
Developmental Phenotypic and Transcriptomic Effects of Exposure to Nanomolar Levels of 4-Nonylphenol, Triclosan, and Triclocarban in Zebrafish (Danio rerio)

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Animal Husbandry
2.2. Chemical Exposures
2.3. Abnormality Screening
2.4. Behavioral Analysis
2.5. Transcriptomics
3. Results
3.1. Triclosan
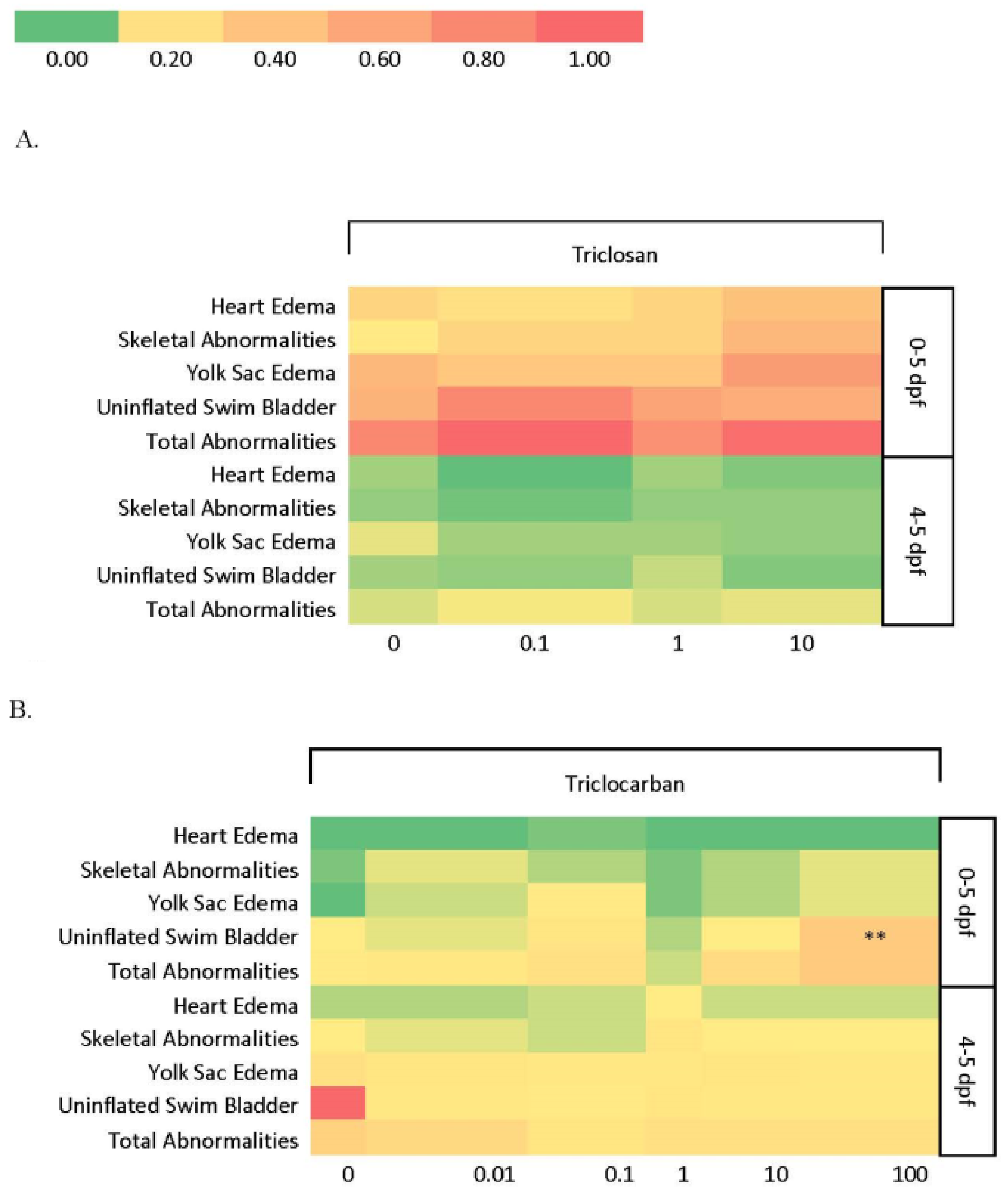
3.1.1. Larval Abnormalities and Mortality
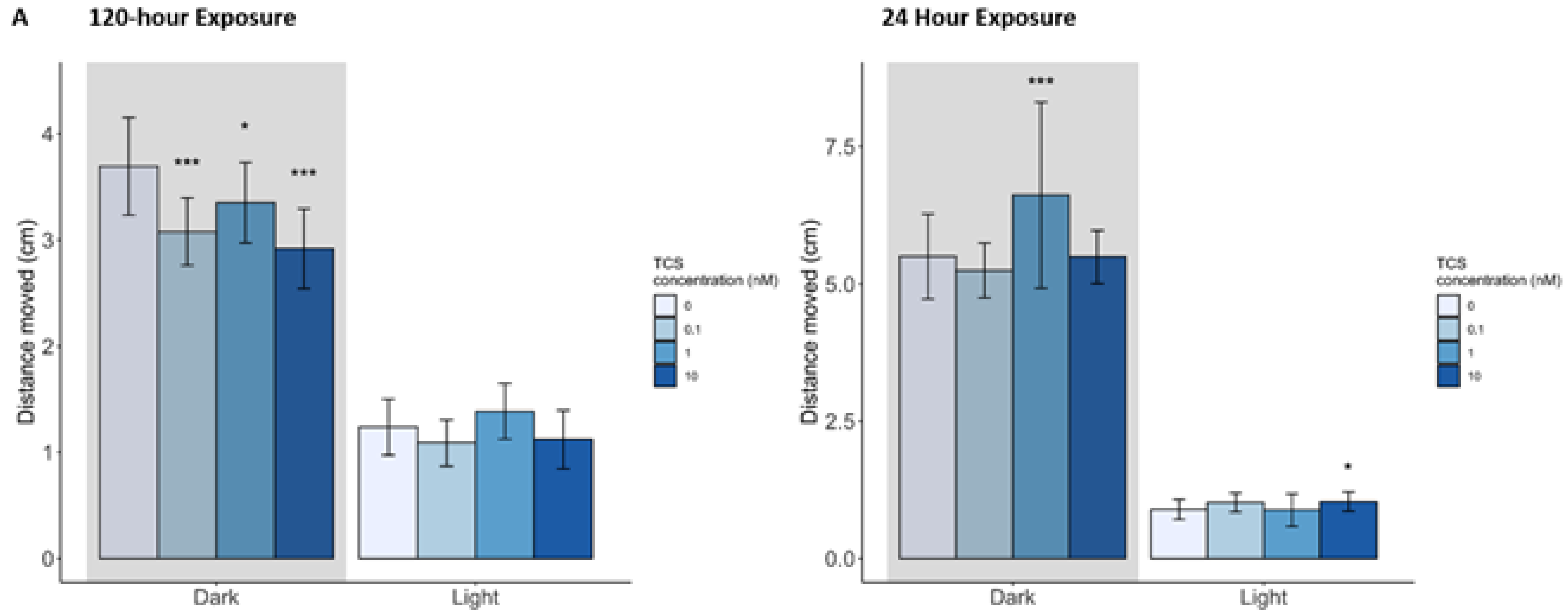
3.1.2. Behavior
3.1.3. Gene Expression and Pathway Analysis
3.2. Triclocarban
