


The Bioaccessibility and Antioxidant Activities of Fermented Mango Cultivar Juices after Simulated In Vitro Digestion

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Mango Juice Preparation
2.3. In Vitro Digestion of Fermented MJs
2.4. Effect of In Vitro Digestion on the Survival of LAB in Fermented MJs
2.5. Total Phenolic Content
2.6. Carotenoid Profile of MJs after Digestion
2.7. Antioxidant Activities of Fermented MJs Digest
2.7.1. Ferric Reducing Antioxidant Power (FRAP)
2.7.2. 1,1-Diphenyl-2-picryl-hydrazyl (DPPH) of Fermented MJs Digest
2.7.3. 2,2-Azinobis-(3-ethyl-benzothiazoline-6-sulfonic acid (ABTS) of Fermented MJs Digest
2.8. Statistical Analysis
3. Results and Discussion
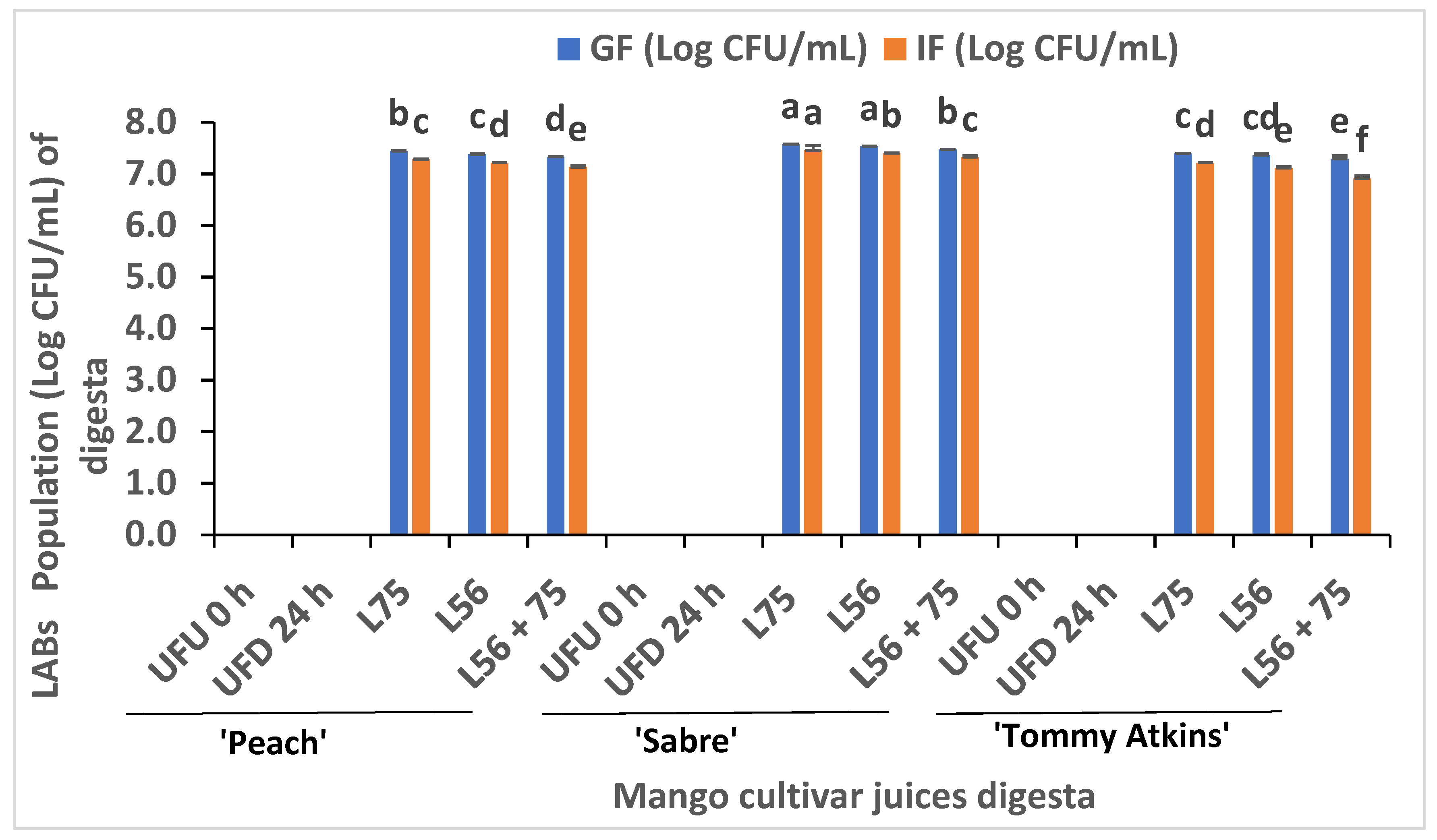
3.1. LAB Surviving Population in Fermented MJs after In Vitro GI Digestion
3.2. Changes in TPC of LAB Strains Fermented MJs from Different Cultivars after In Vitro GI Digestion
3.3. Changes in Carotenoids of MJs from Different Cultivars Fermented with LAB after In Vitro GI Digestion
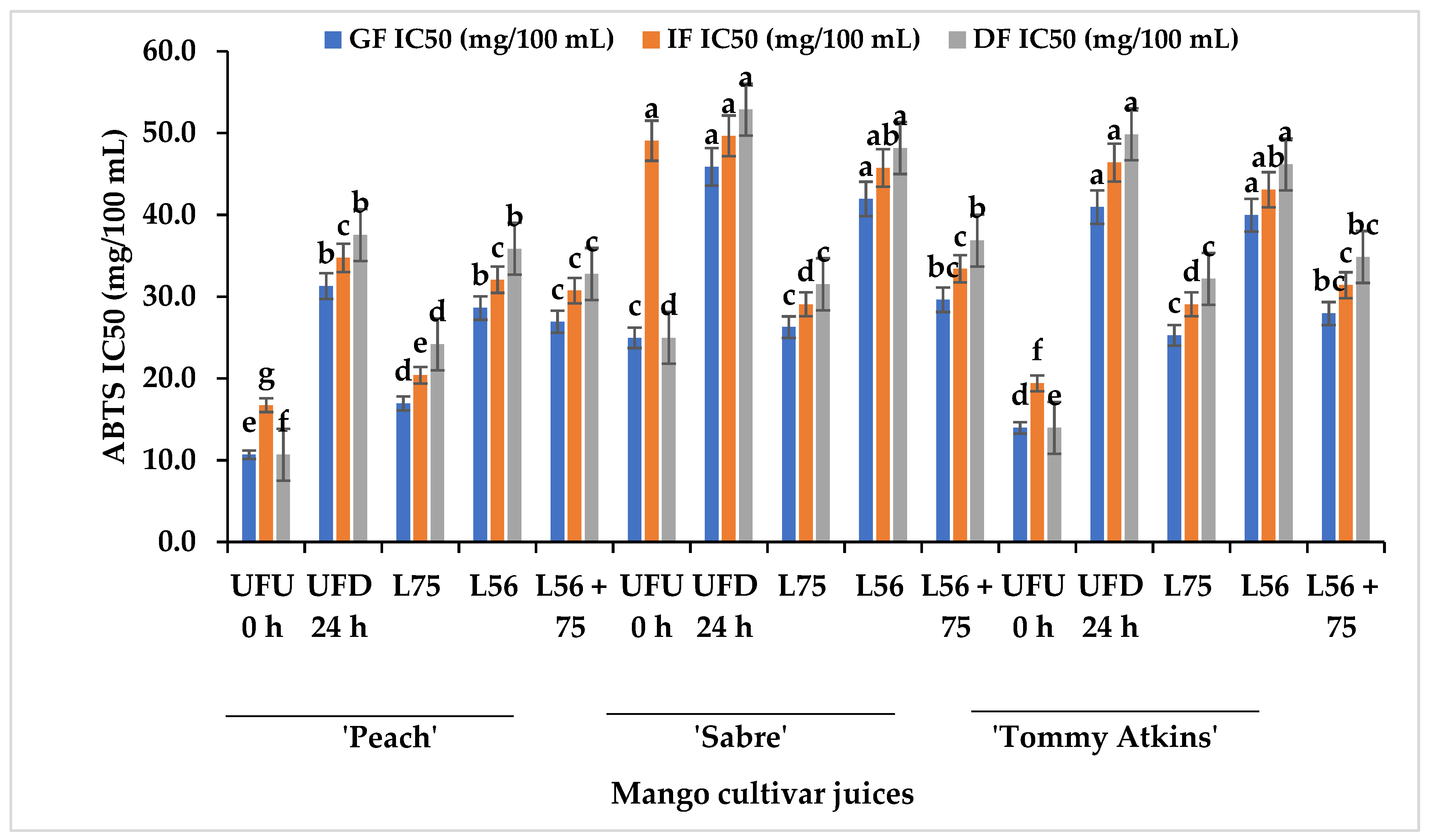
3.4. Changes in Antioxidant Activities of MJs from Different Cultivars Fermented with LAB after In Vitro GI Digestion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fivaz, J. Mango production in South Africa as compared to the rest of the world. Acta Hortic. 2009, 820, 29–46. [Google Scholar] [CrossRef]
- Bally, I.S. Advances in research and development of mango industry. Rev. Bras. Frutic. 2011, 33, 57–63. [Google Scholar] [CrossRef]
- Ntombela, S. South African Fruit Trade Flow Markets; Economic Research Center, National Agricultural Marketing Council: Pretoria, South Africa, 2014; Volume 14, pp. 1–15. [Google Scholar]
- De Sousa, A.S.B.; da Silva, M.C.A.; Lima, R.P.; de Albuquerque Meireles, B.R.L.; Cordeiro, A.T.M.; da Silva Santos, E.F.; Amaro, A.L.; Pintado, M.M.E.; de Melo Silva, S. Phenolic compounds and antioxidant activity as discriminating markers and adding value of mango varieties. Sci. Hortic. 2021, 287, 110259. [Google Scholar] [CrossRef]
- Manthey, J.A.; Perkins-Veazie, P. Influences of harvest date and location on the levels of β-carotene, ascorbic acid, total phenols, the in vitro antioxidant capacity, and phenolic profiles of five commercial varieties of mango (Mangifera indica L.). J. Agric. Food Chem. 2009, 57, 10825–10830. [Google Scholar] [CrossRef]
- Cele, N.P.; Akinola, S.A.; Manhivi, V.E.; Shoko, T.; Remize, F.; Sivakumar, D. Influence of Lactic Acid Bacterium Strains on Changes in Quality, Functional Compounds and Volatile Compounds of Mango Juice from Different Cultivars during Fermentation. Foods 2022, 11, 682. [Google Scholar] [CrossRef]
- Mapelli-Brahm, P.; Barba, F.J.; Remize, F.; Garcia, C.; Fessard, A.; Khaneghah, A.M.; Sant’Ana, A.S.; Lorenzo, J.M.; Montesano, D.; Meléndez-Martínez, A.J. The impact of fermentation processes on the production, retention and bioavailability of carotenoids: An overview. Trends Food Sci. Technol. 2020, 99, 389–401. [Google Scholar] [CrossRef]
