 3658-80-8 | 0.0099 µg/L (water) 4 | Faeces, meat broth, sewer (pea protein)/Meaty, metallic, sulphur (whole lupin) | (ND) | %D 83 (0.2 µg/g) | OI 14 (1 µg/g) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karolkowski, A.; Guichard, E.; Briand, L.; Salles, C. Volatile Compounds in Pulses: A Review. Foods 2021, 10, 3140. https://doi.org/10.3390/foods10123140
Karolkowski A, Guichard E, Briand L, Salles C. Volatile Compounds in Pulses: A Review. Foods. 2021; 10(12):3140. https://doi.org/10.3390/foods10123140
Chicago/Turabian StyleKarolkowski, Adeline, Elisabeth Guichard, Loïc Briand, and Christian Salles. 2021. "Volatile Compounds in Pulses: A Review" Foods 10, no. 12: 3140. https://doi.org/10.3390/foods10123140
APA StyleKarolkowski, A., Guichard, E., Briand, L., & Salles, C. (2021). Volatile Compounds in Pulses: A Review. Foods, 10(12), 3140. https://doi.org/10.3390/foods10123140