
Optimization and Molecular Mechanism of Novel α-Glucosidase Inhibitory Peptides Derived from Camellia Seed Cake through Enzymatic Hydrolysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of CSCP
2.3. Preparation of CSCPH
2.4. Optimization of Enzymatic Parameters via RSM
2.5. Fractionation and Purification of α-Glucosidase Inhibitory Peptides
2.5.1. Ultrafiltration
2.5.2. Sephadex G25 Chromatography
2.6. Assaying the Inhibitory Action of α-Glucosidase In Vitro
2.7. O-Phthaladehyde (OPA) Assay of Hydrolysis Degree (DH) of CSCPH
2.8. Examination of Amino Acids
2.9. Identification of the α-Glucosidase Inhibitory Peptides
2.10. Virtual Screening
2.11. Synthesis of α-Glucosidase Inhibitory Peptides
2.12. Molecular Docking
2.13. Mechanism of α-Glucosidase Inhibitions
2.14. Simulated Gastrointestinal Digestion In Vitro
2.15. Statistical Analysis
3. Results and Discussion
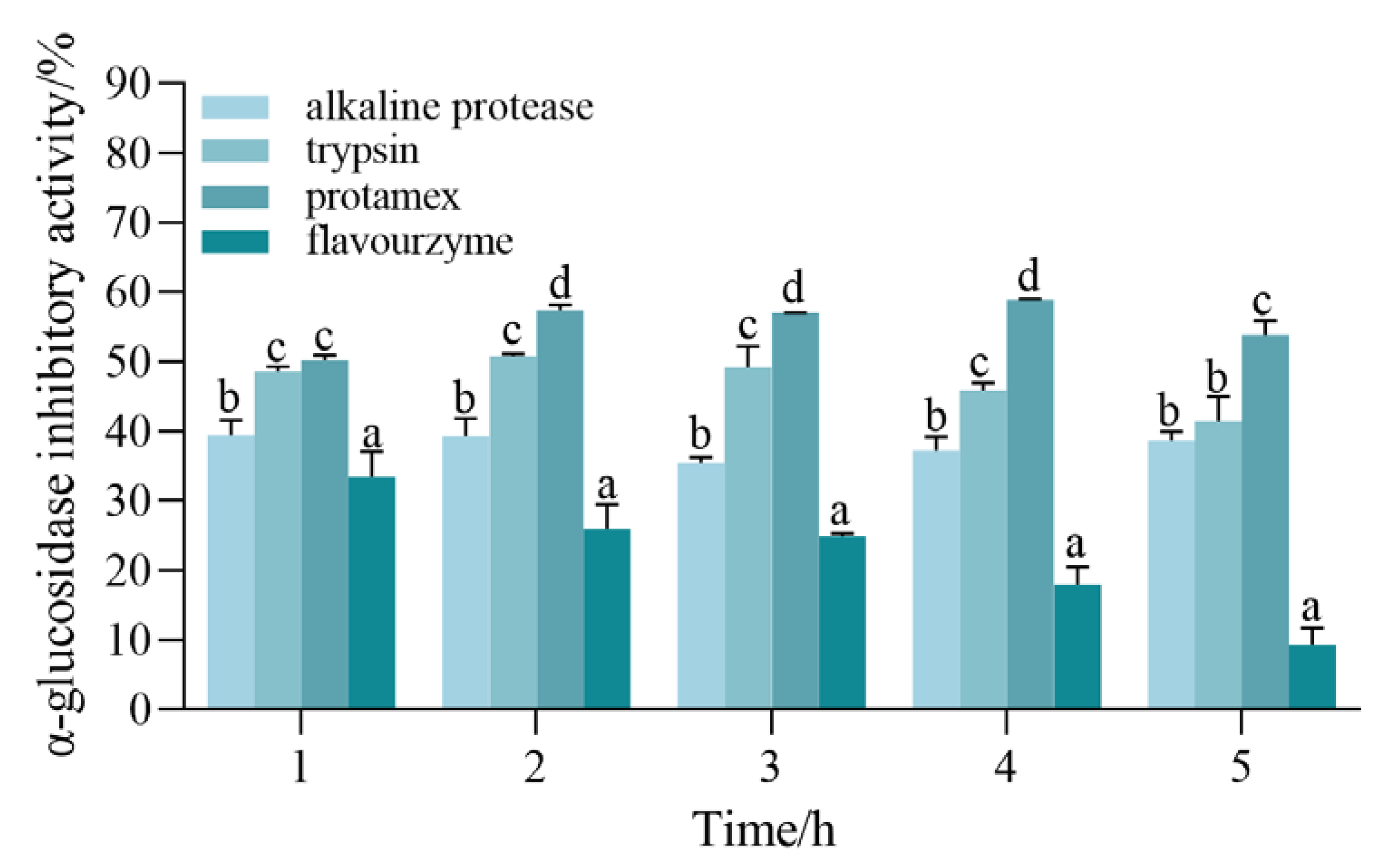
3.1. Protease Screening
3.2. Optimization of CSCPH
3.2.1. Preliminary Assessment
3.2.2. Optimization Analysis of RSM
0.2775AD − 0.0675BC + 0.1775BD + 0.355CD − 1.95A2 − 6.2B2 − 2.66C2 −
10.62D2
3.3. Separation and Purification
3.3.1. Ultrafiltration
3.3.2. Separation of α-Glucosidase Inhibitory Peptides by Gel Filtration Chromatography
3.4. Amino Acid Analysis
3.5. Screening for Peptides with α-Glucosidase Inhibitory Activity
3.6. Mechanism of α-Glucosidase Inhibition
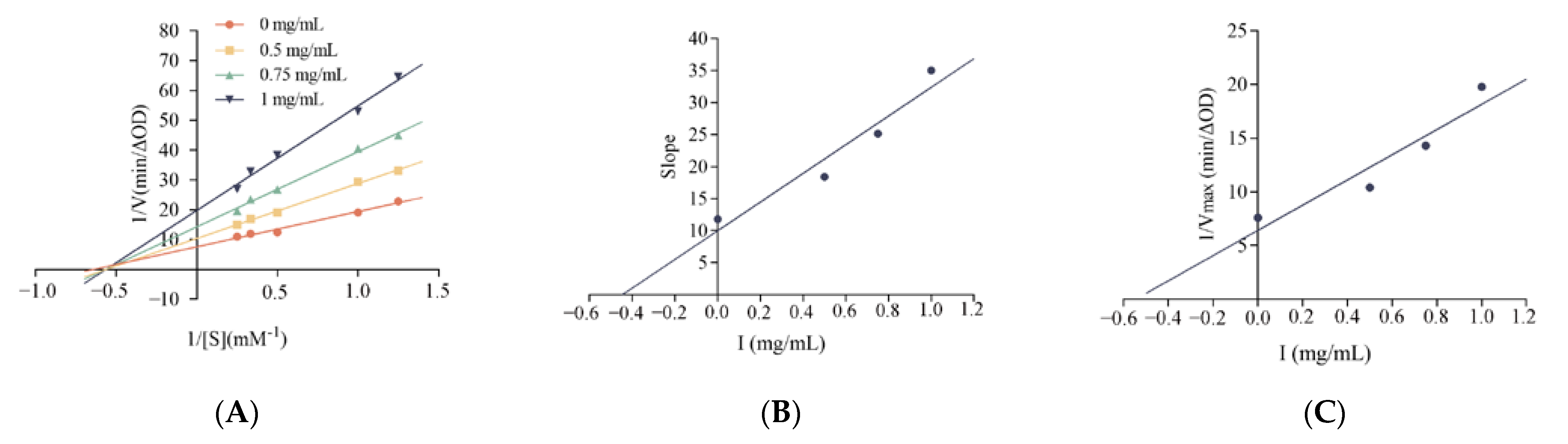
3.6.1. Kinetics of α-Glucosidase Inhibition
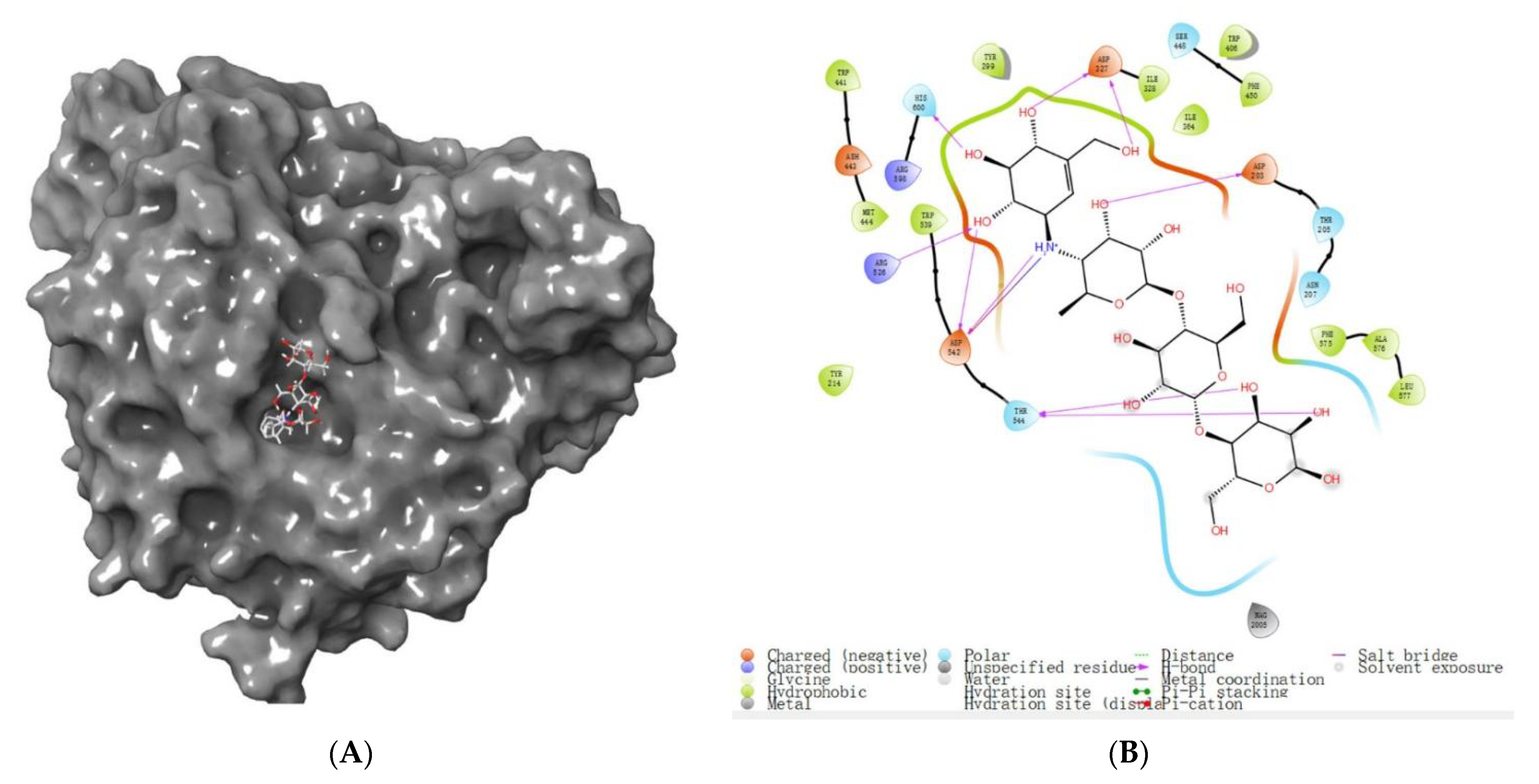
3.6.2. Molecular Docking
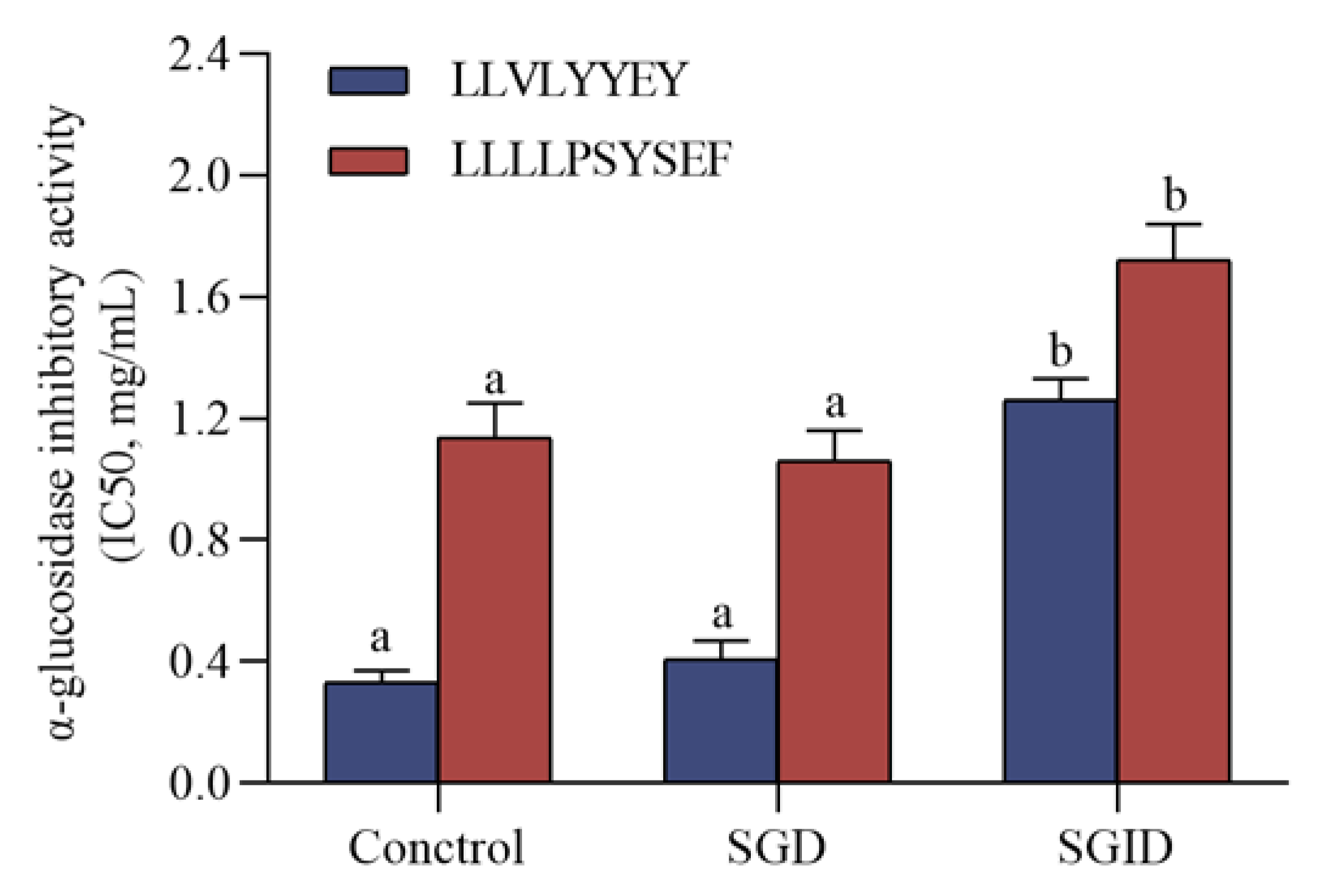
3.7. α-Glucosidase Inhibitory Peptides Simulated Gastrointestinal Digestion In Vitro
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gorelick, J.; Kitron, A.; Pen, S.; Rosenzweig, T.; Madar, Z. Anti-diabetic activity of chiliadenus iphionoides. J. Ethnopharmacol. 2012, 137, 1245–1249. [Google Scholar] [CrossRef]
- Liu, W.W.; Li, H.Y.; Wen, Y.Y.; Liu, Y.L.; Wang, J.; Sun, B.G. Molecular mechanism for the α-glucosidase inhibitory effect of wheat germ peptides. J. Agric. Food Chem. 2021, 69, 15231–15239. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, D.; Malisova, S.; Lindberg, F.A.; Karaniki, G. Glycemic index (GI) or glycemic load (GL) and dietary interventions for optimizing postprandial hyperglycemia in patients with T2 diabetes: A Review. Nutrients 2020, 12, 1561. [Google Scholar] [CrossRef] [PubMed]
- Brand-Miller, J.; Buyken, A.E. The relationship between glycemic index and health. Nutrients 2020, 12, 536. [Google Scholar] [CrossRef] [Green Version]
- Nong, N.T.P.; Hsu, J.L. Characteristics of food protein-derived antidiabetic bioactive peptides: A literature update. Int. J. Mol. Sci. 2021, 22, 9508. [Google Scholar] [CrossRef] [PubMed]
- Liu, I.M.; Tzeng, T.F.; Liou, S.S.; Chang, C.J. Angelica acutiloba root alleviates advanced glycation end-product-mediated renal injury in streptozotocin-diabetic rats. J. Food Sci. 2011, 76, H165–H174. [Google Scholar] [CrossRef]
- Jin, X.C.; Ning, Y. Antioxidant and antitumor activities of the polysaccharide from seed cake of camellia oleifera abel. Int. J. Biol. Macromol. 2012, 51, 364–368. [Google Scholar] [CrossRef]
- Tai, Y.L.; Wei, C.L.; Yang, H.; Zhang, L.; Chen, Q.; Deng, W.W.; Wei, S.; Zhang, J.; Fang, C.B.; Ho, C.T. Transcriptomic and phytochemical analysis of the biosynthesis of characteristic constituents in tea (Camellia sinensis) compared with oil tea (Camellia oleifera). BMC Plant Biol. 2015, 15, 150. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.P.; Yen, G.C. Antioxidant activity and bioactive compounds of tea seed (Camellia oleifera Abel.) oil. J. Agric. Food Chem. 2006, 54, 779–784. [Google Scholar] [CrossRef]
- Feng, J.; Ma, Y.L.; Sun, P.; Thakur, K.; Wang, S.Y.; Zhang, J.G.; Wei, Z.J. Purification and characterisation of α-glucosidase inhibitory peptides from defatted camellia seed cake. Int. J. Food Sci. Technol. 2020, 56, 138–147. [Google Scholar] [CrossRef]
- Wang, J.; Wu, T.; Fang, L.; Liu, C.L.; Liu, X.T.; Li, H.M.; Shi, J.H.; Li, M.H.; Min, W.H. Anti-diabetic effect by walnut (Juglans mandshurica Maxim.)-derived peptide LPLLR through inhibiting α-glucosidase and α-amylase, and alleviating insulin resistance of hepatic HepG2 cells. J. Funct. Foods 2020, 69, 103944. [Google Scholar] [CrossRef]
- Mahmoudreza, O.; Abdolmohammad, A.K.; Ali, M.; Rajab, M.N. Optimization of enzymatic hydrolysis of visceral waste proteins of yellowfin tuna (Thunnus albacares). Food Bioprocess Technol. 2012, 5, 696–705. [Google Scholar] [CrossRef]
- Keskin Gündoğdu, T.; Deniz, İ.; Çalışkan, G.; Şahin, E.S.; Azbar, N. Experimental design methods for bioengineering applications. Crit. Rev. Biotechnol. 2016, 36, 368–388. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.Z.; He, Z.P.; Wang, Q.Q.; Wu, F.H.; Liu, X.Q. Response surface optimization of enzymatic hydrolysis of peptides of chinese pecan (Carya cathayensis) and analysis of their antioxidant capacities and structures. Int. J. Pept. Res. Ther. 2021, 27, 1239–1251. [Google Scholar] [CrossRef]
- Yao, H.L.; Yang, J.N.; Zhan, J.J.; Lu, Q.; Su, M.; Jiang, Y.J. Preparation, amino acid composition, and in vitro antioxidant activity of okra seed meal protein hydrolysates. Food Sci. Nutr. 2021, 9, 3059–3070. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Gao, T.; Hou, Y.K.; Li, D.; Fu, L. Identification and characterization of two novel α-glucosidase inhibitory peptides from almond (Armeniaca sibirica) oil manufacture residue. LWT 2020, 134, 110215. [Google Scholar] [CrossRef]
- Karthiraj, T.; Harish Babu, B.; Senthil Kumar, R. Task-specific deep eutectic solvent based extraction coupled cascade chromatography quantification of α-glucosidase inhibitory peptide from Ocimum tenuriflorum seeds. Microchem. J. 2020, 157, 104883. [Google Scholar] [CrossRef]
- Mojica, L.; González, E.; Mejía, D. Optimization of enzymatic production of anti-diabetic peptides from black bean (Phaseolus vulgaris L.) proteins, their characterization and biological potential. Food Funct. 2016, 7, 713–727. [Google Scholar] [CrossRef]
- Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642–646. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: a new approach for rapid, accurate docking and scoring. 1. method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra precision glide: docking and scoring incorporating a model of hydrophobic enclosure for protein−ligand complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Kong, K.W.; Wu, D.T.; Liu, H.Y.; Li, H.B.; Zhang, J.R.; Gan, R.Y. Pomegranate peel-derived punicalagin: Ultrasonic-assisted extraction, purification, and its α-glucosidase inhibitory mechanism. Food Chem. 2021, 374, 131635. [Google Scholar] [CrossRef]
- Ren, Y.; Wu, H.; Lai, F.; Yang, M.Y.; Li, X.F.; Tang, Y.Q. Isolation and identification of a novel anticoagulant peptide from enzymatic hydrolysates of scorpion (Buthus martensii Karsch) protein. Food Res. Int. 2014, 64, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Admassu, H.; Gasmalla, M.; Yang, R.; Zhao, W. Identification of novel bioactive peptides with α-amylase inhibitory potential from enzymatic protein hydrolysates of red seaweed (Porphyra spp). J. Agric. Food Chem. 2018, 66, 4872–4882. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.D.; Zhang, F.M.; Ma, Z.R.; Chen, S.; Ding, G.T.; Tian, X.J.; Feng, R.F. Isolation and identification of the angiotensin-I converting enzyme (ACE) inhibitory peptides derived from cottonseed protein: Optimization of hydrolysis conditions. Int. J. Food Prop. 2019, 22, 1296–1309. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Zhang, G.Z.; Zhang, Y.H. Optimization of enzymatic hydrolysis of protein from small yellow croaker (Psendosciaena polyactis) using response surface methodology. Adv. J. Food Sci. Technol. 2016, 11, 1–6. [Google Scholar] [CrossRef]
- Wu, S.G.; Sun, J.H.; Tong, Z.F.; Lan, X.D.; Zhao, Z.X.; Liao, D.K. Optimization of hydrolysis conditions for the production of angiotensin-I converting enzyme-inhibitory peptides and isolation of a novel peptide from lizard fish (Saurida elongata) muscle protein hydrolysate. Mar. Drugs 2012, 10, 1066–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Liang, K.; Jin, Y.Q.; Zhang, M.M.; Chen, Y.; Wu, H.; Lai, F.R. Identification and characterization of two novel α-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Oseguera-Toledo, M.E.; Elvira, G.; Amaya-Llano, S.L. Hard-to-cook bean (Phaseolus vulgaris L.) proteins hydrolyzed by alcalase and bromelain produced bioactive peptide fractions that inhibit targets of type-2 diabetes and oxidative stress. J. Food Sci. 2015, 76, 839–851. [Google Scholar] [CrossRef]
- Feng, Y.X.; Ruan, G.R.; Jin, F.; Xu, J.; Wang, F.J. Purification, identification, and synthesis of five novel antioxidant peptides from chinese chestnut (Castanea mollissima Blume) protein hydrolysates. LWT 2018, 92, 40–46. [Google Scholar] [CrossRef]
- Church, D.D.; Hirsch, K.R.; Park, S.; Kim, I.Y.; Gwin, J.A.; Pasiakos, S.M.; Wolfe, R.R.; Ferrando, A.A. Essential amino acids and protein synthesis: Insights into maximizing the muscle and whole-body response to feeding. Nutrients 2020, 12, 3717. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.G.; Dias, S.S.; de Angelis, T.R.; Fernandes, E.V.; Bernardes, A.G.; Milanez, V.F.; Jussiani, E.I.; de Paula Ramos, S. The use of BCAA to decrease delayed-onset muscle soreness after a single bout of exercise: A systematic review and meta-analysis. Amino Acids 2021, 53, 1663–1678. [Google Scholar] [CrossRef] [PubMed]
- Mollica, A.; Zengin, G.; Durdagi, S.; Ekhteiari Salmas, R.; Macedonio, G.; Stefanucci, A.; Dimmito, M.P.; Novellino, E. Combinatorial peptide library screening for discovery of diverse α-glucosidase inhibitors using molecular dynamics simulations and binary QSAR models. J. Biomol. Struct. Dyn. 2019, 37, 726–740. [Google Scholar] [CrossRef] [Green Version]
- Auwal, I.M.; Bester, M.J.; Neitz, A.W.; Gaspar, A. Rational in silico design of novel α-glucosidase inhibitory peptides and in vitro evaluation of promising candidates. Biomed. Pharmacother. 2018, 107, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.C.; Zhao, H.X.; Pan, X.X.; Orfila, C.; Lu, W.H.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of alpha-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Cardullo, N.; Muccilli, V.; Pulvirenti, L.; Cornu, A.; Pouységu, L.; Deffieux, D.; Quideau, S.; Tringali, C. C-glucosidic ellagitannins and galloylated glucoses as potential functional food ingredients with anti-diabetic properties: A study of α-glucosidase and α-amylase inhibition. Food Chem. 2020, 313, 126099. [Google Scholar] [CrossRef]
- Wu, J.J.; Xu, Y.B.; Liu, X.Y.; Chen, M.M.; Zhu, B.; Wang, H.J.; Shi, S.S.; Qin, L.P.; Wang, S.C. Isolation and structural characterization of a non-competitive α-glucosidase inhibitory polysaccharide from the seeds of Litchi chinensis Sonn. Int. J. Biol. Macromol. 2020, 154, 1105–1115. [Google Scholar] [CrossRef]
- Morris, G.M.; Lim-Wilby, M. Molecular docking. Methods Mol. Biol. 2008, 443, 365–382. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Aluko, R.E. Kinetics and molecular docking studies of the inhibitions of angiotensin converting enzyme and renin activities by hemp seed (Cannabis sativa L.) peptides. J. Agric. Food Chem. 2014, 62, 4135–4144. [Google Scholar] [CrossRef]
- Han, L.; Fang, C.; Zhu, R.X.; Peng, Q.; Li, D.; Wang, M. Inhibitory effect of phloretin on α-glucosidase: Kinetics, interaction mechanism and molecular docking. Int. J. Biol. Macromol. 2017, 95, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Sim, L.; Quezada-Calvillo, R.; Sterchi, E.E.; Nichols, B.L.; Rose, D.R. Human intestinal maltase–glucoamylase: Crystal structure of the N-terminal catalytic subunit and basis of inhibition and substrate specificity. J. Mol. Biol. 2008, 375, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Campos, M.S.; Guerrero, L.C.; Betancur, D.A.; Hernandez-Escalante, V.M. Bioavailability of Bioactive Peptides. Food Rev. Int. 2011, 27, 213–226. [Google Scholar] [CrossRef]
- Qian, J.J.; Zheng, L.; Zhao, Y.J.; Zhao, M.M. Stability, bioavailability, and structure–activity relationship of casein-derived peptide YPVEPF with a sleep-enhancing effect. J. Agric. Food Chem. 2022, 70, 14947–14958. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Sum of Squares | df | Mean Square | F Value | p-Value | Significance |
|---|---|---|---|---|---|---|
| Model | 992.59 | 14 | 70.90 | 21.37 | <0.0001 | ** |
| A-Temperature | 0.6120 | 1 | 0.6120 | 0.1845 | 0.6741 | |
| B-pH | 38.81 | 1 | 38.81 | 11.70 | 0.0041 | ** |
| C-Time | 1.12 | 1 | 1.12 | 0.3383 | 0.5701 | |
| D-Protease concentration | 96.22 | 1 | 96.22 | 29.00 | <0.0001 | ** |
| AB | 0.7056 | 1 | 0.7056 | 0.2127 | 0.6518 | |
| AC | 0.6889 | 1 | 0.6889 | 0.2076 | 0.6556 | |
| AD | 0.3080 | 1 | 0.3080 | 0.0928 | 0.7651 | |
| BC | 0.0182 | 1 | 0.0182 | 0.0055 | 0.9420 | |
| BD | 0.1260 | 1 | 0.1260 | 0.0380 | 0.8483 | |
| CD | 0.5041 | 1 | 0.5041 | 0.1519 | 0.7025 | |
| A2 | 24.75 | 1 | 24.75 | 7.46 | 0.0162 | * |
| B2 | 249.52 | 1 | 249.52 | 75.21 | <0.0001 | ** |
| C2 | 45.93 | 1 | 45.93 | 13.84 | 0.0023 | ** |
| D2 | 731.54 | 1 | 731.54 | 220.50 | <0.0001 | ** |
| Residual | 46.45 | 14 | 3.32 | |||
| Lack of Fit | 43.18 | 10 | 4.32 | 5.29 | 0.0614 | Not significance |
| Pure Error | 3.27 | 4 | 0.8170 | |||
| Cor Total | 1039.04 | 28 |
| Molecular Weight (kDa) | α-glucosidase Inhibitory Activity (IC50, mg/mL) |
|---|---|
| CSCPH (Unfractionated) | 8.442 ± 0.33 b |
| CSCPH-I (MW <1 kDa) | 59.450 ± 0.893 d |
| CSCPH-II (1 kDa < MW < 3 kDa) | 3.896 ± 0.148 a |
| CSCPH-III (3 kDa < MW < 10 kDa) | 62.440 ± 0.965 d |
| CSCPH-IV (MW > 10 kDa) | 15.800 ± 0.760 c |
| Amino Acids | CSCP (%) | CSCPH (%) | CSCPH-II (%) | CSCPH-II-4 (%) |
|---|---|---|---|---|
| Asp | 7.64 ± 0.11 a | 8.22 ± 1.07 a | 7.92 ± 0.74 a | 6.79 ± 0.11 b |
| Glu | 19.78 ± 0.71 a | 21.18 ± 4.37 a | 16.47 ± 0.61 b | 10.42 ± 0.08 c |
| Ser | 3.98 ± 0.02 bc | 3.86 ± 0.48 c | 4.37 ± 0.32 a | 4.16 ± 0.13 ab |
| Gly | 3.94 ± 0.03 b | 3.93 ± 0.34 b | 4.84 ± 0.17 a | 4.91 ± 0.06 a |
| His | 1.48 ± 0.01 b | 1.54 ± 0.21 b | 1.72 ± 0.33 b | 4.03 ± 0.05 a |
| Arg | 2.42 ± 0.01 c | 2.47 ± 0.28 c | 3.81 ± 0.33 b | 4.41 ± 0.10 a |
| Thr | 4.92 ± 0.04 b | 4.79 ± 0.57 b | 6.19 ± 0.79 a | 5.70 ± 0.07 a |
| Ala | 15.81 ± 0.07 a | 16.48 ± 1.13 a | 13.79 ± 1.60 a | 7.90 ± 0.09 b |
| Pro | 10.89 ± 0.09 bc | 10.24 ± 1.31 c | 11.59 ± 1.02 b | 13.19 ± 0.13 a |
| Tyr | 2.55 ± 0.08 b | 2.53 ± 0.30 b | 2.58 ± 0.15 b | 6.67 ± 0.07 a |
| Val | 2.86 ± 0.02 b | 2.64 ± 0.32 b | 3.77 ± 0.41 a | 3.81 ± 0.04 a |
| Met | 2.53 ± 0.04 a | 2.40 ± 0.33 a | 2.50 ± 0.19 a | 0.81 ± 0.03 b |
| Leu | 1.46 ± 0.10 b | 1.22 ± 0.03 b | 1.47 ± 0.17 b | 4.66 ± 0.06 a |
| Ile | 3.23 ± 0.02 c | 3.20 ± 0.34 c | 3.90 ± 0.21 b | 4.21 ± 0.05 a |
| Phe | 7.01 ± 0.03 ab | 6.70 ± 0.78 b | 7.08 ± 0.57 ab | 7.43 ± 0.14 a |
| Trp | 3.49 ± 0.02 c | 3.31 ± 0.36 d | 3.97 ± 0.15 b | 7.41 ± 0.16 a |
| Lys | 6.00 ± 0.05 a | 5.28 ± 0.64 b | 4.02 ± 0.38 c | 3.48 ± 0.09 d |
| EAA | 31.51 ± 0.13 b | 29.58 ± 1.11 c | 32.87 ± 0.95 b | 37.50 ± 0.41 a |
| HAA | 47.30 ± 0.28 ab | 46.36 ± 1.50 b | 48.15 ± 1.61 ab | 49.47 ± 0.41 a |
| BCAA | 7.55 ± 0.06 c | 7.09 ± 0.27 d | 9.14 ± 0.08 b | 12.63 ± 0.33 a |
| α-glucosidase inhibitory (IC50, mg/mL) activity (IC50, mg/mL) | nd | 8.44 ± 0.33 a | 3.90 ± 0.15 b | 2.03 ± 0.09 c |
| Peptides Sequence | de Novo Score | Mass | m/z | XP Score (kcal/mol) | α-glucosidase Inhibitory Activity (IC50, mM) |
|---|---|---|---|---|---|
| LLVLYYEY | 94 | 1074.56 | 538.287 | −9.335 | 0.33 |
| LLLLPSYSEF | 92 | 1180.64 | 591.3289 | −9.060 | 1.11 |
| LCDQCPPHA | 91 | 1096.44 | 549.229 | −8.535 | 4.32 |
| ATNPPCCQP | 90 | 1043.42 | 522.7232 | −8.868 | >10 |
| KDDFVEKR | 99 | 1035.53 | 518.7742 | −8.307 | >10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wu, F.; He, Z.; Fang, X.; Liu, X. Optimization and Molecular Mechanism of Novel α-Glucosidase Inhibitory Peptides Derived from Camellia Seed Cake through Enzymatic Hydrolysis. Foods 2023, 12, 393. https://doi.org/10.3390/foods12020393
Zhang Y, Wu F, He Z, Fang X, Liu X. Optimization and Molecular Mechanism of Novel α-Glucosidase Inhibitory Peptides Derived from Camellia Seed Cake through Enzymatic Hydrolysis. Foods. 2023; 12(2):393. https://doi.org/10.3390/foods12020393
Chicago/Turabian StyleZhang, Yuanping, Fenghua Wu, Zhiping He, Xuezhi Fang, and Xingquan Liu. 2023. "Optimization and Molecular Mechanism of Novel α-Glucosidase Inhibitory Peptides Derived from Camellia Seed Cake through Enzymatic Hydrolysis" Foods 12, no. 2: 393. https://doi.org/10.3390/foods12020393
APA StyleZhang, Y., Wu, F., He, Z., Fang, X., & Liu, X. (2023). Optimization and Molecular Mechanism of Novel α-Glucosidase Inhibitory Peptides Derived from Camellia Seed Cake through Enzymatic Hydrolysis. Foods, 12(2), 393. https://doi.org/10.3390/foods12020393