
Nanoplastic Exposure at Predicted Environmental Concentrations Induces Activation of Germline Ephrin Signal Associated with Toxicity Formation in the Caenorhabditis elegans Offspring



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. PS-NP Properties
2.2. C. elegans Maintenance
2.3. Exposure
2.4. Endpoints
2.5. Transcriptional Expression Analysis
2.6. RNA Interference (RNAi)
2.7. HiSeq 2000 Sequencing
2.8. Construct Generation and Transgene
2.9. Data Analysis
3. Results
3.1. Dysregulated Genes Required for Stress Response Control Were Not Candidate Targets of KSR-1/2 in Controlling Transgenerational PS-NP Toxicity
3.2. Identification of Ephrin Ligand EFN-3 as Candidate Downstream Target of KSR-1/2 in Controlling Transgenerational PS-NP Toxicity
3.3. Genetic Interaction between EFN-3 and KSR-1/2 in Controlling Transgenerational PS-NP Toxicity
3.4. Ephrin Receptor VAB-1 Was Involved in Controlling Transgenerational PS-NP Toxicity
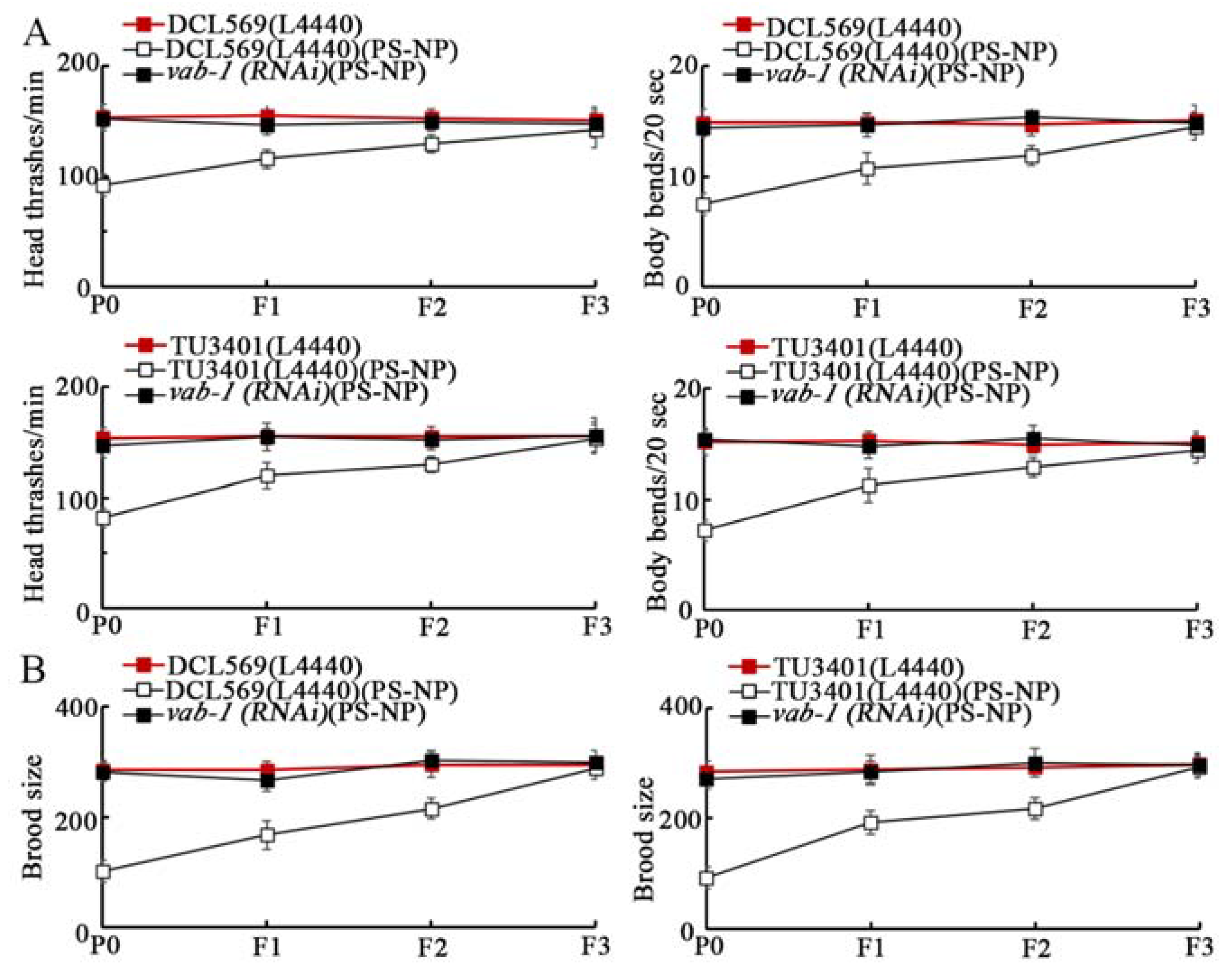
3.5. Transgenerational Association between Germline Ephrin Ligand EFN-3 and Its Receptor in Controlling PS-NP Toxicity
3.6. Tissue-Specific Activity of VAB-1 in Controlling Transgenerational PS-NPs Toxicity
3.7. Identification of Potential Targets of Neuronal VAB-1 in Controlling PS-NP Toxicity
3.8. Identification of Potential Targets of Germline VAB-1 in Controlling PS-NP Toxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Geyer, R.; Jambeck, J.; Law, K. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, F.; Watanabe, M.; Shibata, M.; Yokoyama, S.; Sudate, Y.; Hayashi, S. Comparative study on biodegradability of polyethylene wax by bacteria and fungi. Polym. Degrad. Stab. 2004, 86, 105–114. [Google Scholar] [CrossRef]
- Jahnke, A.; Arp, H.P.H.; Escher, B.I.; Gewert, B.; Gorokhova, E.; Kühnel, D.; MacLeod, M. Reducing uncertainty and confronting ignorance about the possible impacts of weathering plastic in the marine environment. Environ. Sci. Technol. Lett. 2017, 4, 85–90. [Google Scholar] [CrossRef] [Green Version]
- NJDEP-Science Advisory Board. Human Health Impacts of Microplastics and Nanoplastics. 2015. Available online: https://dep.nj.gov/wp-content/uploads/sab/sab-microplastics-nanoplastics.pdf (accessed on 1 October 2022).
- Sharma, S.; Chatterjee, S. Microplastic pollution, a threat to marine ecosystem and human health: A short review. Environ. Sci. Pollut. Res. Int. 2017, 24, 21530–21547. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.C.; Yu, H.; Fu, P.P. Toxicity and environmental risks of nanomaterials: Challenges and future needs. J. Environ. Sci. Health Part C Environ. Carcino. Ecotoxicol. Rev. 2009, 27, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Torres-Ruiz, M.; De la Vieja, A.; de Alba Gonzalez, M.; Esteban Lopez, M.; Castaño Calvo, A.; Cañas Portilla, A.I. Toxicity of nanoplastics for zebrafish embryos, what we know and where to go next. Sci. Total Environ. 2021, 797, 149125. [Google Scholar] [CrossRef]
- Bhagat, J.; Nishimura, N.; Shimada, Y. Toxicological interactions of microplastics/nanoplastics and environmental contaminants: Current knowledge and future perspectives. J. Hazard. Mater. 2021, 405, 123913. [Google Scholar] [CrossRef]
- Singh, S.; Kumar Naik, T.S.S.; Anil, A.G.; Dhiman, J.; Kumar, V.; Dhanjal, D.S.; Aguilar-Marcelino, L.; Singh, J.; Ramamurthy, P.C. Micro (nano) plastics in wastewater: A critical review on toxicity risk assessment, behaviour, environmental impact and challenges. Chemosphere 2022, 290, 133169. [Google Scholar] [CrossRef]
- Qu, M.; Nida, A.; Kong, Y.; Du, H.-H.; Xiao, G.-S.; Wang, D.-Y. Nanopolystyrene at predicted environmental concentration enhances microcystin-LR toxicity by inducing intestinal damage in Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2019, 183, 109568. [Google Scholar] [CrossRef]
- Ma, C.; Chen, Q.; Li, J.; Li, B.; Liang, W.; Su, L.; Shi, H. Distribution and translocation of micro- and nanoplastics in fish. Crit. Rev. Toxicol. 2021, 51, 740–753. [Google Scholar] [CrossRef]
- Leung, M.C.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef]
- Ruszkiewicz, J.A.; Pinkas, A.; Miah, M.R.; Weitz, R.L.; Lawes, M.J.A.; Akinyemi, A.J.; Ijomone, O.M.; Aschner, M. C. elegans as a model in developmental neurotoxicology. Toxicol. Appl. Pharmacol. 2018, 354, 126–135. [Google Scholar] [CrossRef]
- Wang, D.-Y. Exposure Toxicology in Caenorhabditis elegans; Springer Nature Singapore Pte Ltd.: Singapore, 2020. [Google Scholar]
- Kim, H.M.; Lee, D.K.; Long, N.P.; Kwon, S.W.; Park, J.H. Uptake of nanopolystyrene particles induces distinct metabolic profiles and toxic effects in Caenorhabditis elegans. Environ. Pollut. 2019, 246, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Wang, S.; Jin, Y.; Xue, W.; Zhong, Y.; Wang, H.; An, J.; Li, H. Polystyrene nanoparticles induced neurodevelopmental toxicity in Caenorhabditis elegans through regulation of dpy-5 and rol-6. Ecotoxicol. Environ. Saf. 2021, 222, 112523. [Google Scholar] [CrossRef] [PubMed]
- Schultz, C.L.; Bart, S.; Lahive, E.; Spurgeon, D.J. What is on the outside matters-surface charge and dissolve organic matter association affect the toxicity and physiological mode of action of polystyrene nanoplastics to C. elegans. Environ. Sci. Technol. 2021, 55, 6065–6075. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.-Y. Molecular Toxicology in Caenorhabditis elegans; Springer Nature Singapore Pte Ltd.: Singapore, 2019. [Google Scholar]
- Zhao, Y.-L.; Chen, J.-Y.; Wang, R.; Pu, X.-X.; Wang, D.-Y. A review of transgenerational and multigenerational toxicology in the in vivo model animal Caenorhabditis elegans. J. Appl. Toxicol. 2022. [Google Scholar] [CrossRef]
- Lenz, R.; Enders, K.; Nielsen, T.G. Microplastic exposure studies should be environmentally realistic. Proc. Natl. Acad. Sci. USA 2016, 113, E4121–E4122. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Qu, M.; Wong, G.; Wang, D.-Y. Transgenerational toxicity of nanopolystyrene particles in the range of μg/L in nematode Caenorhabditis elegans. Environ. Sci. Nano 2017, 4, 2356–2366. [Google Scholar] [CrossRef]
- Liu, H.-L.; Zhao, Y.-L.; Hua, X.; Wang, D.-Y. Induction of transgenerational toxicity is associated with the activated germline insulin signals in nematodes exposed to nanoplastic at predicted environmental concentrations. Ecotoxicol. Environ. Saf. 2022, 243, 114022. [Google Scholar] [CrossRef]
- Liu, H.-L.; Tian, L.-J.; Wang, S.-T.; Wang, D.-Y. Size-dependent transgenerational toxicity induced by nanoplastics in nematode Caenorhabditis elegans. Sci. Total Environ. 2021, 790, 148217. [Google Scholar] [CrossRef]
- Sun, L.-M.; Liao, K.; Wang, D.-Y. Comparison of transgenerational reproductive toxicity induced by pristine and amino modified nanoplastics in Caenorhabditis elegans. Sci. Total Environ. 2021, 768, 144362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, S.-T.; Zhao, Y.; Bi, K.; Wang, D.-Y. Increase in germline methyltransferases governing methylation of histone H3K9 is associated with transgenerational nanoplastic toxicity in Caenorhabditis elegans. Environ. Sci. Nano 2021, 9, 265–274. [Google Scholar] [CrossRef]
- Yu, C.W.; Luk, T.C.; Liao, V.H. Long-term nanoplastics exposure results in multi and trans-generational reproduction decline associated with germline toxicity and epigenetic regulation in Caenorhabditis elegans. J. Hazard. Mater. 2021, 412, 125173. [Google Scholar] [CrossRef]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2011, 466, 835–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, X.; Zhao, Y.; Yuan, Y.-J.; Zhang, L.; Bian, Q.; Wang, D.-Y. Nanoplastics cause transgenerational toxicity through inhibiting germline microRNA mir-38 in C. elegans. J. Hazard. Mater. 2022, 437, 129302. [Google Scholar] [CrossRef]
- Wong, D.J.; Lu, D.Y.; Protack, C.D.; Kuwahara, G.; Bai, H.; Sadaghianloo, N.; Tellides, G.; Dardik, A. Ephrin type-B receptor 4 activation reduces neointimal hyperplasia in human saphenous vein in vitro. J. Vasc. Surg. 2016, 63, 795–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, S.; Pei, D.; Shi, L.; Cui, Y.; Hong, Z. Ephrin-B2 inhibits Aβ25–35-induced apoptosis by alleviating endoplasmic reticulum stress and promoting autophagy in HT22 cells. Neurosci. Lett. 2019, 704, 50–56. [Google Scholar] [CrossRef]
- Theofanous, S.A.; Florens, M.V.; Appeltans, I.; Denadai Souza, A.; Wood, J.N.; Wouters, M.M.; Boeckxstaens, G.E. Ephrin-B2 signaling in the spinal cord as a player in post-inflammatory and stress-induced visceral hypersensitivity. Neurogastroenterol. Motil. 2020, 32, e13782. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, S.-T.; Zhao, Y.-L.; Nurdebek, B.; Bu, Y.-Q.; Wang, D.-Y. Long-term exposure to polystyrene nanoparticles causes transgenerational toxicity by affecting the function and expression of MEV-1 and DAF-2 signals in Caenorhabditis elegans. NanoImpact 2022, 26, 100403. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Wu, Q.-L.; Wang, D.-Y. Neuronal Gα subunits required for the control of response to polystyrene nanoparticles in the range of μg/L in C. elegans. Ecotoxicol. Environ. Saf. 2021, 225, 112732. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-Y.; Xu, R.-R.; Hua, X.; Rui, Q.; Wang, D.-Y. Multi-walled carbon nanotubes induce transgenerational toxicity associated with activation of germline long non-coding RNA linc-7 in C. elegans. Chemosphere 2022, 301, 134687. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-T.; Liu, H.-L.; Qu, M.; Wang, D.-Y. Response of tyramine and glutamate related signals to nanoplastic exposure in Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2021, 217, 112239. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.-R.; Hua, X.; Rui, Q.; Wang, D.-Y. Polystyrene nanoparticles caused dynamic alteration in mitochondrial unfolded protein response from parents to the offspring in C. elegans. Chemosphere 2022, 308, 136154. [Google Scholar] [CrossRef]
- Kurz, C.L.; Ewbank, J.J. Caenorabditis elegans: An emerging genetic model for the study of innate immunity. Nat. Rev. Genet. 2003, 4, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.-X.; Luo, L.-B.; Yang, Y.-H.; Kong, Y.; Li, Y.-H.; Wang, D.-Y. Potential toxicity of nanopolystyrene on lifespan and aging process of nematode Caenorhabditis elegans. Sci. Total Environ. 2020, 705, 135918. [Google Scholar] [CrossRef]
- Shao, H.-M.; Han, Z.-Y.; Krasteva, N.; Wang, D.-Y. Identification of signaling cascade in the insulin signaling pathway in response to nanopolystyrene particles. Nanotoxicology 2019, 13, 174–188. [Google Scholar] [CrossRef]
- Wang, X.; Roy, P.J.; Holland, S.J.; Zhang, L.W.; Culotti, J.G.; Pawson, T. Multiple ephrins control cell organization in C. elegans using kinase-dependent and –independent functions of the VAB-1 Eph receptor. Mol. Cell 1999, 4, 903–913. [Google Scholar] [CrossRef]
- Brisbin, S.; Liu, J.; Boudreau, J.; Peng, J.; Evangelista, M.; Chin-Sang, I. A role for C. elegans Eph RTK signaling in PTEN regulation. Dev. Cell 2009, 17, 459–469. [Google Scholar] [CrossRef] [Green Version]
- Grossman, E.N.; Giurumescu, C.A.; Chisholm, A.D. Mechanisms of ephrin receptor protein kinase-independent signaling in amphid axon guidance in Caenorhabditis elegans. Genetics 2013, 195, 899–913. [Google Scholar] [CrossRef]
- Qu, M.; Li, D.; Zhao, Y.-L.; Yuan, Y.-J.; Wang, D.-Y. Exposure to low-dose nanopolystyrene induces the response of neuronal JNK MAPK signaling pathway in nematode Caenorhabditis elegans. Environ. Sci. Euro. 2020, 32, 58. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-L.; Zhang, R.-J.; Wang, D.-Y. Response of DBL-1/TGF-β signaling-mediated neuron-intestine communication to nanopolystyrene in nematode Caenorhabditis elegans. Sci. Total Environ. 2020, 745, 1141047. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-T.; Liu, H.-L.; Zhao, Y.-Y.; Rui, Q.; Wang, D.-Y. Dysregulated mir-354 enhanced the protective response to nanopolystyrene by affecting the activity of TGF-β signaling pathway in nematode Caenorhabditis elegans. NanoImpact 2020, 20, 100256. [Google Scholar] [CrossRef]
- Liu, H.-L.; Zhao, Y.-Y.; Bi, K.; Rui, Q.; Wang, D.-Y. Dysregulated mir-76 mediated a protective response to nanopolystyrene by modulating heme homeostasis related molecular signaling in nematode Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2021, 212, 112018. [Google Scholar] [CrossRef]
- Qu, M.; Li, D.; Qiu, Y.-X.; Wang, D.-Y. Neuronal ERK MAPK signaling in response to low-dose nanopolystyrene exposure by suppressing insulin peptide expression in Caenorhabditis elegans. Sci. Total Environ. 2020, 724, 138378. [Google Scholar] [CrossRef]
- Qu, M.; Qiu, Y.-X.; Kong, Y.; Wang, D.-Y. Amino modification enhances reproductive toxicity of nanopolystyrene on gonad development and reproductive capacity in nematode Caenorhabditis elegans. Environ. Pollut. 2019, 254, 112978. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-H.; Wu, Q.-L.; Wang, D.-Y. Epigenetic response to nanopolystyrene in germline of nematode Caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2020, 206, 111404. [Google Scholar] [CrossRef]
- Wei, C.C.; Yen, P.L.; Chaikritsadakarn, A.; Huang, C.W.; Chang, C.H.; Liao, V.H. Parental CuO nanoparticles exposure results in transgenerational toxicity in Caenorhabditis elegans associated with possible epigenetic regulation. Ecotoxicol. Environ. Saf. 2020, 203, 111001. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.W.; Liao, V.H. Transgenerational reproductive effects of arsenite are associated with H3K4 dimethylation and SPR-5 downregulation in Caenorhabditis elegans. Environ. Sci. Technol. 2016, 50, 10673–10681. [Google Scholar] [CrossRef]
- Xu, R.-R.; Hua, X.; Rui, Q.; Wang, D.-Y. Alteration in Wnt signaling mediates induction of transgenerational toxicity of polystyrene nanoplastics in C. elegans. NanoImpact 2022, 28, 100425. [Google Scholar] [CrossRef]
- George, S.E.; Simokat, K.A.; Hardin, J.D.; Chisholm, A.D. The VAB-1 Eph receptor tyrosine kinase functions in neural and epithelial morphogenesis in C. elegans. Cell 1998, 92, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Ghosh-Roy, A.; Yanik, M.F.; Zhang, J.Z.; Jin, Y.; Chisholm, A.D. Caenorhabditis elegans neuronal regeneration is influenced by life stage, ephrin signaling, and synaptic branching. Proc. Natl. Acad. Sci. USA 2007, 104, 15132–15137. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, A.M.; Boudreau, J.R.; Yu, F.P.; Liu, J.; Chin-Sang, I.D. The Caenorhabditis elegans Eph receptor activates NCK and N-WASP, and inhibits Ena/VASP to regulate growth cone dynamics during axon guidance. PLoS Genet. 2012, 8, e1002513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrigan, C.; Subramanian, R.; Miller, M.A. Eph and NMDA receptors control Ca2+/calmodulin-dependent protein kinase II activation during C. elegans oocyte meiotic maturation. Development 2005, 132, 5225–5237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Johnson, R.W.; Park, D.; Chin-Sang, I.; Chamberlin, H.M. Somatic gonad sheath cells and Eph receptor signaling promote germ-cell death in C. elegans. Cell Death Differ. 2012, 19, 1080–1089. [Google Scholar] [CrossRef] [Green Version]
- Kipreos, E.T.; Gohel, S.P.; Hedgecock, E.M. The C. elegans F-box/WD-repeat protein LIN-23 functions to limit cell division during development. Development 2000, 127, 5071–5082. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.-X.; Cao, C.; Zhu, Y.-K.; Huang, W.; Yang, Y.; Qiu, H.-B.; Liu, S.-Q.; Wang, D.-Y. Beneficial effect of Xuebijing against Pseudomonas aeruginosa infection in Caenorhabditis elegans. Front. Pharmacol. 2022, 13, 949608. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Hua, X.; Bian, Q.; Wang, D. Nanoplastic Exposure at Predicted Environmental Concentrations Induces Activation of Germline Ephrin Signal Associated with Toxicity Formation in the Caenorhabditis elegans Offspring. Toxics 2022, 10, 699. https://doi.org/10.3390/toxics10110699
Zhao Y, Hua X, Bian Q, Wang D. Nanoplastic Exposure at Predicted Environmental Concentrations Induces Activation of Germline Ephrin Signal Associated with Toxicity Formation in the Caenorhabditis elegans Offspring. Toxics. 2022; 10(11):699. https://doi.org/10.3390/toxics10110699
Chicago/Turabian StyleZhao, Yue, Xin Hua, Qian Bian, and Dayong Wang. 2022. "Nanoplastic Exposure at Predicted Environmental Concentrations Induces Activation of Germline Ephrin Signal Associated with Toxicity Formation in the Caenorhabditis elegans Offspring" Toxics 10, no. 11: 699. https://doi.org/10.3390/toxics10110699
APA StyleZhao, Y., Hua, X., Bian, Q., & Wang, D. (2022). Nanoplastic Exposure at Predicted Environmental Concentrations Induces Activation of Germline Ephrin Signal Associated with Toxicity Formation in the Caenorhabditis elegans Offspring. Toxics, 10(11), 699. https://doi.org/10.3390/toxics10110699