


Triflumizole Induces Developmental Toxicity, Liver Damage, Oxidative Stress, Heat Shock Response, Inflammation, and Lipid Synthesis in Zebrafish




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Zebrafish Maintenance and Breeding
2.3. Zebrafish Exposure
2.4. Fish Intestinal Cell Line Exposure
2.5. Morphological Assay
2.6. Enzyme Activities Assay
2.7. Quantitative Real Time PCR (qPCR)
2.8. Statistical Analysis
3. Results
3.1. Exposure Concentrations
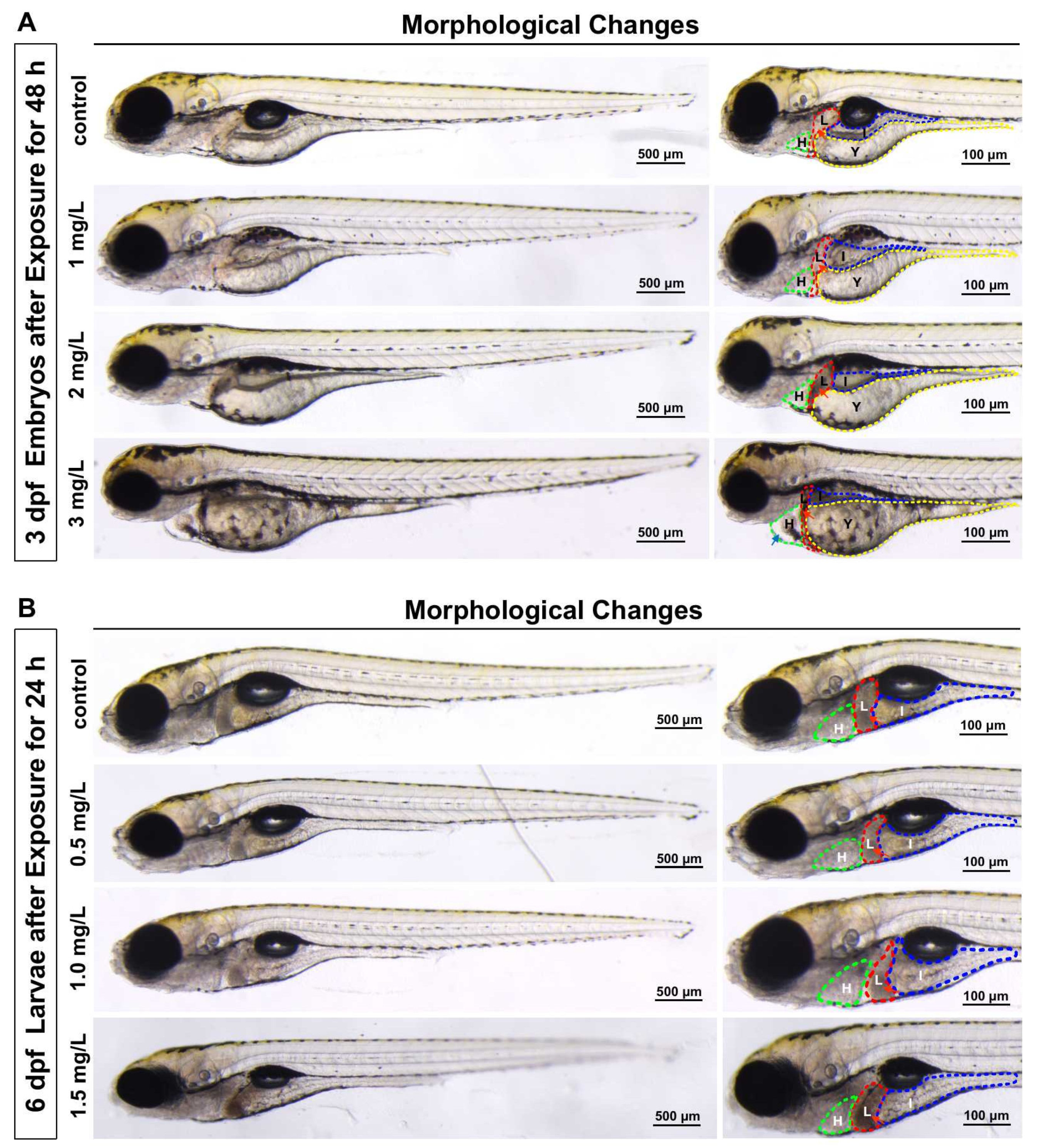
3.2. Zebrafish Larvae Morphology after TFZ Exposure
3.3. Enzyme Activities after TFZ Exposure
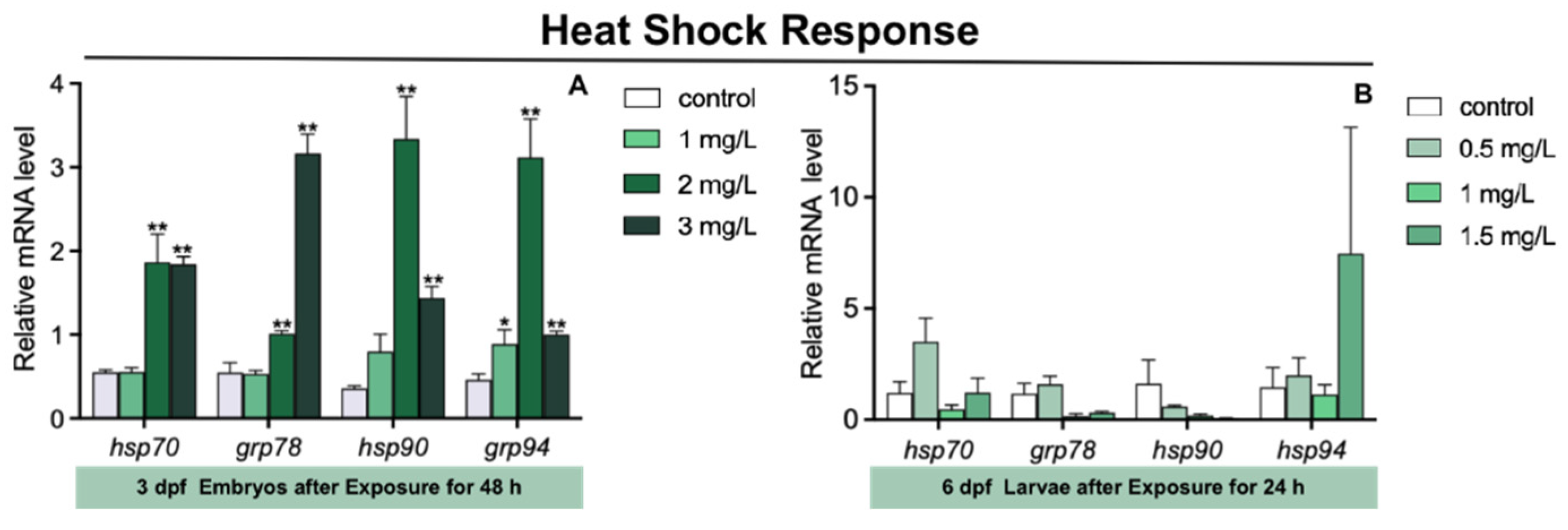
3.4. Heat Shock Response in Zebrafish Larvae after TFZ Exposure
3.5. Inflammatory Genes Expression in Zebrafish Larvae after TFZ Exposure
3.6. Lipid Synthesis Gene Expression in Zebrafish Larvae after TFZ Exposure
3.7. Effects of TFZ on Fish Intestinal Cell Line
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oerke, E. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Popp, J.; Pető, K.; Nagy, J. Pesticide productivity and food security. A review. Agron. Sustain 2013, 33, 243–255. [Google Scholar] [CrossRef]
- Konwick, B.J.; Garrison, A.W.; Avants, J.K.; Fisk, A.T. Bioaccumulation and biotransformation of chiral triazole fungicides in rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2006, 80, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Nakata, A.; Hashimoto, S.; Ikura, K.; Katsuura, K. Development of a New Fungicide Triflumizole. J. Pestic. Sci. 1991, 16, 301–313. [Google Scholar] [CrossRef] [Green Version]
- İnam, R.; Gülerman, E.; Sarıgül, T. Determination of triflumizole by differential pulse polarography in formulation, soil and natural water samples. Anal. Chim. Acta 2006, 579, 117–123. [Google Scholar] [CrossRef]
- Fajardo, J. Triflumizole (Procure): Control of diseases in tree nut, fruit, vegetable, and vine crops. Phytopathology 2006, 96. [Google Scholar]
- Tang, G.; Zhang, W.B.; Tang, J.Y.; Niu, J.F.; Yang, J.L.; Dong, H.Q.; Liang, Y.; Luo, L.X.; Cao, Y.S. Development of triflumizole ionic liquids containing anions of natural origin for improving the utilization and minimizing the adverse impacts on aquatic ecosystems. Sci. Total Environ. 2019, 670, 606–612. [Google Scholar] [CrossRef]
- Brancato, A.; Brocca, D.; De Lentdecker, C.; Erdos, Z.; Ferreira, L.; Greco, L.; Jarrah, S.; Kardassi, D.; Leuschner, R.; Lythgo, C. European Food Safety. Efsa J. 2017, 15, 47–49. [Google Scholar]