- Wang, J.; Xie, B.; Sun, Z. Quality parameters and bioactive compound bioaccessibility changes in probiotics fermented mango juice using ultraviolet-assisted ultrasonic pre-treatment during cold storage. LWT 2021, 137, 110438. [Google Scholar] [CrossRef]
- Burton-Freeman, B.M.; Sandhu, A.K.; Edirisinghe, I. Mangos and their bioactive components: Adding variety to the fruit plate for health. Food Funct. 2017, 8, 3010–3032. [Google Scholar] [CrossRef]
- Managa, M.G.; Akinola, S.A.; Remize, F.; Garcia, C.; Sivakumar, D. Physicochemical parameters and bioaccessibility of lactic acid bacteria fermented chayote Leaf (Sechium edule) and pineapple (Ananas comosus) smoothies. Front. Nutr. 2021, 8, 649189. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Pannella, G.; Tipaldi, L.; Cozzolino, A.; Coppola, R.; Sorrentino, E. Survival of commercial probiotic strains in dark chocolate with high cocoa and phenols content during the storage and in a static in vitro digestion model. J. Funct. Foods 2017, 35, 60–67. [Google Scholar] [CrossRef]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef] [PubMed]
- Rocchetti, M.T.; Russo, P.; Capozzi, V.; Drider, D.; Spano, G.; Fiocco, D. Bioprospecting antimicrobials from Lactiplantibacillus plantarum: Key factors underlying its probiotic action. Int. J. Mol. Sci. 2021, 22, 12076. [Google Scholar] [CrossRef] [PubMed]
- Mashitoa, F.M.; Akinola, S.A.; Manhevi, V.E.; Garcia, C.; Remize, F.; Slabbert, R.; Sivakumar, D. Influence of Fermentation of Pasteurised Papaya Puree with Different Lactic Acid Bacterial Strains on Quality and bioaccessibility of phenolic compounds during in vitro digestion. Foods 2021, 10, 962. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharmacol. 2013, 75, 588–602. [Google Scholar] [CrossRef] [PubMed]
- Blancas-Benitez, F.J.; Mercado-Mercado, G.; Quirós-Sauceda, A.E.; Montalvo-González, E.; González-Aguilar, G.A.; Sáyago-Ayerdi, S.G. Bioaccessibility of polyphenols associated with dietary fiber and in vitro kinetics release of polyphenols in Mexican ‘Ataulfo’mango (Mangifera indica L.) by-products. Food Funct. 2015, 6, 859–868. [Google Scholar] [CrossRef]
- Moloto, M.R.; Phan, A.D.T.; Shai, J.L.; Sultanbawa, Y.; Sivakumar, D. Comparison of phenolic compounds, carotenoids, amino acid composition, in vitro antioxidant and anti-diabetic activities in the leaves of seven cowpea (Vigna unguiculata) cultivars. Foods 2020, 9, 1285. [Google Scholar] [CrossRef]
- Havenaar, R.; Huis In’t Veld, J.H. Probiotics: A general view. In The Lactic Acid Bacteria; Springer: Boston, MA, USA, 1992; Volume 1, pp. 151–170. [Google Scholar]
- Zendeboodi, F.; Khorshidian, N.; Mortazavian, A.M.; da Cruz, A.G. Probiotic: Conceptualization from a new approach. Curr. Opin. Food Sci. 2020, 32, 103–123. [Google Scholar] [CrossRef]
- Champagne, C.P.; Ross, R.P.; Saarela, M.; Hansen, K.F.; Charalampopoulos, D. Recommendations for the viability assessment of probiotics as concentrated cultures and in food matrices. Int. J. Food Microbiol. 2011, 149, 185–193. [Google Scholar] [CrossRef]
- Kumar, Y.; Singh, L. Health benefits of fermented and functional foods. J. Plant Dev. Sci. 2009, 1, 151–155. [Google Scholar]
- Muñoz, R.; de Las Rivas, B.; de Felipe, F.L.; Reverón, I.; Santamaría, L.; Esteban-Torres, M.; Curiel, J.; Rodríguez, H.; Landete, J. Biotransformation of phenolics by Lactobacillus plantarum in fermented foods. In Fermented Foods in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2017; pp. 63–83. [Google Scholar]
- Selma, M.V.; Espin, J.C.; Tomas-Barberan, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef]
- Oliveira-Alves, S.C.; Vendramini-Costa, D.B.; Cazarin, C.B.B.; Maróstica, M.R., Jr.; Ferreira, J.P.B.; Silva, A.B.; Prado, M.A.; Bronze, M.R. Characterization of phenolic compounds in chia (Salvia hispanica L.) seeds, fiber flour and oil. Food Chem. 2017, 232, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, R.; Zhang, Y.; Yang, Y.; Sun, X.; Zhang, Q.; Yang, N. Biotransformation of phenolics and metabolites and the change in antioxidant activity in kiwifruit induced by Lactobacillus plantarum fermentation. J. Sci. Food Agric. 2020, 100, 3283–3290. [Google Scholar] [CrossRef] [PubMed]
- Degrain, A.; Manhivi, V.; Remize, F.; Garcia, C.; Sivakumar, D. Effect of lactic acid fermentation on color, phenolic compounds and antioxidant activity in African nightshade. Microorganisms 2020, 8, 1324. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. Changes in vitamin C, phenolic, and carotenoid profiles throughout in vitro gastrointestinal digestion of a blended fruit juice. J. Agric. Food Chem. 2013, 61, 1859–1867. [Google Scholar] [CrossRef]
- Seke, F.; Manhivi, V.E.; Shoko, T.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. Effect of freeze drying and simulated gastrointestinal digestion on phenolic metabolites and antioxidant property of the Natal plum (Carissa macrocarpa). Foods 2021, 10, 1420. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; de Ancos, B.; Sánchez-Vega, R.; Sánchez-Moreno, C.; Cano, M.P.; Elez-Martínez, P.; Martín-Belloso, O. Food matrix and processing influence on carotenoid bioaccessibility and lipophilic antioxidant activity of fruit juice-based beverages. Food Funct. 2016, 7, 380–389. [Google Scholar] [CrossRef]
- Biehler, E.; Kaulmann, A.; Hoffmann, L.; Krause, E.; Bohn, T. Dietary and host-related factors influencing carotenoid bioaccessibility from spinach (Spinacia oleracea). Food Chem. 2011, 125, 1328–1334. [Google Scholar] [CrossRef]
- Courraud, J.; Berger, J.; Cristol, J.-P.; Avallone, S. Stability and bioaccessibility of different forms of carotenoids and vitamin A during in vitro digestion. Food Chem. 2013, 136, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, J.N.; Luu, A.Y.; Dragsted, L.O.; Arrigoni, E. Adaption of an in vitro digestion method to screen carotenoid liberation and in vitro accessibility from differently processed spinach preparations. Food Chem. 2017, 224, 407–413. [Google Scholar] [CrossRef]
- Saxholt, E.; Christensen, A.; Møller, A.; Hartkopp, H.; Hess Ygil, K.; Hels, O. Danish Food Composition Databank; Revision 7; Department of Nutrition, National Food Institute, Technical University of Denmark: Lyngby, Denmark, 2008. [Google Scholar]
- Jeffery, J.L.; Turner, N.D.; King, S.R. Carotenoid bioaccessibility from nine raw carotenoid-storing fruits and vegetables using an in vitro model. J. Sci. Food Agric. 2012, 92, 2603–2610. [Google Scholar] [CrossRef]
- Bartkiene, E.; Vidmantiene, D.; Juodeikiene, G.; Viskelis, P.; Urbonaviciene, D. Lactic acid fermentation of tomato: Effects on cis/trans lycopene isomer ratio, β-carotene mass fraction and formation of L (+)-and D (−)-lactic acid. Food Technol. Biotechnol. 2013, 51, 471–478. [Google Scholar]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin–Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Zhang, G.; Yan, X.; Wu, S.; Ma, M.; Yu, P.; Gong, D.; Deng, S.; Zeng, Z. Ethanol extracts from Cinnamomum camphora seed kernel: Potential bioactivities as affected by alkaline hydrolysis and simulated gastrointestinal digestion. Food Res. Int. 2020, 137, 109363. [Google Scholar] [CrossRef]
- Valero-Cases, E.; Nuncio-Jáuregui, N.; Frutos, M.J. Influence of fermentation with different lactic acid bacteria and in vitro digestion on the biotransformation of phenolic compounds in fermented pomegranate juices. J. Agric. Food Chem. 2017, 65, 6488–6496. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Reginio, F.C., Jr.; Thuengtung, S.; Ogawa, Y. Changes in bioactive compounds and antioxidant activity of plant-based foods by gastrointestinal digestion: A review. Crit. Rev. Food Sci. Nutr. 2021, 62, 4684–4705. [Google Scholar] [CrossRef]
- Koca Bozalan, N.; Karadeniz, F. Carotenoid profile, total phenolic content, and antioxidant activity of carrots. Int. J. Food Prop. 2011, 14, 1060–1068. [Google Scholar] [CrossRef]
- Wang, C.; Wu, H.; Liu, Z.; Barrow, C.; Dunshea, F.; Suleria, H.A.R. Bioaccessibility and movement of phenolic compounds from tomato (Solanum lycopersicum) during in vitro gastrointestinal digestion and colonic fermentation. Food Funct. 2022, 13, 4954–4966. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mango Juices (MJs) | GF (mg/100 mL) | Fold Decrease | Bioacc % | IF (mg/100 mL) | Fold Decrease | Bioacc % | DF (mg/100 mL) | Fold Decrease | Bioacc % |
|---|---|---|---|---|---|---|---|---|---|
| ‘Peach’ | |||||||||
| UFU 0 h | 1171.41 ± 0.57 a | 1171.41 ± 0.57 a | 1171.41 ± 0.57 a | ||||||
| UFD 24 h | 612.50 ± 0.57 k | 1.91 ± 0.05 c | 52.29 | 314.06 ± 0.1 n | 3.73 ± 0.10 b | 26.81 | 216.68 ± 0.53 n | 5.41 ± 0.50 a | 18.50 |
| L75 | 822.17 ± 0.50 g | 1.42 ± 0.05 h | 70.19 | 511.45 ± 0.32 g | 2.29 ± 0.31 i | 43.66 | 366.21 ± 0.52 g | 3.20 ± 0.50 i | 31.26 |
| L56 | 785.22 ± 0.56 i | 1.49 ± 0.5 f | 67.03 | 508.89 ± 0.18 h | 2.30 ± 0.16 h | 43.44 | 336.53 ± 0.55 i | 3.48 ± 0.55 e | 28.73 |
| L56 + 75 | 821.80 ± 0.47 h | 1.43 ± 0.15 g | 70.15 | 452.15 ± 0.57 i | 2.59 ± 0.57 g | 38.60 | 342.58 ± 0.10 h | 3.42 ± 0.15 f | 29.25 |
| ‘Sabre’ | |||||||||
| UFU 0 h | 1167.03 ± 0.17 b | 1167.03 ± 0.17 b | 1167.03 ± 0.17 b | ||||||
| UFD 24 h | 690.95 ± 0.52 j | 1.69 ± 0.05 e | 59.21 | 387.02 ± 0.55 j | 3.02 ± 0.55 c | 33.16 | 226.56 ± 0.57 m | 5.15 ± 0.53 c | 19.41 |
| L75 | 947.19 ± 0.50 c | 1.23 ± 0.10 j | 81.16 | 649.00 ± 0.15 d | 1.80 ± 0.16 l | 55.61 | 399.46 ± 0.57 d | 2.92 ± 0.57 l | 34.23 |
| L56 | 882.84 ± 0.57 e | 1.32 ± 0.15 i | 75.65 | 584.74 ± 0.28 e | 2.00 ± 0.25 k | 50.10 | 379.57 ± 0.57 f | 3.07 ± 0.55 j | 32.52 |
| L56 + 75 | 881.79 ± 0.55 f | 1.32 ± 0.05 i | 75.56 | 579.13 ± 0.81 f | 2.02 ± 0.80 j | 49.62 | 387.94 ± 0.57 e | 3.01 ± 0.26 k | 33.24 |
| ‘Tommy Atkins’ | |||||||||
| UFU 0 h | 923.06 ± 0.44 d | 923.06 ± 0.44 c | 923.06 ± 0.44 c | ||||||
| UFD 24 h | 372.19 ± 0.57 o | 2.48 ± 0.05 a | 40.32 | 241.77 ± 0.57 o | 3.82 ± 0.57 a | 26.19 | 172.95 ± 0.57 o | 5.34 ± 0.40 b | 18.74 |
| L75 | 496.02 ± 0.27 l | 1.86 ± 0.05 d | 53.74 | 329.58 ± 3.56 k | 2.80 ± 3.45 f | 35.71 | 283.22 ± 0.57 j | 3.26 ± 0.47 h | 30.68 |
| L56 | 479.51 ± 0.50 m | 1.93 ± 0.15 b | 51.95 | 318.79 ± 0.57 l | 2.90 ± 0.57 e | 34.54 | 270.83 ± 0.57 k | 3.41 ± 0.85 g | 29.34 |
| L56 + 75 | 478.08 ± 0.35 n | 1.93 ± 0.05 b | 51.79 | 314.83 ± 0.13 m | 2.93 ± 0.15 d | 34.11 | 251.23 ± 1.15 l | 3.67 ± 0.20 d | 27.22 |
| LSD | 0.51 *** | 0.15 *** | 0.18 *** | 0.31 *** | 0.17 *** | 0.53 *** |
| Mango Cultivar Juices (MJs) | GF (mg/mL) | Fold Increase/Decrease | Bioacc (%) | IF (mg/mL) | Fold Decrease | Bioacc (%) |
| β-carotene concentration | ||||||
| ‘Peach’ | ||||||
| UFU 0 h | 61.20 ± 0.57 e | 61.20 ± 0.57 a | ||||
| UFD 24 h | 70.63 ± 0.37 d | 1.15 ± 0.01 h | 115.41 | 41.83 ± 0.57 e | −1.46 ± 0.01 b | 68.35 |
| L75 | 74.43 ± 0.40 c | 1.21 ± 0.01 g | 121.62 | 48.63 ± 0.57 c | −1.26 ± 0.01 g | 79.46 |
| L56 | 80.63 ± 0.30 a | 1.32 ± 0.01 e | 131.75 | 50.43 ± 0.57 b | −1.21 ± 0.01 h | 82.40 |
| L56 + 75 | 77.96 ± 0.22 b | 1.27 ± 0.01 f | 127.39 | 44.86 ± 0.57 d | −1.36 ± 0.01 e | 73.30 |
| ‘Sabre’ | ||||||
| UFU 0 h | 19.9 ± 0.57 l | 19.9 ± 0.57 g | ||||
| UFD 24 h | 28.63 ± 0.49 i | 1.44 ± 0.01 d | 143.87 | 14.83 ± 0.57 m | −1.34 ± 0.01 f | 74.52 |
| L75 | 34.73 ± 0.57 h | 1.75 ± 0.01 c | 174.52 | 18.07 ± 0.57 k | −1.10 ± 0.01 i | 90.80 |
| L56 | 51.53 ± 0.57 f | 2.59 ± 0.01 a | 258.94 | 19.7 ± 0.57 h | −1.01 ± 0.01 k | 98.99 |
| L56 + 75 | 48.73 ± 0.57 g | 2.45 ± 0.01 b | 244.87 | 18.8 ± 0.57 i | −1.06 ± 0.01 j | 94.47 |
| ‘Tommy Atkins’ | ||||||
| UFU 0 h | 20.2 ± 0.57 k | 20.2 ± 0.57 f | ||||
| UFD 24 h | 14.4 ± 0.1 o | −1.40 ± 0.01 m | 71.29 | 12.63 ± 0.57 o | −1.60 ± 0.01 a | 62.52 |
| L75 | 16.83 ± 0.57 m | −1.20 ± 0.01 k | 83.32 | 14.54 ± 0.57 m | −1.39 ± 0.01 d | 71.98 |
| L56 | 25.83 ± 0.57 j | 1.27 ± 0.01 i | 127.87 | 18.35 ± 0.57 j | −1.10 ± 0.01 i | 90.84 |
| L56 + 75 | 15.83 ± 0.57 n | −1.28 ± 0.01 l | 78.37 | 14.11 ± 0.57 n | −1.43 ± 0.01 c | 69.85 |
| LSD | 0.12 ** | 0.11 ** | 0.16 ** | 0.05 ** | ||
| Trans-β-carotene concentration | ||||||
| Mango juice | GF (µg/mL) | Fold increase | Bioacc (%) | IF (µg/mL) | Fold increase/decrease | Bioacc (%) |
| ‘Peach’ | ||||||
| UFU 0 h | 5.27 ± 0.05 f | 5.26 ± 0.17 b | ||||
| UFD 24 h | 6.03 ± 0.1 e | 1.14 ± 0.11 f | 114.42 | 4.00 ± 0.13 f | −1.31 ± 0.13 e | 76.4 |
| L75 | 7.23 ± 0.05 b | 1.37 ± 0.05 e | 137.19 | 4.23 ± 0.15 d | −1.24 ± 0.15 d | 75.54 |
| L56 | 9.17 ± 0.25 a | 1.74 ± 0.21 b | 174.00 | 7.2 ± 0.27 a | 1.37 ± 0.11 a | 136.88 |
| L56 + 75 | 6.30 ± 0.51 c | 1.19 ± 0.45 f | 119.54 | 4.3 ± 0.19 d | 1.22 ± 0.11 b | 81.74 |
| ‘Sabre’ | ||||||
| UFU 0 h | 4.03 ± 0.05 i | 4.03 ± 0.19 f | ||||
| UFD 24 h | 4.27 ± 0.12 h | 1.05 ± 0.30 g | 105.95 | 2.26 ± 0.23 h | −1.78 ± 0.21 f | 56.07 |
| L75 | 5.27 ± 0.23 f | 1.30 ± 0.31 e | 130.76 | 3.2 ± 0.27 g | −1.25 ± 0.22 d | 79.4 |
| L56 | 7.20 ± 0.55 b | 1.78 ± 0.42 b | 178.66 | 4.4 ± 0.57 c | 1.09 ± 0.16 c | 109.18 |
| L56 + 75 | 6.23 ± 0.45 cd | 1.54 ± 0.18 c | 154.59 | 4.23 ± 0.15 d | 1.04 ± 0.20 c | 104.96 |
| ‘Tommy Atkins’ | ||||||
| UFU 0 h | 3.03 ± 0.03 K | 3.03 ± 0.07 g | ||||
| UFD 24 h | 3.27 ± 0.07 J | 1.07 ± 0.41 g | 107.92 | 1.26 ± 0.24 i | −2.40 ± 0.32 g | 41.58 |
| L75 | 4.27 ± 0.43 h | 1.4 ± 0.22 d | 140.92 | 2.26 ± 0.30 h | −1.34 ± 0.33 e | 74.58 |
| L56 | 6.17 ± 0.11 d | 2.03 ± 0.53 a | 203.63 | 4.16 ± 0.48 e | 1.37 ± 0.21 a | 137.29 |
| L56 + 75 | 5.17 ± 0.75 g | 1.70 ± 0.48 b | 170.62 | 3.16 ± 0.03 g | 1.04 ± 0.11 c | 104.29 |
| LSD | 0.01 ** | 0.01 ** | 0.03 ** | 0.02 ** | 0.04 ** | 0.08 ** |
| Zeaxanthin concentrations | ||||||
| Mango juice | GF (µg/mL) | Fold increase | Bioacc (%) | GF (µg/mL) | Fold increase/decrease | Bioacc (%) |
| ‘Peach’ | ||||||
| UFU 0 h | 4.26 ± 0.25 e | 4.26 ± 0.25 b | ||||
| UFD 24 h | 6.23 ± 0.35 b | 1.46 ± 0.57 e | 146.2 | 4.23 ± 0.23 b | −1.00 ± 0.25 e | 99.29 |
| L75 | 5.03 ± 0.55 d | 1.18 ± 0.21 h | 118.07 | 3.03 ± 0.53 d | −1.41 ± 0.53 f | 71.12 |
| L56 | 8.16 ± 0.25 a | 1.91 ± 0.09 b | 191.54 | 6.16 ± 0.16 a | 1.44 ± 0.15 b | 144.60 |
| L56 + 75 | 5.3 ± 0.32 c | 1.24 ± 0.34 g | 124.41 | 3.3 ± 0.30 c | −1.29 ± 0.35 f | 77.46 |
| ‘Sabre’ | ||||||
| UFU 0 h | 3.13 ± 0.51 g | 3.13 ± 0.51 c | ||||
| UFD 24 h | 3.43 ± 0.33 f | 1.09 ± 0.11 i | 109.58 | 1.43 ± 0.35 g | −2.18 ± 0.35 h | 45.68 |
| L75 | 4.26 ± 0.75 e | 1.36 ± 0.43 f | 136.10 | 2.26 ± 0.22 e | −1.38 ± 0.22 f | 72.20 |
| L56 | 6.23 ± 0.09 b | 1.99 ± 0.19 a | 199.04 | 4.23 ± 0.22 b | 1.35 ± 0.23 c | 135.14 |
| L56 + 75 | 5.23 ± 0.32 c | 1.67 ± 0.27 c | 167.09 | 3.23 ± 0.37 c | 1.03 ± 0.38 d | 103.19 |
| ‘Tommy Atkins’ | ||||||
| UFU 0 h | 2.06 ± 0.05 i | 2.06 ± 0.05 f | ||||
| UFD 24 h | 2.26 ± 0.83 h | 1.09 ± 0.21 i | 109.71 | 0.26 ± 0.09 i | −7.92 ± 0.09 i | 12.62 |
| L75 | 3.2 ± 0.52 g | 1.55 ± 0.60 d | 155.33 | 1.2 ± 0.23 h | −1.71 ± 0.24 g | 58.25 |
| L56 | 5.16 ± 0.77 c | 2.50 ± 0.17 a | 250.48 | 3.16 ± 0.65 c | 1.53 ± 0.55 a | 153.39 |
| L56 + 75 | 4.16 ± 0.06 e | 2.01 ± 0.14 a | 201.94 | 2.16 ± 0.15 f | 1.04 ± 0.16 d | 104.85 |
| LSD | 0.08 ** | 0.05 ** | 0.02 ** | 0.02 ** | 0.01 ** | 0.03 ** |
| Lutein concentration | ||||||
| Mango juice | GF (µg/mL) | Fold decrease | Bioacc (%) | IF (µg/mL) | Fold increase/ decrease | Bioacc (%) |
| ‘Peach’ | ||||||
| UFU 0 h | 3.93 ± 0.15 d | 3.93 ± 0.15 b | ||||
| UFD 24 h | 5.23 ± 0.23 b | 1.33 ± 0.25 e | 133.03 | 3.23 ± 0.20 b | −1.21 ± 0.20 d | 82.18 |
| L75 | 5.03 ± 0.09 b | 1.27 ± 0.10 f | 127.98 | 3.03 ± 0.05 b | −1.29 ± 0.05 d | 77.09 |
| L56 | 6.5 ± 0.63 a | 1.65 ± 0.09 b | 165.39 | 6.16 ± 0.18 a | 1.56 ± 0.20 a | 156.74 |
| L56 + 75 | 4.63 ± 0.36 c | 1.17 ± 0.35 g | 117.81 | 2.63 ± 0.36 c | −1.49 ± 0.38 d | 66.92 |
| ‘Sabre’ | ||||||
| UFU 0 h | 2.8 ± 0.25 e | 2.8 ± 0.25 c | ||||
| UFD 24 h | 3.1 ± 0.31 d | 1.10 ± 0.31 h | 110.71 | 0.76 ± 0.63 | −3.68 ± 0.60 e | 27.14 |
| L75 | 3.93 ± 0.35 d | 1.40 ± 0.33 d | 140.35 | 1.93 ± 0.33 d | −1.45 ± 0.33 d | 68.92 |
| L56 | 5.23 ± 0.55 b | 1.86 ± 0.53 a | 186.78 | 3.23 ± 0.28 b | 1.15 ± 0.29 b | 115.35 |
| L56 + 75 | 4.56 ± 0.65 c | 1.62 ± 0.55 b | 162.85 | 2.9 ± 0.28 c | 1.03 ± 0.28 c | 103.57 |
| ‘Tommy Atkins’ | ||||||
| UFU 0 h | 2.4 ± 0.19 e | 2.4 ± 0.19 c | ||||
| UFD 24 h | 2.6 ± 0.20 e | 1.08 ± 0.25 i | 108.33 | 0.6 ± 0.35 e | −4.00 ± 0.36 f | 25.00 |
| L75 | 2.8 ± 0.82 e | 1.16 ± 0.85 h | 116.66 | 0.53 ± 0.30 e | -4.52 ± 0.30 g | 22.08 |
| L56 | 4.5 ± 0.95 c | 1.87 ± 0.95 a | 187.5 | 2.5 ± 0.25 c | 1.04 ± 0.24 c | 104.16 |
| L56 + 75 | 3.83 ± 0.33 d | 1.59 ± 0.32 c | 159.58 | 1.83 ± 0.30 d | −1.31 ± 0.30 d | 76.25 |
| LSD | 0.01 ** | 0.04 ** | 0.44 ** | 0.01 ** | 0.01 ** | 0.03 ** |
| Alpha carotene concentration | ||||||
| Mango juice | GF (µg/mL) | Fold increase | Bioacc (%) | IF (µg/mL) | Fold increase/decrease | Bioacc (%) |
| ‘Peach’ | ||||||
| UFU 0 h | 4.93 ± 0.35 d | 4.93 ± 0.05 b | ||||
| UFD 24 h | 5.36 ± 0.13 c | 1.08 ± 0.15 g | 108.72 | 4.23 ± 0.30 b | −1.16 ± 0.30 c | 85.80 |
| L75 | 5.63 ± 0.65 c | 1.14 ± 0.64 f | 114.19 | 3.36 ± 0.22 c | −1.46 ± 0.20 d | 68.15 |
| L56 | 7.5 ± 0.55 a | 1.52 ± 0.56 b | 152.12 | 5.5 ± 0.15 a | 1.11 ± 0.15 a | 111.56 |
| L56 + 75 | 6.23 ± 0.13 b | 1.26 ± 0.14 e | 126.36 | 2.96 ± 0.23 d | −1.66 ± 0.25 d | 60.04 |
| ‘Sabre’ | ||||||
| UFU 0 h | 4.03 ± 0.05 d | 4.03 ± 0.05 b | ||||
| UFD 24 h | 3.93 ± 0.33 e | −1.02 ± 0.30 h | 95.51 | 2.26 ± 0.22 d | −1.78 ± 0.24 d | 56.07 |
| L75 | 4.93 ± 0.15 d | 1.22 ± 0.15 e | 122.33 | 2.93 ± 0.13 d | −1.37 ± 0.12 cd | 72.70 |
| L56 | 6.2 ± 0.27 b | 1.53 ± 0.27 b | 153.84 | 4.56 ± 0.25 b | 1.13 ± 0.25 a | 113.15 |
| L56 + 75 | 5.56 ± 0.45 c | 1.37 ± 0.46 d | 137.96 | 4.2 ± 0.12 b | 1.04 ± 0.12 b | 104.21 |
| ‘Tommy Atkins’ | ||||||
| UFU 0 h | 3.36 ± 0.25 e | 3.36 ± 0.25 c | ||||
| UFD 24 h | 3.6 ± 0.09 e | 1.07 ± 0.10 g | 107.14 | 1.26 ± 0.15 e | −2.66 ± 0.16 e | 37.5 |
| L75 | 3.93 ± 0.45 e | 1.16 ± 0.50 f | 116.96 | nd | nd | nd |
| L56 | 5.5 ± 0.15 c | 1.63 ± 0.14 a | 163.69 | 3.5 ± 0.45 c | 1.04 ± 0.28 b | 104.16 |
| L56 + 75 | 4.83 ± 0.12 d | 1.43 ± 0.12 c | 143.75 | nd | nd | nd |
| LSD | 0.06 * | 0.02 ** | 0.01 ** | 0.05 ** | 0.02 ** | |
| Mango Juices | GF (uM TEAC/mL) | Fold Decrease | Bioacc % | IF (uM TEAC/mL) | Fold Decrease | Bioacc % | DF (uM TEAC/mL) | Fold Decrease | Bioacc % |
|---|---|---|---|---|---|---|---|---|---|
| ‘Peach’ | |||||||||
| UFU 0 h | 354.19 ± 5.74 b | 354.19 ± 5.74 b | 354.19 ± 5.74 a | ||||||
| UFD 24 h | 235.17 ± 3.01 f | −1.51 ± 0.31 c | 66.40 | 102.51 ± 42.63 f | −3.46 ± 0.19 d | 28.94 | 55.84 ± 2.30 d | −6.34 ± 0.30 d | 15.77 |
| L75 | 250.06 ± 7.00 d | −1.42 ± 0.06 b | 70.60 | 142.95 ± 17.05 de | −2.48 ± 0.05 b | 40.36 | 58.95 ± 3.67 cd | −6.01 ± 0.50 c | 16.64 |
| L56 | 249.4 ± 14.24 d | −1.42 ± 0.04 b | 70.41 | 126.4 ± 0.57 e | −2.80 ± 0.70 c | 35.69 | 56.95 ± 2.14 d | −6.22 ± 0.15 d | 16.08 |
| L56+75 | 238.73 ± 9.43 f | −1.48 ± 0.23 c | 67.40 | 137.4 ± 4.16 de | −2.58 ± 0.16 b | 38.79 | 55.84 ± 2.90 d | −6.34 ± 0.20 d | 15.77 |
| ‘Sabre’ | |||||||||
| UFU 0 h | 356.91 ± 7.42 a | 356.91 ± 10.33 a | 356.91 ± 0.76 a | ||||||
| UFD 24 h | 238.06 ± 4.07 f | −1.50 ± 0.07 c | 66.70 | 124.28 ± 10.02 e | −2.87 ± 0.22 c | 34.82 | 54.95 ± 3.00 d | −6.50 ± 0.58 e | 15.40 |
| L75 | 252.73 ± 30.30 d | −1.41 ± 0.40 b | 70.81 | 152.51 ± 28.83 d | −2.34 ± 0.08 a | 42.73 | 67.4 ± 3.28 c | −5.30 ± 0.25 b | 18.88 |
| L56 | 251.17 ± 31.67 d | −1.42 ± 0.61 b | 70.37 | 127.17 ± 2.34 e | −2.81 ± 0.41 c | 35.63 | 65.4 ± 4.72 c | −5.46 ± 0.20 c | 18.32 |
| L56 + 75 | 247.84 ± 3.01 de | −1.44 ± 0.13 b | 69.44 | 139.4 ± 11.37 de | −2.56 ± 0.13 b | 39.06 | 63.62 ± 1.53 c | −5.61 ± 0.06 c | 17.83 |
| ‘Tommy Atkins’ | |||||||||
| UFU 0 h | 297.74 ± 48.07 c | 297.74 ± 48.07 c | 297.74 ± 48.07 b | ||||||
| UFD 24 h | 197.4 ± 44.77 g | −1.51 ± 0.17 c | 66.30 | 96.73 ± 34.11 f | −3.08 ± 0.11 d | 32.49 | 41.4 ± 1.38 e | −7.19 ± 0.33 f | 13.90 |
| L75 | 244.73 ± 8.00 e | −1.22 ± 0.33 a | 82.20 | 133.73 ± 0.57 e | −2.23 ± 0.51 a | 44.92 | 60.73 ± 8.44 c | −4.90 ± 0.24 a | 20.40 |
| L56 | 241.4 ± 8.22 e | −1.23 ± 0.28 a | 81.08 | 104.73 ± 34.66 f | −2.84 ± 0.33 c | 35.17 | 46.95 ± 10.79 e | −6.34 ± 0.68 d | 15.77 |
| L56 + 75 | 241.4 ± 6.93 e | −1.23 ± 0.31 a | 81.08 | 127.84 ± 2.1 e | −2.33 ± 0.12 a | 42.94 | 44.95 ± 1.53 e | −6.62 ± 0.33 e | 15.10 |
| LSD | 3.46 ** | 0.01 ** | 3.48 ** | 0.22 ** | 3.49 ** | 0.33 ** | |||
| TPC | TCC | FRAP | ABTS | DPPH | |
|---|---|---|---|---|---|
| TPC | 1 | ||||
| TCC | 0.41 | 1 | |||
| FRAP | 0.92 | 0.44 | 1 | ||
| ABTS | 0.68 | −0.31 | −0.56 | 1 | |
| DPPH | 0.61 | 0.15 | −0.52 | 0.23 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cele, N.P.; Akinola, S.A.; Shoko, T.; Manhevi, V.E.; Remize, F.; Sivakumar, D. The Bioaccessibility and Antioxidant Activities of Fermented Mango Cultivar Juices after Simulated In Vitro Digestion. Foods 2022, 11, 2702. https://doi.org/10.3390/foods11172702
Cele NP, Akinola SA, Shoko T, Manhevi VE, Remize F, Sivakumar D. The Bioaccessibility and Antioxidant Activities of Fermented Mango Cultivar Juices after Simulated In Vitro Digestion. Foods. 2022; 11(17):2702. https://doi.org/10.3390/foods11172702
Chicago/Turabian StyleCele, Nobahle P., Stephen A. Akinola, Tinotenda Shoko, Vimbainashe E. Manhevi, Fabienne Remize, and Dharini Sivakumar. 2022. "The Bioaccessibility and Antioxidant Activities of Fermented Mango Cultivar Juices after Simulated In Vitro Digestion" Foods 11, no. 17: 2702. https://doi.org/10.3390/foods11172702
APA StyleCele, N. P., Akinola, S. A., Shoko, T., Manhevi, V. E., Remize, F., & Sivakumar, D. (2022). The Bioaccessibility and Antioxidant Activities of Fermented Mango Cultivar Juices after Simulated In Vitro Digestion. Foods, 11(17), 2702. https://doi.org/10.3390/foods11172702