3.2.1. Larval Abnormalities and Mortality
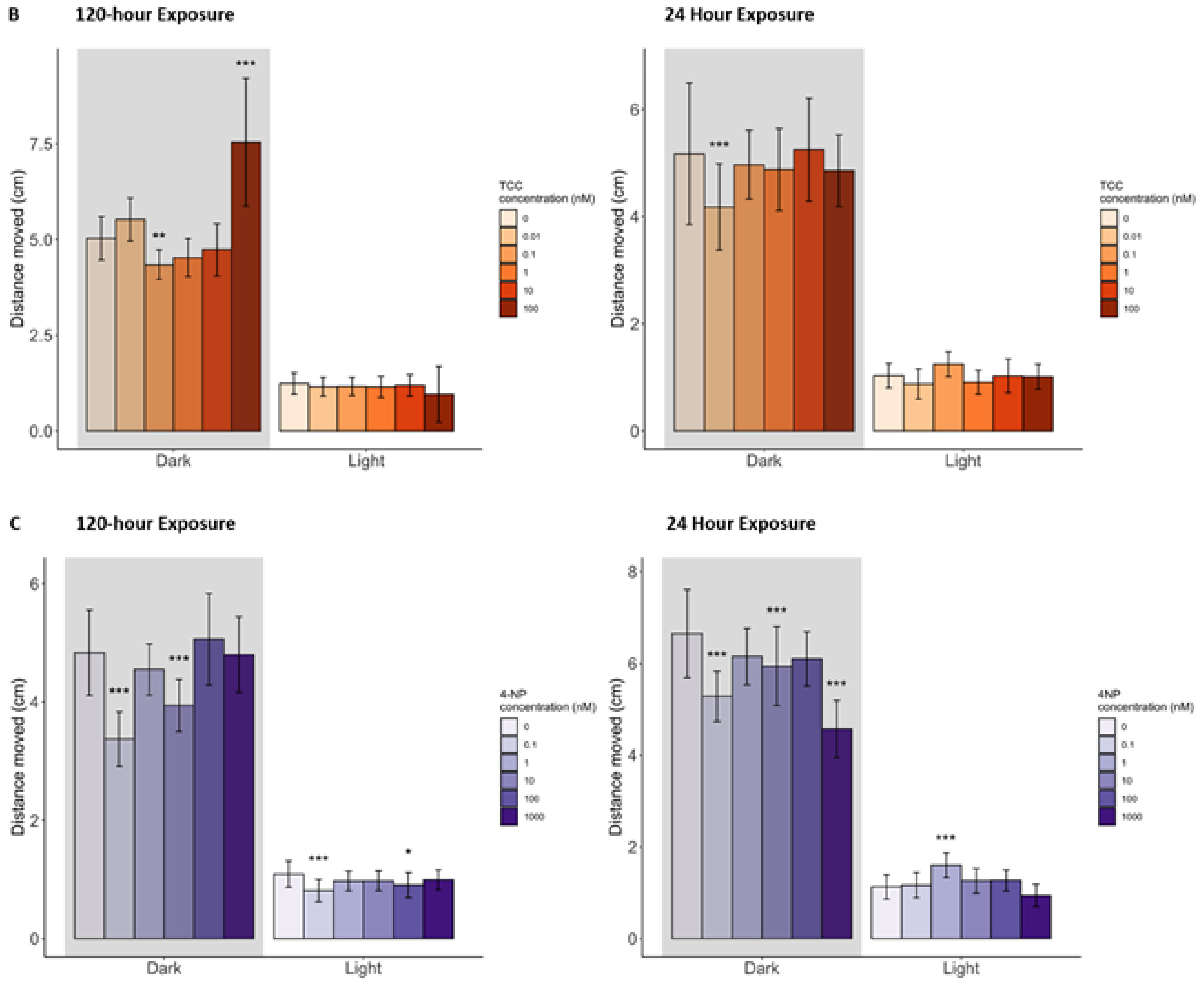
3.2.2. Behavior
3.2.3. Gene Expression and Pathway Analysis
3.3. 4-nonylphenol
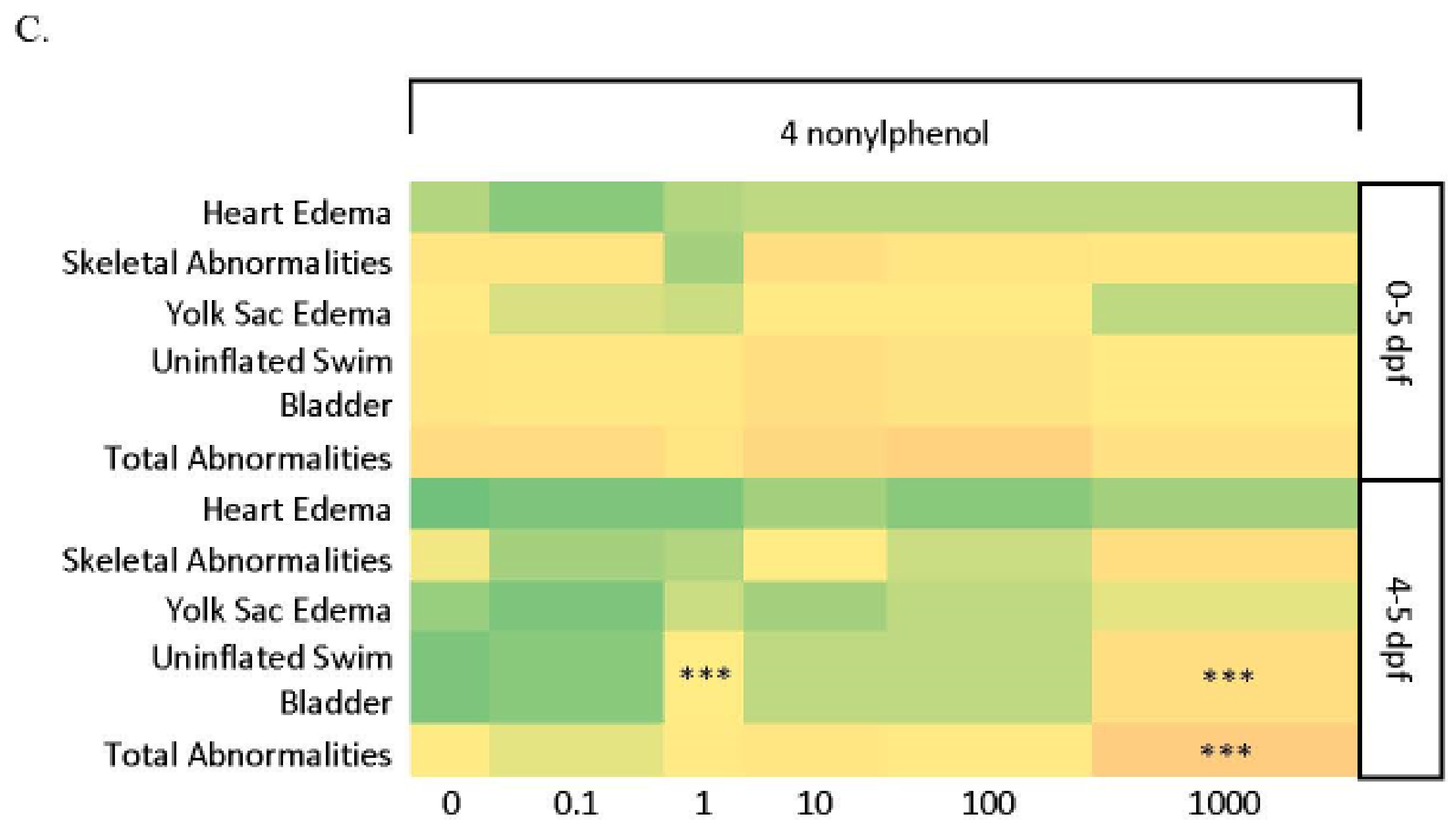
3.3.1. Larval Abnormalities and Mortality
3.3.2. Behavior
3.3.3. Gene Expression and Pathway Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NIEHS. National Institute of Environmental Health Sciences. Endocrine Disruptors. Available online: https://www.niehs.nih.gov/health/topics/agents/endocrine/index.cfm (accessed on 22 December 2021).
