
Microbiological Characterization of Protected Designation of Origin Serra da Estrela Cheese

 ,
,  ,
,  , and
, and 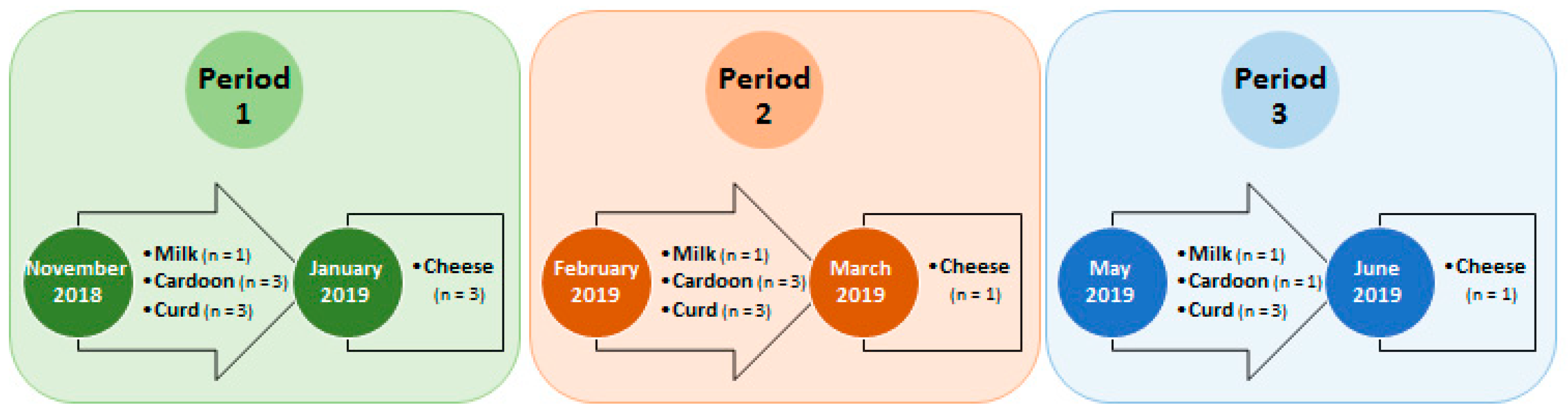{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Strategy and Cheese Manufacture
2.2. Microbial Characterization Analysis
2.3. DNA Extraction and Genetic Identification of LAB Isolates
2.4. Statistical Analysis
3. Results and Discussion
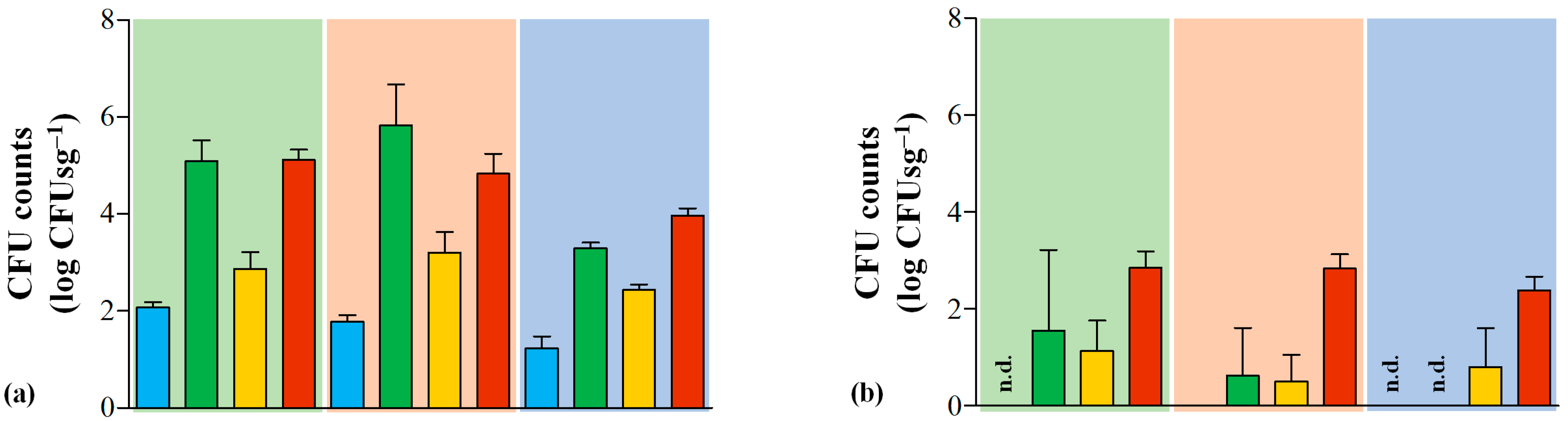
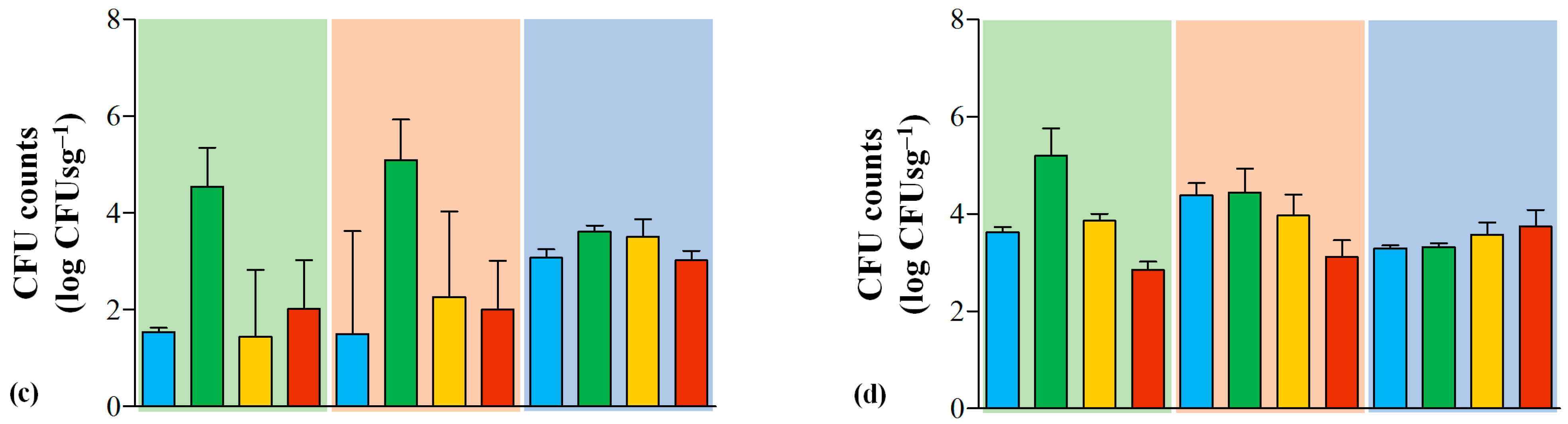
3.1. Hygiene and Safety of Serra da Estrela PDO Cheeses
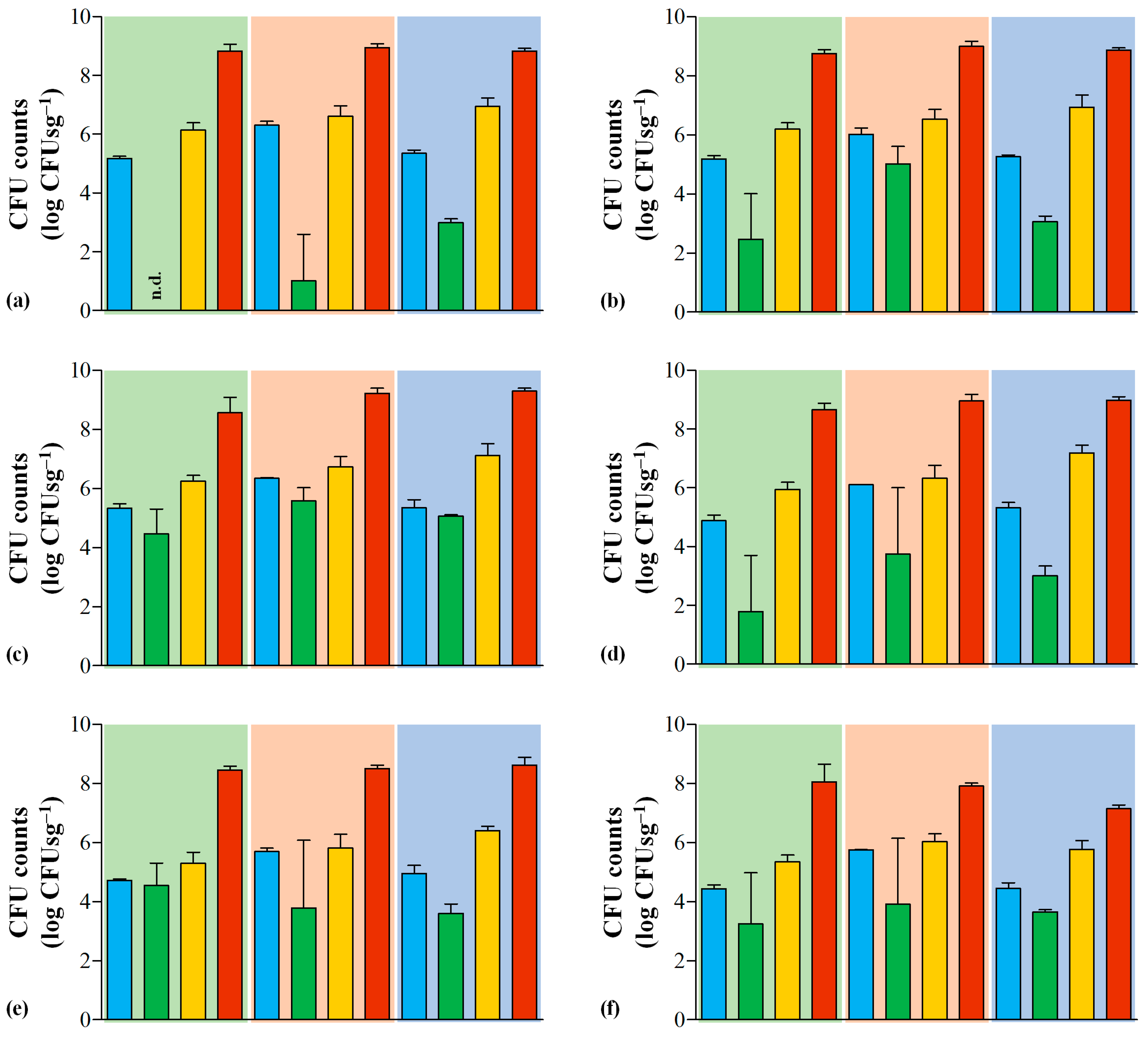
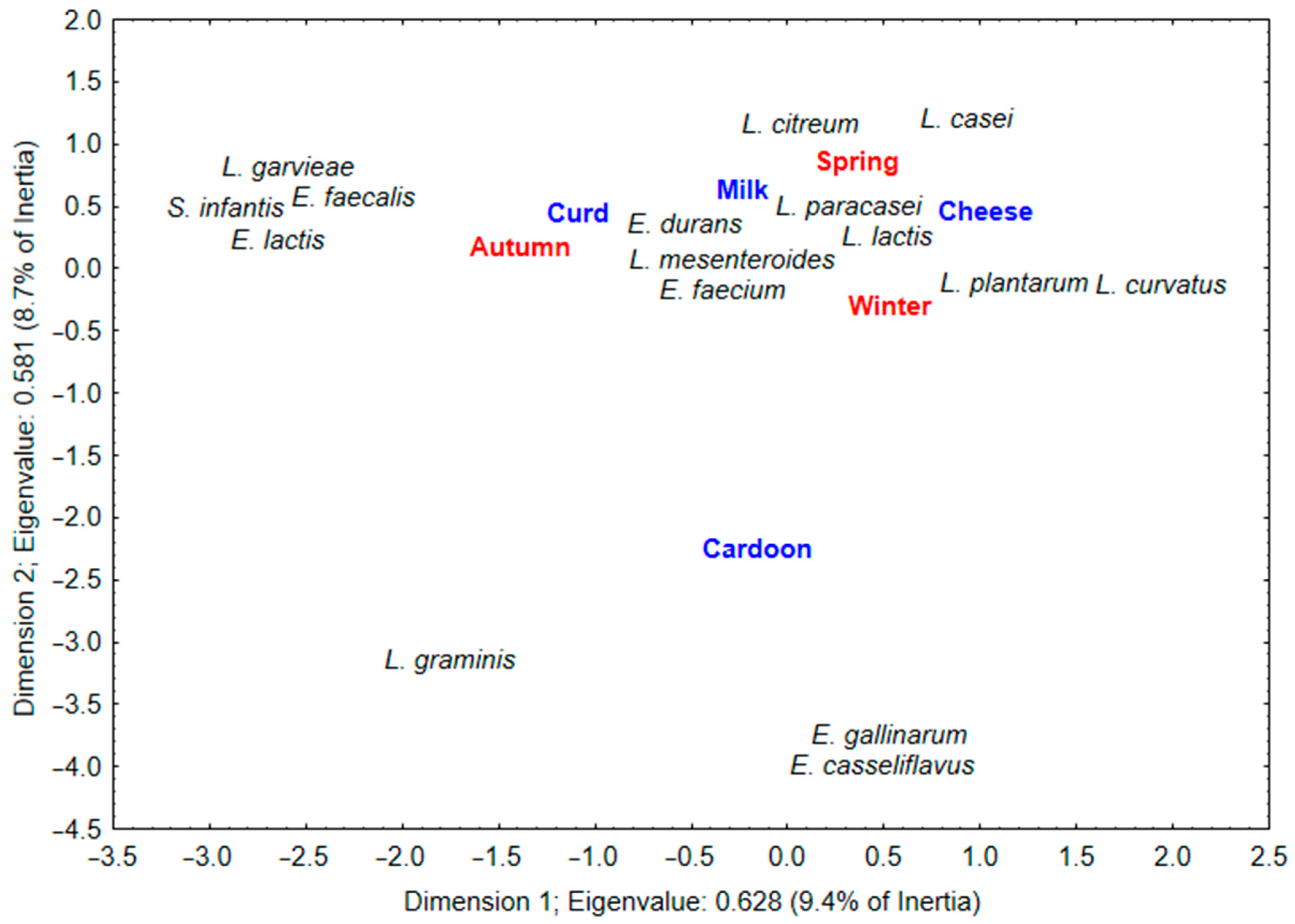
3.2. LAB Microbiota in Serra da Estrela PDO Cheeses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fox, P.F.; McSweeney, P. Cheese: An Overview. In Cheese−Chemistry, Physics and Microbiology, 4th ed.; Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 5–21. [Google Scholar] [CrossRef]
- Jaros, D.; Rohm, H. Rennets: Applied Aspects. In Cheese−Chemistry, Physics and Microbiology, 4th ed.; Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 53–67. [Google Scholar] [CrossRef]
- Zheng, X.; Shi, X.; Wang, B. A Review on the General Cheese Processing Technology, Flavor Biochemical Pathways and the Influence of Yeasts in Cheese. Front. Microbiol. 2021, 12, 703284. [Google Scholar] [CrossRef]
- Kilcawley, K.; O’Sullivan, M. Cheese Flavour Development and Sensory Characteristics. In Global Cheesemaking Technology: Cheese Quality and Characteristics, 1st ed.; Papademas, P., Bintsis, T., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 45–70. [Google Scholar] [CrossRef]
- McSweeney, P.; Ottogalli, G.; Fox, P.F. Diversity and Classification of Cheese Varieties: An Overview. In Cheese−Chemistry, Physics and Microbiology, 4th ed.; Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 781–808. [Google Scholar] [CrossRef]
- Irlinger, F.; Layec, S.; Hélinck, S.; Dugat-Bony, E. Cheese rind microbial communities: Diversity, composition and origin. FEMS Microbiol. Lett. 2015, 362, 1–11. [Google Scholar] [CrossRef]
- Jonnala, B.R.; McSweeney, P.; Sheehan, J.J.; Cotter, P.D. Sequencing of the Cheese Microbiome and Its Relevance to Industry. Front. Microbiol. 2018, 9, 1020. [Google Scholar] [CrossRef]
- Montel, M.C.; Buchin, S.; Mallet, A.; Delbes−Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef]
- Cotter, P.D.; Beresford, T.P. Microbiome Changes During Ripening. In Cheese−Chemistry, Physics and Microbiology, 4th ed.; Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 389–409. [Google Scholar] [CrossRef]
- Irlinger, F.; Helinck, S.; Jany, J.L. Secondary and Adjunct Cultures. In Cheese−Chemistry, Physics and Microbiology, 4th ed.; Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 273–300. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The complex microbiota of raw milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef]
- O’Sullivan, O.; Cotter, P.D. Microbiota of Raw Milk and Raw Milk Cheeses. In Cheese−Chemistry, Physics and Microbiology, 4th ed.; McSweeney, P., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 301–316. [Google Scholar] [CrossRef]
- Fuka, M.M.; Wallisch, S.; Engel, M.; Welzl, G.; Havranek, J.; Schloter, M. Dynamics of bacterial communities during the ripening process of different Croatian cheese types derived from raw ewe’s milk cheeses. PLoS ONE 2013, 8, e80734. [Google Scholar] [CrossRef]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L.H. Pathogens in Cheese and Foodborne Illnesses. In Fundamentals of Cheese Science; Fox, P.F., Guinee, T.P., Cogan, T.M., McSweeney, P.L.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 681–713. [Google Scholar] [CrossRef]
- Picon, A. Cheese Microbial Ecology and Safety. In Global Cheesemaking Technology: Cheese Quality and Characteristics, 1st ed.; Papademas, P., Bintsis, T., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 71–99. [Google Scholar] [CrossRef]
- Verraes, C.; Vlaemynck, G.; Van Weyenberg, S.; De Zutter, L.; Daube, G.; Sindic, M.; Uyttendaele, M.; Herman, L. A review of the microbiological hazards of dairy products made from raw milk. Int. Dairy J. 2015, 50, 32–44. [Google Scholar] [CrossRef]
- Morales, M.; Bintsis, T.; Alichanidis, E.; Herian, K.; Jelen, P.; Hynes, E.R.; Perotti, M.C.; Bergamini, C.V.; Attard, E.; Grupetta, A.; et al. Soft cheeses (with rennet). In Global Cheesemaking Technology; Papademas, P., Bintsis, T., Eds.; Jonh Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 301–325. [Google Scholar] [CrossRef]
- Inácio, R.S.; Gomes, A.; Saraiva, J.A. Serra da Estrela cheese: A review. J. Food Process. Preserv. 2000, 44, e14412. [Google Scholar] [CrossRef]
- Macedo, A.C.; Malcata, F.X.; Oliveira, J.C. The technology, Chemistry and Microbiology of Serra Cheese: A review. J. Dairy Sci. 1993, 76, 1725–1739. [Google Scholar] [CrossRef]
- Macedo, A.C.; Malcata, F.X.; Hogg, T.A. Microbiological profile in Serra ewes’ cheese during ripening. J. Appl. Bacteriol. 1995, 79, 1–11. [Google Scholar] [CrossRef]
- Macedo, A.C.; Costa, M.L.; Malcata, F.X. Changes in the Microflora of Serra Cheese Evolution Throughout Ripening Time, Lactation Period and Axial Location. Int. Dairy J. 1996, 6, 79–94. [Google Scholar] [CrossRef]
- Queijo Serra da Estrela–Denominação de Origem Protegida-Caderno de Especificações. 2011. Available online: https://tradicional.dgadr.gov.pt/images/prod_imagens/queijos/docs/CE_Queijo_Serra.pdf.pdf (accessed on 5 February 2023).
- Rocha, R.; Vaz Velho, M.; Santos, J.; Fernandes, P. Serra da Estrela PDO Cheese Microbiome as Revealed by Next Generation Sequencing. Microorganisms 2021, 9, 2007. [Google Scholar] [CrossRef]
- ISO 16649-2:2001; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Beta-Glucuronidase-Positive Escherichia coli—Part 2: Colony-Count Technique at 44 Degrees C Using 5-bromo-4-chloro-3-indolyl beta-D-glucuronide. ISO: Geneva, Switzerland, 2001. Available online: https://www.iso.org/standard/29824.html (accessed on 10 September 2018).
- ISO 21528-2:2017; Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Enterobacteriaceae—Part 2: Colony-Count Technique. ISO: Geneva, Switzerland, 2017. Available online: https://www.iso.org/standard/63504.html (accessed on 10 September 2018).
- ISO 7932:2004; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Presumptive Bacillus cereus—Colony-Count Technique at 30 Degrees C. ISO: Geneva, Switzerland, 2004. Available online: https://www.iso.org/standard/38219.html (accessed on 10 September 2018).
- ISO 6888-1:1999; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Coagulase-Positive Staphylococci (Staphylococcus aureus and Other Species)—Part 1: Technique Using Baird-Parker Agar Medium. ISO: Geneva, Switzerland, 1999.
- ISO 11290-2:2017; Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria monocytogenes and of Listeria spp.—Part 2: Enumeration Method. ISO: Geneva, Switzerland, 2017. Available online: https://www.iso.org/standard/60314.html (accessed on 10 September 2018).
- ISO 21527-1:2008; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Yeasts and moulds—Part 1: Colony Count Technique in Products with Water Activity Greater Than 0.95. ISO: Geneva, Switzerland, 2008. Available online: https://www.iso.org/standard/38275.html (accessed on 10 September 2018).
- ISO 6579-1:2017; Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp. ISO: Geneva, Switzerland, 2017. Available online: https://www.iso.org/standard/56712.html (accessed on 10 September 2018).
- ISO 7937:2004; Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Clostridium perfringens—Colony-Count Technique. ISO: Geneva, Switzerland, 2004. Available online: https://www.iso.org/standard/36588.html (accessed on 10 September 2018).
- Alves, S.; Teixeira, L.; Ribeiro, O.; Paul, C. Examining Frailty Phenotype Dimensions in the Oldest Old. Front. Psychol. 2020, 11, 434. [Google Scholar] [CrossRef]
- Xia, Y. Correlation and association analyses in microbiome study integrating multiomics in health and disease. In Progress in Molecular Biology and Translational Science; Sun, J., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 309–491. [Google Scholar] [CrossRef]
- Langer, A.J.; Ayers, T.; Grass, J.; Lynch, M.; Angulo, F.J.; Mahon, B.E. Nonpasteurized Dairy Products, Disease Outbreaks, and State Laws-United States, 1993–2006. Emerg. Infect. Dis. 2012, 18, 385–391. [Google Scholar] [CrossRef]
- Mungai, E.A.; Behravesh, C.; Gould, L. Increased Outbreaks Associated with Nonpasteurized Milk, United States, 2007–2012. Emerg. Infect. Dis. 2015, 21, 119–122. [Google Scholar] [CrossRef]
- Adams, N.; Byrne, L.; Edge, J.; Hoban, A.; Jenkins, C.; Larkin, L. Gastrointestinal infections caused by consumption of raw drinking milk in England & Wales, 1992–2017. Epidemiol. and Infect. 2019, 147, e281. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs, Consolidated Version of 08/03/2020. Available online: http://data.europa.eu/eli/reg/2005/2073/oj (accessed on 21 January 2021).
- Instituto Nacional de Saúde Doutor Ricardo Jorge. Interpretação de Resultados de Ensaios Microbiológicos em Alimentos Prontos para Consumo e em Superfícies do Ambiente de Preparação e Distribuição Alimentar: Valores-guia. Lisboa: INSA IP, 2019. Available online: https://www.insa.min-saude.pt/wp-content/uploads/2019/12/INSA_Valores-guia.pdf (accessed on 21 January 2021).
- Tavaria, F.K.; Malcata, F.X. Microbiological Characterization of Serra da Estrela Cheese throughout Its Appellation d’Origine Protégée Region. J. Food Prot. 1998, 61, 601–607. [Google Scholar] [CrossRef]
- Tavaria, F.K.; Malcata, F.X. On the microbiology of Serra da Estrela cheese: Geographical and chronological considerations. Food Microbiol. 2000, 17, 293–304. [Google Scholar] [CrossRef]
- Gaya, P.; Medina, M.; Nuñez, M. Enterobacteriaceae, coliforms, faecal coliforms and salmonellas in raw ewes’ milk. J. Appl. Bacteriol. 1987, 62, 321–326. [Google Scholar] [CrossRef]
- Fernández-Salguero, J.; Sánchez, E.; Gómez, R.; Mata, C.; Vioque, M.; Tejada, L. Preliminary study of microbiological quality of cardoons of the genus Cynara L. used in manufacture of traditional cheeses. Milchwissenschaft 1999, 54, 688–689. [Google Scholar]
- Gómez, R.; Sánchez, E.; Vioque, M.; Ferreira, J.; Tejada, L.; Fernández-Salguero, J. Microbiological characteristics of ewes’ milk cheese manufactured using aqueous extracts of flowers from various species of cardoon Cynara L. Milchwissenschaft 2001, 56, 16–19. [Google Scholar]
- Parente, E.; Cogan, T.M.; Powell, I.B. Starter Cultures: General Aspects. In Cheese−Chemistry, Physics and Microbiology, 4th ed.; Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 201–226. [Google Scholar] [CrossRef]
- Dahl, S.; Tavaria, F.K.; Malcata, F.X. Relationships between flavor and microbiological profiles in Serra da Estrela cheese throughout ripening. Int. Dairy J. 2000, 10, 255–262. [Google Scholar] [CrossRef]
- Byappanahalli, M.N.; Nevers, M.B.; Korajkic, A.; Staley, Z.R.; Harwood, V.J. Enterococci in the Environment. Microbiol. Mol. Biol. Rev. 2012, 76, 685–706. [Google Scholar] [CrossRef]
- Ramette, A. Multivariate analyses in microbial ecology. FEMS Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [PubMed]
- Greenacre, M. Correspondence analysis in medical research. Stat. Methods Med. Res. 1992, 1, 97–117. [Google Scholar] [CrossRef] [PubMed]
- Esteves, A.; Aymerich, T.; Garriga, M.; Patarata, L.; Fontes, M.C.; Martins, C. Tracing Salmonella in Alheira processing plants. J. Appl. Microbiol. 2007, 103, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.P.; Zhang, Y.X.; Yuan, Y.H.; Dai, L.; Yue, T.L. Characteristic fruit wine production via reciprocal selection of juice and non-Saccharomyces species. Food Microbiol. 2019, 79, 66–74. [Google Scholar] [CrossRef]
- Yap, S.K.; Chin, N.L.; Shah, N.; Than, L.T.L.; Niranjan, K. Physicochemical characteristics, antioxidant properties and bacterial profiling of three Malaysian honey varieties: A study using multivariate analysis. J. Sci. Food Agric. 2022, 102, 5440–5451. [Google Scholar] [CrossRef] [PubMed]
- Macedo, A.C.; Malcata, F.X. Role of adventitious microflora in proteolysis and lipolysis of Serra cheese: Preliminary screening. Z. Lebensm. Unters. Forsch. 1997, 205, 25–30. [Google Scholar] [CrossRef]
- Tavaria, F.K.; Ferreira, A.C.; Malcata, F.X. Volatile Free Fatty Acids as Ripening Indicators for Serra da Estrela Cheese. J. Dairy Sci. 2004, 87, 4064–4072. [Google Scholar] [CrossRef]
- Franciosi, E.; Settanni, L.; Cavazza, A.; Poznanski, E. Presence of enterococci in raw cow’s milk and “Puzzone Di Moena” cheese. J. Food Proc. Pres. 2009, 33, 204–217. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, R.; Couto, N.; Pinto, R.P.; Vaz-Velho, M.; Fernandes, P.; Santos, J. Microbiological Characterization of Protected Designation of Origin Serra da Estrela Cheese. Foods 2023, 12, 2008. https://doi.org/10.3390/foods12102008
Rocha R, Couto N, Pinto RP, Vaz-Velho M, Fernandes P, Santos J. Microbiological Characterization of Protected Designation of Origin Serra da Estrela Cheese. Foods. 2023; 12(10):2008. https://doi.org/10.3390/foods12102008
Chicago/Turabian StyleRocha, Rui, Nélson Couto, Ricardo Pereira Pinto, Manuela Vaz-Velho, Paulo Fernandes, and Joana Santos. 2023. "Microbiological Characterization of Protected Designation of Origin Serra da Estrela Cheese" Foods 12, no. 10: 2008. https://doi.org/10.3390/foods12102008
APA StyleRocha, R., Couto, N., Pinto, R. P., Vaz-Velho, M., Fernandes, P., & Santos, J. (2023). Microbiological Characterization of Protected Designation of Origin Serra da Estrela Cheese. Foods, 12(10), 2008. https://doi.org/10.3390/foods12102008