- Li, X.; Pham, H.T.; Janesick, A.S.; Blumberg, B. Triflumizole is an obesogen in mice that acts through peroxisome proliferator activated receptor gamma (PPAR γ). Environ. Health Perspect. 2012, 120, 1720–1726. [Google Scholar] [CrossRef] [Green Version]
- Khay, S.; El-Aty, A.M.A.; Choi, J.H.; Shim, J.H. Analysis of residual triflumizole, an imidazole fungicide, in apples, pears and cucumbers using high performance liquid chromatography. Toxicol. Res. 2008, 24, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Maurya, P.K.; Malik, D.S.; Sharma, A. Impacts of pesticide application on aquatic environments and fish diversity. Contam. Agric. Environ. Health Risks Remediat. 2019, 1, 111. [Google Scholar]
- Xi, J.; Shao, J.; Wang, Y.; Wang, X.; Yang, H.; Zhang, X.; Xiong, D. Acute toxicity of triflumizole to freshwater green algae Chlorella vulgaris. Pestic. Biochem. Physiol. 2019, 158, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Liu, L.; Gong, Y.X.; Ling, F.; Wang, G.X. Triazole-induced toxicity in developing rare minnow (Gobiocypris rarus) embryos. Environ. Sci. Pollut. Res. 2014, 21, 13625–13635. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.L.; Trivella, D.B.; Trevisan, R.; Dinslaken, D.F.; Marques, M.R.; Bainy, A.C.; Dafre, A.L. Antioxidant status and stress proteins in the gills of the brown mussel Perna perna exposed to zinc. Chem.-Biol. Interact. 2006, 160, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, I.; Parvez, S.; Pandey, S.; Bin-Hafeez, B.; Haque, R.; Raisuddin, S. Oxidative stress biomarkers of exposure to deltamethrin in freshwater fish, Channa punctatus Bloch. Ecotoxicol. Environ. Saf. 2003, 56, 295–301. [Google Scholar] [CrossRef]
- Li, Z.H.; Zlabek, V.; Velisek, J.; Grabic, R.; Machova, J.; Randak, T. Modulation of antioxidant defence system in brain of rainbow trout (Oncorhynchus mykiss) after chronic carbamazepine treatment. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2010, 151, 137–141. [Google Scholar] [CrossRef]
- Pereira, T.C.B.; Campos, M.M.; Bogo, M.R. Copper toxicology, oxidative stress and inflammation using zebrafish as experimental model. J. Appl. Toxicol. 2016, 36, 86–885. [Google Scholar] [CrossRef]
- Kurashova, N.A.; Madaeva, I.M.; Kolesnikova, L.I. Expression of heat shock proteins HSP70 under oxidative stress. Adv. Gerontol.=Uspekhi Gerontol. 2019, 32, 502–508. [Google Scholar] [CrossRef]
- Madaeva, I.M.; Kurashova, N.A.; Semenova, N.V.; Ukhinov, E.B.; Kolesnikov, S.I.; Kolesnikova, L.I. Heat Shock Protein HSP70 in Oxidative Stress in Apnea Patients. Bull. Exp. Biol. Med. 2020, 169, 695–697. [Google Scholar] [CrossRef]
- Szyller, J.; Bil-Lula, I. Heat shock proteins in oxidative stress and ischemia/reperfusion injury and benefits from physical exercises: A review to the current knowledge. Oxidative Med. Cell. Longev. 2021, 2021, 6678457. [Google Scholar] [CrossRef]
- Wang, Y.; Teng, M.; Wang, D.; Yan, J.; Miao, J.; Zhou, Z.; Zhu, W. Enantioselective bioaccumulation following exposure of adult zebrafish (Danio rerio) to epoxiconazole and its effects on metabolomic profile as well as genes expression. Environ. Pollut. 2017, 229, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, W.; Wang, D.; Teng, M.; Yan, J.; Miao, J.; Zhou, Z. 1H NMR-based metabolomics analysis of adult zebrafish (Danio rerio) after exposure to diniconazole as well as its bioaccumulation behavior. Chemosphere 2017, 168, 1571–1577. [Google Scholar] [CrossRef] [PubMed]
- Jia, M.; Wang, Y.; Wang, D.; Teng, M.; Yan, J.; Yan, S.; Meng, Z.; Li, R.; Zhou, Z.; Zhu, W. The effects of hexaconazole and epoxiconazole enantiomers on metabolic profile following exposure to zebrafish (Danio rerio) as well as the histopathological changes. Chemosphere 2019, 226, 520–533. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Jia, K.; Huang, L.; Liao, X.; Guo, X.; Lu, H. Hepatotoxicity of tricyclazole in zebrafish (Danio rerio). Chemosphere 2019, 232, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.; Chen, G.; Zeng, J.; Liu, F.; Liao, X.; Guo, C.; Luo, J.; Xiong, G.; Lu, H. Low trifloxystrobin-tebuconazole concentrations induce cardiac and developmental toxicity in zebrafish by regulating notch mediated-oxidative stress generation. Ecotoxicol. Environ. Saf. 2022, 241, 113752. [Google Scholar] [CrossRef]
- Shi, P.; Liao, K.; Xu, J.; Wang, Y.; Xu, S.; Yan, X. Eicosapentaenoic acid mitigates palmitic acid-induced heat shock response, inflammation and repair processes in fish intestine. Fish Shellfish Immunol. 2022, 124, 362–371. [Google Scholar] [CrossRef]
- Batista-Silva, H.; Dambrós, B.F.; de Moura, K.R.S.; Elie, N.; Delalande, C.; Van Der Kraak, G.; Silva, F.R.M.B. Calcium influx and spermatogenesis in the testis and liver enzyme activities in the zebrafish are rapidly modulated by the calcium content of the water. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2022, 270, 111227. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.J.; Green, A. An international database for pesticide risk assessments and management. Hum. Ecol. Risk Assess. Int. J. 2016, 22, 1050–1064. [Google Scholar] [CrossRef] [Green Version]
- Hermsen, S.A.; van den Brandhof, E.J.; van der Ven, L.T.; Piersma, A.H. Relative embryotoxicity of two classes of chemicals in a modified zebrafish embryotoxicity test and comparison with their in vivo potencies. Toxicol. Vitr. 2011, 25, 745–753. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Teng, M.; Meng, Z.; Yan, S.; Jia, M.; Li, R.; Liu, L.; Yan, J.; Zhou, Z.; Zhu, W. Toxicity effects in zebrafish embryos (Danio rerio) induced by prothioconazole. Environ. Pollut. 2019, 255, 113269. [Google Scholar] [CrossRef]
- Qin, Z.; Wang, W.; Weng, Y.; Bao, Z.; Yang, G.; Jin, Y. Bromuconazole exposure induces cardiotoxicity and lipid transport disorder in larval zebrafish. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 262, 109451. [Google Scholar] [CrossRef]
- Mu, X.; Pang, S.; Sun, X.; Gao, J.; Chen, J.; Chen, X.; Li, X.; Wang, C. Evaluation of acute and developmental effects of difenoconazole via multiple stage zebrafish assays. Environ. Pollut. 2013, 175, 147–157. [Google Scholar] [CrossRef]
- Jiang, J.; Chen, L.; Wu, S.; Lv, L.; Liu, X.; Wang, Q.; Zhao, X. Effects of difenoconazole on hepatotoxicity, lipid metabolism and gut microbiota in zebrafish (Danio rerio). Environ. Pollut. 2020, 265, 114844. [Google Scholar] [CrossRef]
- Yu, Q.; Huo, J.; Zhang, Y.; Liu, K.; Cai, Y.; Xiang, T.; Jiang, Z.; Zhang, L. Tamoxifen-induced hepatotoxicity via lipid accumulation and inflammation in zebrafish. Chemosphere 2020, 239, 124705. [Google Scholar] [CrossRef] [PubMed]
- Jia, K.; Cheng, B.; Huang, L.; Xiao, J.; Bai, Z.; Liao, X.; Cao, Z.; Shen, T.; Zhang, C.; Hu, C. Thiophanate-methyl induces severe hepatotoxicity in zebrafish. Chemosphere 2020, 248, 125941. [Google Scholar] [CrossRef] [PubMed]
- Zoupa, M.; Machera, K. Zebrafish as an alternative vertebrate model for investigating developmental toxicity—The triadimefon example. Int. J. Mol. Sci. 2017, 18, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.; Parvez, S.; Sayeed, I.; Haque, R.; Bin-Hafeez, B.; Raisuddin, S. Biomarkers of oxidative stress: A comparative study of river Yamuna fish Wallago attu (Bl. & Schn.). Sci. Total Environ. 2003, 309, 105–115. [Google Scholar]
- John, S.; Kale, M.; Rathore, N.; Bhatnagar, D. Protective effect of vitamin E in dimethoate and malathion induced oxidative stress in rat erythrocytes. J. Nutr. Biochem. 2001, 12, 500–504. [Google Scholar] [CrossRef]
- Liu, Z.; Shangguan, Y.; Zhu, P.; Sultan, Y.; Feng, Y.; Li, X.; Ma, J. Developmental toxicity of glyphosate on embryo-larval zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2022, 236, 113493. [Google Scholar] [CrossRef]
- Lu, K.; Qiao, R.; An, H.; Zhang, Y. Influence of microplastics on the accumulation and chronic toxic effects of cadmium in zebrafish (Danio rerio). Chemosphere 2018, 202, 514–520. [Google Scholar] [CrossRef]
- Mu, X.; Chai, T.; Wang, K.; Zhang, J.; Zhu, L.; Li, X.; Wang, C. Occurrence and origin of sensitivity toward difenoconazole in zebrafish (Danio reio) during different life stages. Aquat. Toxicol. 2015, 160, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.N.; Zhang, J.L.; Ren, H.T.; Zhou, B.H.; Wu, Q.J.; Sun, P. Effect of tributyltin on antioxidant ability and immune responses of zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2017, 138, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Zhang, Z.; Ran, C.; He, S.; Yang, Y.; Du, Z.; Zhang, J.; Zhou, Z. The hepatotoxicity of palmitic acid in zebrafish involves the intestinal microbiota. J. Nutr. 2018, 148, 1217–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pressley, M.E.; Phelan III, P.E.; Witten, P.E.; Mellon, M.T.; Kim, C.H. Pathogenesis and inflammatory response to Edwardsiella tarda infection in the zebrafish. Dev. Comp. Immunol. 2005, 29, 501–513. [Google Scholar] [CrossRef]
- Ko, E.Y.; Cho, S.H.; Kwon, S.H.; Eom, C.Y.; Jeong, M.S.; Lee, W.; Kim, S.Y.; Heo, S.J.; Ahn, G.; Lee, K.P.; et al. The roles of NF-κB and ROS in regulation of pro-inflammatory mediators of inflammation induction in LPS-stimulated zebrafish embryos. Fish Shellfish Immunol. 2017, 68, 525–529. [Google Scholar] [CrossRef]
- Krishnaraj, C.; Harper, S.L.; Yun, S.I. In Vivo toxicological assessment of biologically synthesized silver nanoparticles in adult Zebrafish (Danio rerio). J. Hazard. Mater. 2016, 301, 480–491. [Google Scholar] [CrossRef] [Green Version]
- Jönsson, M.E.; Kubota, A.; Timme-Laragy, A.R.; Woodin, B.; Stegeman, J.J. Ahr2-dependence of PCB126 effects on the swim bladder in relation to expression of CYP1 and cox-2 genes in developing zebrafish. Toxicol. Appl. Pharmacol. 2012, 265, 166–174. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.X.; Zhuo, M.Q.; Li, D.D.; Xu, Y.H.; Wu, K.; Luo, Z. SREBP-1 and LXRα pathways mediated Cu-induced hepatic lipid metabolism in zebrafish Danio rerio. Chemosphere 2019, 215, 370–379. [Google Scholar] [CrossRef]
- Zhang, J.; Qian, L.; Teng, M.; Mu, X.; Qi, S.; Chen, X.; Zhou, Y.; Cheng, Y.; Pang, S.; Li, X.; et al. The lipid metabolism alteration of three spirocyclic tetramic acids on zebrafish (Danio rerio) embryos. Environ. Pollut. 2019, 248, 715–725. [Google Scholar] [CrossRef]
- Jin, M.; Zhang, B.; Sun, Y.; Zhang, S.; Li, X.; Sik, A.; Bai, Y.; Zheng, X.; Liu, K. Involvement of peroxisome proliferator-activated receptor γ in anticonvulsant activity of α-asaronol against pentylenetetrazole-induced seizures in zebrafish. Neuropharmacology 2020, 162, 107760. [Google Scholar] [CrossRef]
- Shi, P.; Meng, R.; Liao, K.; Li, S.; Hu, J.; Xu, J.; Zhang, L.; Cao, J.; Ran, Z.; Wang, D.; et al. Cadmium transcriptionally regulates Scd1 expression in silver pomfret. Environ. Toxicol. 2020, 35, 404–413. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, L.; Shi, P.; Jia, K.; Yin, H.; Xu, J.; Yan, X.; Liao, K. Triflumizole Induces Developmental Toxicity, Liver Damage, Oxidative Stress, Heat Shock Response, Inflammation, and Lipid Synthesis in Zebrafish. Toxics 2022, 10, 698. https://doi.org/10.3390/toxics10110698
Bai L, Shi P, Jia K, Yin H, Xu J, Yan X, Liao K. Triflumizole Induces Developmental Toxicity, Liver Damage, Oxidative Stress, Heat Shock Response, Inflammation, and Lipid Synthesis in Zebrafish. Toxics. 2022; 10(11):698. https://doi.org/10.3390/toxics10110698
Chicago/Turabian StyleBai, Lina, Peng Shi, Kun Jia, Hua Yin, Jilin Xu, Xiaojun Yan, and Kai Liao. 2022. "Triflumizole Induces Developmental Toxicity, Liver Damage, Oxidative Stress, Heat Shock Response, Inflammation, and Lipid Synthesis in Zebrafish" Toxics 10, no. 11: 698. https://doi.org/10.3390/toxics10110698
APA StyleBai, L., Shi, P., Jia, K., Yin, H., Xu, J., Yan, X., & Liao, K. (2022). Triflumizole Induces Developmental Toxicity, Liver Damage, Oxidative Stress, Heat Shock Response, Inflammation, and Lipid Synthesis in Zebrafish. Toxics, 10(11), 698. https://doi.org/10.3390/toxics10110698