- Halden, R.U.; Lindeman, A.E.; Aiello, A.E.; Andrews, D.; Arnold, W.A.; Fair, P.; Fuoco, R.E.; Geer, L.A.; Johnson, P.I.; Lohmann, R.; et al. The florence statement on triclosan and triclocarban. Environ. Health Perspect. 2017, 125, 064501. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.B.; Haimbaugh, A.S.; Sperone, F.G.; Johnson, D.M.; Baker, T.R. Persistent contaminants of emerging concern in a great lakes urban-dominant watershed. J. Great Lakes Res. 2021. [Google Scholar] [CrossRef]
- Weatherly, L.M.; Gosse, J.A. Triclosan exposure, transformation, and human health effects. J. Toxicol. Environ. Health. Part B Crit. Rev. 2017, 20, 447–469. [Google Scholar] [CrossRef] [PubMed]
- Halden, R.U.; Paull, D.H. Co-occurrence of triclocarban and triclosan in U.S. water resources. Environ. Sci. Technol. 2002, 39, 1420–1426. [Google Scholar] [CrossRef] [PubMed]
- Pycke, B.F.G.; Geer, L.A.; Dalloul, M.; Abulafia, O.; Jenck, A.M.; Halden, R.U. Human fetal exposure to triclosan and triclocarban in an urban population from Brooklyn, New York. Environ. Sci. Technol. 2014, 48, 8831–8838. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhang, P.; Zhang, Z.; Shi, J.; Jiao, Z.; Shao, B. Endocrine Disrupting Effects of Triclosan on the Placenta in Pregnant Rats. PLoS ONE 2016, 11, e0154758. [Google Scholar] [CrossRef] [Green Version]
- Radwan, P.; Wielgomas, B.; Radwan, M.; Krasiński, R.; Klimowska, A.; Zajdel, R.; Kaleta, D.; Jurewicz, J. Triclosan exposure and in vitro fertilization treatment outcomes in women undergoing in vitro fertilization. Environ. Sci. Pollut. Res. Int. 2021, 28, 12993–12999. [Google Scholar] [CrossRef]
- James, M.O.; Li, W.; Summerlot, D.P.; Rowland-Faux, L.; Wood, C.E. Triclosan is a potent inhibitor of estradiol and estrone sulfonation in sheep placenta. Environ. Int. 2010, 36, 942–949. [Google Scholar] [CrossRef] [Green Version]
- Vélez, M.P.; Arbuckle, T.E.; Fraser, W.D. Female exposure to phenols and phthalates and time to pregnancy: The Maternal-Infant Research on Environmental Chemicals (MIREC) Study. Fertil. Steril. 2015, 103, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Koeppe, E.S.; Ferguson, K.K.; Colacino, J.A.; Meeker, J.D. Relationship between urinary triclosan and paraben concentrations and serum thyroid measures in NHANES 2007–2008. Sci. Total Environ. 2013, 445–446, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Hinther, A.; Bromba, C.M.; Wulff, J.E.; Helbing, C.C. Effects of triclocarban, triclosan, and methyl triclosan on thyroid hormone action and stress in frog and mammalian culture systems. Environ. Sci. Technol. 2011, 45, 5395–5402. [Google Scholar] [CrossRef] [PubMed]
- Christen, V.; Crettaz, P.; Oberli-Schrämmli, A.; Fent, K. Some flame retardants and the antimicrobials triclosan and triclocarban enhance the androgenic activity in vitro. Chemosphere 2010, 81, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Villeneuve, D.L.; Jensen, K.M.; Cavallin, J.E.; Durhan, E.J.; Garcia-Reyero, N.; Kahl, M.D.; Ankley, G.T. Effects of the antimicrobial contaminant triclocarban, and co-exposure with the androgen 17β-trenbolone, on reproductive function and ovarian transcriptome of the fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2017, 36, 231–242. [Google Scholar] [CrossRef] [PubMed]
- High Priority Chemicals Data System (HPCDS). 2020. Available online: https://hpcds.theic2.org/ (accessed on 14 November 2021).
- Kourouma, A.; Keita, H.; Duan, P.; Quan, C.; Bilivogui, K.K.; Qi, S.; Yang, K. Effects of 4-nonylphenol on oxidant/antioxidant balance system inducing hepatic steatosis in male rat. Toxicol. Rep. 2015, 2, 1423–1433. [Google Scholar] [CrossRef] [Green Version]
- Bistakova, J.; Forgacs, Z.; Bartos, Z.; Szivosne, M.R.; Jambor, T.; Knazicka, Z.; Lukac, N. Effects of 4-nonylphenol on the steroidogenesis of human adrenocarcinoma cell line (NCI-H295R). J. Environ. Sci. Health Part A 2017, 52, 221–227. [Google Scholar] [CrossRef]
- Duan, P.; Hu, C.; Butler, H.J.; Quan, C.; Chen, W.; Huang, W.; Yang, K. Effects of 4-nonylphenol on spermatogenesis and induction of testicular apoptosis through oxidative stress-related pathways. Reprod. Toxicol. 2016, 62, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Mao, Z.; Zheng, X.; Zhang, Y.; Tao, X.; Li, Y.; Wang, W. Occurrence and Biodegradation of Nonylphenol in the Environment. Int. J. Mol. Sci. 2012, 13, 491–505. [Google Scholar] [CrossRef]
- Hua, W.; Bennett, E.R.; Letcher, R.J. Triclosan in waste and surface waters from the upper Detroit River by liquid chromatography-electrospray-tandem quadrupole mass spectrometry. Environ. Int. 2005, 31, 621–630. [Google Scholar] [CrossRef]
- Shen, J.Y.; Chang, M.S.; Yang, S.; Wu, G.J. Simultaneous determination of triclosan, triclocarban, and transformation products of triclocarban in aqueous samples using solid-phase micro-extraction-HPLC-MS/MS. J. Sep. Sci. 2012, 35, 2544–2552. [Google Scholar] [CrossRef]
- Gilroy, È.A.M.; Muir, D.C.G.; McMaster, M.E.; Darling, C.; Campbell, L.M.; Alaee, M.; Sherry, J.P. Halogenated phenolic compounds in wild fish from Canadian Areas of Concern. Environ. Toxicol. Chem. 2017, 36, 2266–2273. [Google Scholar] [CrossRef]
- Blair, B.D.; Crago, J.P.; Hedman, C.J.; Klaper, R.D. Pharmaceuticals and personal care products found in the Great Lakes above concentrations of environmental concern. Chemosphere 2013, 93, 2116–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vimalkumar, K.; Arun, E.; Krishna-Kumar, S.; Poopal, R.K.; Nikhil, N.P.; Subramanian, A.; Babu-Rajendran, R. Occurrence of triclocarban and benzotriazole ultraviolet stabilizers in water, sediment, and fish from Indian rivers. Sci. Total Environ. 2018, 625, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing [Computer Software Manual]; R Core Team: Vienna, Austria, 2016. [Google Scholar]
- Wirt, H.; Botka, R.; Perez, K.E.; King-Heiden, T. Embryonic exposure to environmentally relevant concentrations of triclosan impairs foraging efficiency in zebrafish larvae. Environ. Toxicol. Chem. 2018, 37, 3124–3133. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.; Domingues, I.; Grisolia, C.K.; Soares, A.M.V.M. Effects of triclosan on zebrafish early-life stages and adults. Environ. Sci. Pollut. Res. 2009, 16, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.C.V.R.; Araújo, C.V.M.; López-Doval, J.C.; Neto, M.B.; Silva, F.T.; Paiva, T.C.B.; Pompêo, M.L.M. Potential effects of triclosan on spatial displacement and local population decline of the fish Poecilia reticulata using a non-forced system. Chemosphere 2017, 184, 329–336. [Google Scholar] [CrossRef]
- Horie, Y.; Yamagishi, T.; Takahashi, H.; Iguchi, T.; Tatarazako, N. Effects of triclosan on Japanese medaka (Oryzias latipes) during embryo development, early life stage and reproduction. J. Appl. Toxicol. 2018, 38, 544–551. [Google Scholar] [CrossRef]
- Salierno, J.D.; Lopes, M.; Rivera, M. Latent effects of early life stage exposure to triclosan on survival in fathead minnows, Pimephales promelas. J. Environ. Sci. Health 2016, 51, 695–702. [Google Scholar] [CrossRef]
- Ishibashi, H.; Matsumura, N.; Hirano, M.; Matsuoka, M.; Shiratsuchi, H.; Ishibashi, Y.; Arizono, K. Effects of triclosan on the early life stages and reproduction of medaka Oryzias latipes and induction of hepatic vitellogenin. Aquat. Toxicol. 2004, 67, 167–179. [Google Scholar] [CrossRef]
- Macedo, S.; Torres, T.; Santos, M.M. Methyl-triclosan and triclosan impact embryonic development of Danio rerio and Paracentrotus lividus. Ecotoxicology 2017, 26, 482–489. [Google Scholar] [CrossRef]
- Falisse, E.; Voisin, A.-S.; Silvestre, F. Impacts of triclosan exposure on zebrafish early-life stage: Toxicity and acclimation mechanisms. Aquat. Toxicol. 2017, 189, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Li, J.; Wang, X.; Gao, X.; Chen, J.; Ai, S.; Li, W.; Huang, Y.; Liu, Z. Study of aquatic life criteria and ecological risk assessment for triclocarban (TCC). Environ. Pollut. 2019, 254, 112956. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Xu, H.; Shen, Y.; Qiu, W.; Yang, M. Oxidative stress in zebrafish embryos induced by short-term exposure to bisphenol A, nonylphenol, and their mixture. Environ. Toxicol. Chem. 2011, 30, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Sano, K.; Inohaya, K.; Kawaguchi, M.; Yoshizaki, N.; Iuchi, I.; Yasumasu, S. Purification and characterization of zebrafish hatching enzyme—An evolutionary aspect of the mechanism of egg envelope digestion. FEBS J. 2008, 275, 5934–5946. [Google Scholar] [CrossRef] [PubMed]
- Willi, R.A.; Faltermann, S.; Hettich, T.; Fent, K. Active Glucocorticoids Have a Range of Important Adverse Developmental and Physiological Effects on Developing Zebrafish Embryos. Environ. Sci. Technol. 2018, 52, 877–885. [Google Scholar] [CrossRef]
- Liang, X.; Souders, C.L., 2nd; Zhang, J.; Martyniuk, C.J. Tributyltin induces premature hatching and reduces locomotor activity in zebrafish (Danio rerio) embryos/larvae at environmentally relevant levels. Chemosphere 2017, 189, 498–506. [Google Scholar] [CrossRef]
- Pullaguri, N.; Nema, S.; Bhargava, Y.; Bhargava, A. Triclosan alters adult zebrafish behavior and targets acetylcholinesterase activity and expression. Environ. Toxicol. Pharmacol. 2020, 75, 103311. [Google Scholar] [CrossRef]
- Poon, K.L.; Wang, X.; Lee, S.G.P.; Ng, A.S.; Goh, W.H.; Zhao, Z.; Al-Haddawi, M.; Wang, H.; Mathavan, S.; Ingham, P.W.; et al. Editor’s Highlight: Transgenic Zebrafish Reporter Lines as Alternative In Vivo Organ Toxicity Models. Toxicol. Sci. 2017, 156, 133–148. [Google Scholar] [CrossRef]
- Van Sebille, Y.Z.; Gibson, R.J.; Wardill, H.R.; Carney, T.J.; Bowen, J.M. Highlight article: Use of zebrafish to model chemotherapy and targeted therapy gastrointestinal toxicity. Exp. Biol. Med. 2019, 244, 1178–1185. [Google Scholar] [CrossRef]
- Lin, C.; Chou, P.; Chen, P. Two azole fungicides (carcinogenic triadimefon and non-carcinogenic myclobutanil) exhibit different hepatic cytochrome P450 activities in medaka fish. J. Hazard. Mater. 2014, 277, 150–158. [Google Scholar] [CrossRef]
- Huang, H.; Zhu, J.; Li, Y.; Zhang, L.; Gu, J.; Xie, Q.; Jin, H.; Che, X.; Li, J.; Huang, C.; et al. Upregulation of SQSTM1/p62 contributes to nickel-induced malignant transformation of human bronchial epithelial cells. Autophagy 2016, 12, 1687–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, J.J.; Archer, S.L. Emerging concepts in the molecular basis of pulmonary arterial hypertension (PAH): Part I: Metabolic plasticity and mitochondrial dynamics in the pulmonary circulation and right ventricle in PAH. Circulation 2015, 131, 1691–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cass, A.N.; Servetnick, M.D.; McCune, A.R. Expression of a lung developmental cassette in the adult and developing zebrafish swimbladder: Swimbladder gene expression. Evol. Dev. 2013, 15, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, Z.; Collins, J.E.; Andrews, R.M.; Stemple, D.; Gong, Z. Comparative transcriptome analyses indicate molecular homology of zebrafish swimbladder and mammalian lung. PloS One 2011, 6, e24019. [Google Scholar] [CrossRef] [Green Version]
- Winata, C.L.; Korzh, S.; Kondrychyn, I.; Korzh, V.; Gong, Z. The role of vasculature and blood circulation in zebrafish swimbladder development. BMC Dev. Biol. 2010, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Kumaran, S.S.; Kavitha, C.; Ramesh, M.; Grummt, T. Toxicity studies of nonylphenol and octylphenol: Hormonal, hematological and biochemical effects in clarias gariepinus. J. Appl. Toxicol. 2011, 31, 752–761. [Google Scholar] [CrossRef]
- Ogungbemi, A.; Leuthold, D.; Scholz, S.; