

Complete Genome Sequencing Revealed the Potential Application of a Novel Weizmannia coagulans PL-W Production with Promising Bacteriocins in Food Preservative

Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Bacterial Culture Conditions
2.2. Isolation of Antimicrobial Substance-Producing W. coagulans and Crude Antimicrobial Substance Preparation
2.3. Characteristics of Antimicrobial Substance Production in W. coagulans PL-W
2.3.1. Kinetics of Growth and Crude Antimicrobial Substance Production in W. coagulans PL-W
2.3.2. Physicochemical Properties of Crude Antimicrobial Substance
2.3.3. Antimicrobial Spectrum Assay of Crude Bacteriocins
2.3.4. Purification of Bacteriocin
2.4. Genome Sequencing, Assembly, Annotation, and Classification
2.5. Prediction of the Safety of W. coagulans PL-W
2.6. Prediction of the Probiotic Characteristics of W. coagulans PL-W
2.7. Statistical Analysis
3. Results and Discussion
3.1. Screening and Identification of Antimicrobial Substance-Producing Strains
3.2. Crude Antimicrobial Substance Production Properties of W. coagulans PL-W
3.2.1. Kinetics of Growth and Crude Antimicrobial Substance Production in W. coagulans PL-W
3.2.2. Characterization of Crude Antimicrobial Substance
3.2.3. Antibacterial Spectrum of Crude Bacteriocins
3.2.4. Purification of Bacteriocin
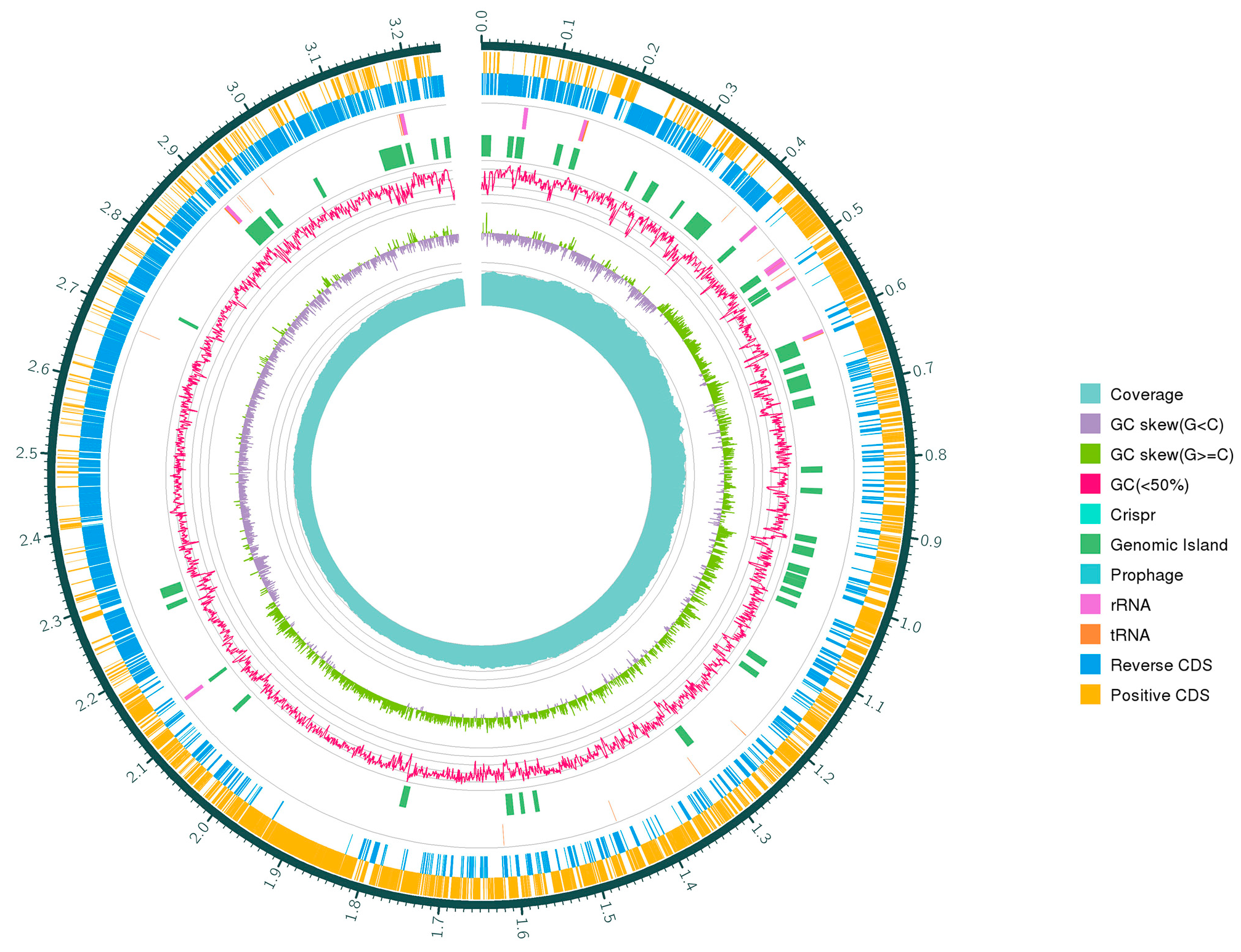
3.3. General Genome Features of W. coagulans PL-W
3.4. Taxonomic Classification and Phylogeny
3.5. Safety Assessment of W. coagulans PL-W
3.6. Assessment of Probiotic Properties
3.7. Antimicrobial Compound Gene Prediction and Validation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, W.; Pires, S.M.; Liu, Z.; Ma, X.; Guo, Y. Surveillance of foodborne disease outbreaks in China, 2003–2017. Food Control 2020, 118, 107359. [Google Scholar] [CrossRef]
- McEwen, S.A.; Collignon, P.J. Antimicrobial resistance: A one health perspective. Microbiol. Spectr. 2018, 6, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Blaser, M. Antibiotic overuse: Stop the killing of beneficial bacteria. Nature 2011, 476, 393–394. [Google Scholar] [CrossRef]
- Heilbronner, S.; Krismer, B.; Brotz-Oesterhelt, H.; Peschel, A. The microbiome-shaping roles of bacteriocins. Nat. Rev. Microbiol. 2021, 19, 726–739. [Google Scholar] [CrossRef]
- Ahmad, V.; Khan, M.S.; Jamal, Q.M.S.; Alzohairy, M.A.; Al Karaawi, M.A.; Siddiqui, M.U. Antimicrobial potential of bacteriocins: In therapy, agriculture and food preservation. Int. J. Antimicrob. Agents 2017, 49, 1–11. [Google Scholar] [CrossRef]
- Choi, G.H.; Holzapfel, W.H.; Todorov, S.D. Diversity of the bacteriocins, their classification and potential applications in combat of antibiotic resistant and clinically relevant pathogens. Crit. Rev. Microbiol. 2022, 48, 1–20. [Google Scholar] [CrossRef]
- O’Connor, P.M.; Kuniyoshi, T.M.; Oliveira, R.P.; Hill, C.; Ross, R.P.; Cotter, P.D. Antimicrobials for food and feed; a bacteriocin perspective. Curr. Opin. Biotechnol. 2020, 61, 160–167. [Google Scholar] [CrossRef]
- Ko, K.Y.; Park, S.R.; Lim, H.S.; Park, S.J.; Kim, M. Improved pretreatment method for determination of Nisins A and Z and monitoring in cheese using liquid chromatrography-tandem mass spectrometry. Food Anal. Method 2016, 9, 122–130. [Google Scholar] [CrossRef]
- Santiago-Silva, P.; Soares, N.F.F.; Nobrega, J.E.; Junior, M.A.W.; Barbosa, K.B.F.; Volp, A.C.P.; Zerdas, E.R.M.A.; Wurlitzer, N.J. Antimicrobial efficiency of film incorporated with pediocin (ALTA (R) 2351) on preservation of sliced ham. Food Control 2009, 20, 85–89. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A viable alternative to antibiotics? Nat. Rev. Microbiol. 2013, 11, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Moll, G.N.; Konings, W.N.; Driessen, A.J. Bacteriocins: Mechanism of membrane insertion and pore formation. Antonie Van Leeuwenhoek 1999, 76, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Babasaki, K.; Takao, T.; Shimonishi, Y.; Kurahashi, K. Subtilosin A, a new antibiotic peptide produced by Bacillus subtilis 168: Isolation, structural analysis, and biogenesis. J. Biochem. 1985, 98, 585–603. [Google Scholar] [CrossRef] [PubMed]
- Shelburne, C.E.; An, F.Y.; Dholpe, V.; Ramamoorthy, A.; Lopatin, D.E.; Lantz, M.S. The spectrum of antimicrobial activity of the bacteriocin subtilosin A. J. Antimicrob. Chemother. 2007, 59, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Stein, T.; Dusterhus, S.; Stroh, A.; Entian, K.D. Subtilosin production by two Bacillus subtilis subspecies and variance of the sbo-alb cluster. Appl. Environ. Microbiol. 2004, 70, 2349–2353. [Google Scholar] [CrossRef] [Green Version]
- Abriouel, H.; Franz, C.M.; Ben Omar, N.; Galvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [Green Version]
- Konuray, G.; Erginkaya, Z. Potential use of Bacillus coagulans in the food industry. Foods 2018, 7, 92. [Google Scholar] [CrossRef] [Green Version]
- Shinde, T.; Vemuri, R.; Shastri, M.D.; Perera, A.P.; Tristram, S.; Stanley, R.; Eri, R. Probiotic Bacillus coagulans MTCC 5856 spores exhibit excellent in-vitro functional efficacy in simulated gastric survival, mucosal adhesion and immunomodulation. J. Funct. Foods 2019, 52, 100–108. [Google Scholar] [CrossRef]
- Nath, S.; Chowdhury, S.; Dora, K. Application of Bacillus sp. as a biopreservative for food preservation. Int. J. Eng. Res. Appl. 2015, 5, 85–95. [Google Scholar]
- Riazi, S.; Dover, S.E.; Chikindas, M.L. Mode of action and safety of lactosporin, a novel antimicrobial protein produced by Bacillus coagulans ATCC 7050. J. Appl. Microbiol. 2012, 113, 714–722. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Wang, C.; Ruan, X.; Li, G.; Zhao, Y.; Wang, Y. Preservation of large yellow croaker (Pseudosciaena crocea) by Coagulin L1208, a novel bacteriocin produced by Bacillus coagulans L1208. Int J Food Microbiol. 2018, 266, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Abdhul, K.; Ganesh, M.; Shanmughapriya, S.; Vanithamani, S.; Kanagavel, M.; Anbarasu, K.; Natarajaseenivasan, K. Bacteriocinogenic potential of a probiotic strain Bacillus coagulans [BDU3] from Ngari. Int. J. Biol. Macromol. 2015, 79, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Orru, L.; Salvetti, E.; Cattivelli, L.; Lamontanara, A.; Michelotti, V.; Capozzi, V.; Spano, G.; Keller, D.; Cash, H.; Martina, A.; et al. Draft genome sequence of Bacillus coagulans GBI-30, 6086, a widely used spore-forming probiotic strain. Genome Announc. 2014, 2, e01080-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saroj, D.B.; Gupta, A.K. Genome based safety assessment for Bacillus coagulans strain LBSC (DSM 17654) for probiotic application. Int. J. Food Microbiol. 2020, 318, 108523. [Google Scholar] [CrossRef]
- Sreenadh, M.; Kumar, K.R.; Nath, S. In vitro evaluation of Weizmannia coagulans strain LMG S-31876 isolated from fermented rice for potential probiotic properties, safety assessment and technological properties. Life 2022, 12, 1388. [Google Scholar] [CrossRef]
- Aulitto, M.; Martinez-Alvarez, L.; Fiorentino, G.; Limauro, D.; Peng, X.; Contursi, P. A comparative analysis of Weizmannia coagulans genomes unravels the genetic potential for biotechnological applications. Int. J. Mol. Sci. 2022, 23, 3135. [Google Scholar] [CrossRef]
- Abdhul, K.; Ganesh, M.; Shanmughapriya, S.; Kanagavel, M.; Anbarasu, K.; Natarajaseenivasan, K. Antioxidant activity of exopolysaccharide from probiotic strain Enterococcus faecium (BDU7) from Ngari. Int. J. Biol. Macromol. 2014, 70, 450–454. [Google Scholar] [CrossRef]
- Voulgari, K.; Hatzikamari, M.; Delepoglou, A.; Georgakopoulos, P.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Antifungal activity of non-starter lactic acid bacteria isolates from dairy products. Food Control 2010, 21, 136–142. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, Y.; Xie, Q.; Zhang, Y.; Hu, J.; Li, P. Purification and characterization of plantaricin LPL-1, a novel class IIa bacteriocin produced by Lactobacillus plantarum LPL-1 isolated from fermented fish. Front. Microbiol. 2018, 9, 2276. [Google Scholar] [CrossRef]
- An, Y.; Wang, Y.; Liang, X.; Yi, H.; Zuo, Z.; Xu, X.; Zhang, D.; Yu, C.; Han, X. Purification and partial characterization of M1-UVs300, a novel bacteriocin produced by Lactobacillus plantarum isolated from fermented sausage. Food Control 2017, 81, 211–217. [Google Scholar] [CrossRef]
- Zhu, H.; Han, L.; Ni, Y.; Yu, Z.; Wang, D.; Zhou, J.; Li, B.; Zhang, W.; He, K. In vitro and in vivo antibacterial effects of Nisin against Streptococcus suis. Probiotics Antimicrob. Proteins 2021, 13, 598–610. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Teng, K.; Liu, Y.; Cao, Y.; Wang, T.; Ma, C.; Zhang, J.; Zhong, J. Bacteriocins: Potential for human health. Oxid. Med. Cell Longev. 2021, 2021, 5518825. [Google Scholar] [CrossRef]
- Johnson, E.M.; Jung, Y.G.; Jin, Y.Y.; Jayabalan, R.; Yang, S.H.; Suh, J.W. Bacteriocins as food preservatives: Challenges and emerging horizons. Crit. Rev. Food Sci. 2018, 58, 2743–2767. [Google Scholar] [CrossRef] [PubMed]
- Vijay Simha, B.; Sood, S.K.; Kumariya, R.; Garsa, A.K. Simple and rapid purification of pediocin PA-1 from Pediococcus pentosaceous NCDC 273 suitable for industrial application. Microbiol. Res. 2012, 167, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Kyriakou, P.K.; Ekblad, B.; Kristiansen, P.E.; Kaznessis, Y.N. Interactions of a class IIb bacteriocin with a model lipid bilayer, investigated through molecular dynamics simulations. Biochim. Biophys. Acta (BBA)—Biomembr. 2016, 1858, 824–835. [Google Scholar] [CrossRef]
- Chauhan, A.K.; Maheshwari, D.K.; Bajpai, V.K. Isolation and preliminary characterization of a bacteriocin-producer Bacillus strain inhibiting methicillin resistant Staphylococcus aureus. Acta Biol. Hung. 2017, 68, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Mercado, V.; Olmos, J. Bacteriocin production by Bacillus Species: Isolation, characterization, and application. Probiotics Antimicrob. Proteins 2022, 14, 1151–1169. [Google Scholar] [CrossRef]
- Kapse, N.G.; Engineer, A.S.; Gowdaman, V.; Wagh, S.; Dhakephalkar, P.K. Functional annotation of the genome unravels probiotic potential of Bacillus coagulans HS243. Genomics 2019, 111, 921–929. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Goker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Goker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Richter, M.; Rossello-Mora, R.; Oliver Glockner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Gueimonde, M.; Sanchez, B.; de Los Reyes-Gavilan, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Ruiz, L.; Ruas-Madiedo, P.; Gueimonde, M.; de los Reyes-Gavilan, C.G.; Margolles, A.; Sanchez, B. How do bifidobacteria counteract environmental challenges? Mechanisms involved and physiological consequences. Genes Nutr. 2011, 6, 307–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padan, E.; Venturi, M.; Gerchman, Y.; Dover, N. Na+/H+ antiporters. Biochim. Biophys. Acta 2001, 1505, 144–157. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, S.; Garcia-Cabado, A.; Yu, F.; Teter, K.; Lukacs, G.; Skorecki, K.; Moore, H.P.; Orlowski, J.; Grinstein, S. The epithelial sodium-hydrogen antiporter Na+/H+ exchanger 3 accumulates and is functional in recycling endosomes. J. Biol. Chem. 1998, 273, 2035–2043. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, L.; Margolles, A.; Sanchez, B. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [Green Version]
- Granato, D.; Bergonzelli, G.E.; Pridmore, R.D.; Marvin, L.; Rouvet, M.; Corthesy-Theulaz, I.E. Cell surface-associated elongation factor Tu mediates the attachment of Lactobacillus johnsonii NCC533 (La1) to human intestinal cells and mucins. Infect. Immun. 2004, 72, 2160–2169. [Google Scholar] [CrossRef] [Green Version]
- Khatri, I.; Sharma, S.; Ramya, T.N.; Subramanian, S. Complete genomes of Bacillus coagulans S-lac and Bacillus subtilis TO-A JPC, two phylogenetically distinct probiotics. PLoS ONE 2016, 11, e0156745. [Google Scholar] [CrossRef] [Green Version]
- Haiko, J.; Westerlund-Wikstrom, B. The role of the bacterial flagellum in adhesion and virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef] [PubMed]
- Henderson, B.; Nair, S.; Pallas, J.; Williams, M.A. Fibronectin: A multidomain host adhesin targeted by bacterial fibronectin-binding proteins. FEMS Microbiol. Rev. 2011, 35, 147–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadimitriou, K.; Zoumpopoulou, G.; Foligne, B.; Alexandraki, V.; Kazou, M.; Pot, B.; Tsakalidou, E. Discovering probiotic microorganisms: In vitro, in vivo, genetic and omics approaches. Front. Microbiol. 2015, 6, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemperman, R.; Kuipers, A.; Karsens, H.; Nauta, A.; Kuipers, O.; Kok, J. Identification and characterization of two novel clostridial bacteriocins, circularin A and closticin 574. Appl. Environ. Microbiol. 2003, 69, 1589–1597. [Google Scholar] [CrossRef] [Green Version]
- Scholz, R.; Vater, J.; Budiharjo, A.; Wang, Z.; He, Y.; Dietel, K.; Schwecke, T.; Herfort, S.; Lasch, P.; Borriss, R. Amylocyclicin, a novel circular bacteriocin produced by Bacillus amyloliquefaciens FZB42. J. Bacteriol. 2014, 196, 1842–1852. [Google Scholar] [CrossRef] [Green Version]
- Havarstein, L.S.; Diep, D.B.; Nes, I.F. A family of bacteriocin Abc transporters carry out proteolytic processing of their substrates concomitant with export. Mol. Microbiol. 1995, 16, 229–240. [Google Scholar] [CrossRef]
- Johny, L.C.; Suresh, P.V. Complete genome sequencing and strain characterization of a novel marine Bacillus velezensis FTL7 with a potential broad inhibitory spectrum against foodborne pathogens. World J. Microbiol. Biotechnol. 2022, 38, 164. [Google Scholar] [CrossRef]
- Teng, K.; Zhang, J.; Zhang, X.; Ge, X.; Gao, Y.; Wang, J.; Lin, Y.; Zhong, J. Identification of ligand specificity determinants in Lantibiotic bovicin HJ50 and the receptor BovK, a multitransmembrane histidine kinase. J. Biol. Chem. 2014, 289, 9823–9832. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicator Strain | Source | Media | Activity (mm) a | MIC (µg/mL) | MBC (µg/mL) |
|---|---|---|---|---|---|
| Gram-positive bacteria | |||||
| Listeria monocytogenes CMCC 54,004 | Lab | TSYEB | +++ | 55.32 | 221.28 |
| Staphylococcus aureus ATCC 25,923 | Lab | TSB | + | 221.28 | 442.56 |
| Bacillus cereus ATCC 14,579 | Lab | LB | + | 221.28 | 442.56 |
| Bacillus subtilis | Lab | LB | ++ | 110.64 | 442.56 |
| Bacillus licheniformis | Lab | LB | +++ | 55.32 | 442.56 |
| Bacillus amyloliquefaciens | Lab | LB | + | 221.28 | 885.13 |
| Lactobacillus plantarum | Lab | MRS | +++ | 55.32 | 221.28 |
| lactococcus lactis MG1363 | Lab | MRS | - | ||
| Enterococcus faecalis | Lab | MRS | - | ||
| Lactobacillus bulgaricus | Lab | MRS | - | ||
| Lactococcus lactis NZ9000 | Lab | MRS | - | ||
| Gram-negative bacteria | |||||
| Escherichia coli BL21 | Lab | LB | - | ||
| Escherichia coli BW25113 | Lab | LB | - | ||
| Pseudomonas aeruginosa | Lab | LB | + | 442.56 | 885.13 |
| Attributes | W. coagulans PL-W |
|---|---|
| Genome size (bp) | 3,666,052 |
| No. of contigs | 2 |
| GC content % | 46.24 |
| Coding DNA sequence (CDS) | 3485 |
| rRNAs | 30 |
| tRNAs | 84 |
| COG Class | Name | Count | Proportion (%) |
|---|---|---|---|
| C | Energy production and conversion | 143 | 4.81 |
| D | Cell cycle control, cell division, chromosome partitioning | 180 | 6.05 |
| E | Amino acid transport and metabolism | 265 | 8.90 |
| F | Nucleotide transport and metabolism | 96 | 3.23 |
| G | Carbohydrate transport and metabolism | 254 | 8.53 |
| H | Coenzyme transport and metabolism | 156 | 5.24 |
| I | Lipid transport and metabolism | 169 | 5.68 |
| J | Translation, ribosomal structure, and biogenesis | 212 | 7.12 |
| K | Transcription | 221 | 7.43 |
| L | Replication, recombination, and repair | 127 | 4.27 |
| M | Cell wall/membrane/envelope biogenesis | 127 | 4.27 |
| N | Cell motility | 49 | 1.65 |
| O | Post-translational modification, protein turnover, chaperones | 119 | 4.00 |
| P | Inorganic ion transport and metabolism | 133 | 4.47 |
| Q | Secondary metabolite biosynthesis, transport and catabolism | 41 | 1.38 |
| R | General function prediction only | 184 | 6.18 |
| S | Function unknown | 122 | 4.10 |
| T | Signal transduction mechanisms | 165 | 5.54 |
| U | Intracellular trafficking, secretion, and vesicular transport | 29 | 0.97 |
| V | Defense mechanisms | 94 | 3.16 |
| W | Extracellular structures | 9 | 0.30 |
| X | Mobilome: prophages and transposons | 75 | 2.52 |
| Z | Cytoskeleton | 6 | 0.20 |
| Antibiotics | MIC (µg/mL) | MIC Cut-off Values (µg/mL) | Interpretation |
|---|---|---|---|
| Clindamycin | 0.125 | 4 | S |
| Gentamicin | 0.25 | 4 | S |
| Streptomycin | 0.25 | 8 | S |
| Vancomycin | 0.5 | 4 | S |
| Erythromycin | 2 | 4 | S |
| Kanamycin | 0.25 | 8 | S |
| Tetracycline | 0.25 | 8 | S |
| Chloramphenicol | 4 | 8 | S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Gu, Z.; Zhang, S.; Li, P. Complete Genome Sequencing Revealed the Potential Application of a Novel Weizmannia coagulans PL-W Production with Promising Bacteriocins in Food Preservative. Foods 2023, 12, 216. https://doi.org/10.3390/foods12010216
Wang Y, Gu Z, Zhang S, Li P. Complete Genome Sequencing Revealed the Potential Application of a Novel Weizmannia coagulans PL-W Production with Promising Bacteriocins in Food Preservative. Foods. 2023; 12(1):216. https://doi.org/10.3390/foods12010216
Chicago/Turabian StyleWang, Yu, Zelin Gu, Shiqi Zhang, and Pinglan Li. 2023. "Complete Genome Sequencing Revealed the Potential Application of a Novel Weizmannia coagulans PL-W Production with Promising Bacteriocins in Food Preservative" Foods 12, no. 1: 216. https://doi.org/10.3390/foods12010216
APA StyleWang, Y., Gu, Z., Zhang, S., & Li, P. (2023). Complete Genome Sequencing Revealed the Potential Application of a Novel Weizmannia coagulans PL-W Production with Promising Bacteriocins in Food Preservative. Foods, 12(1), 216. https://doi.org/10.3390/foods12010216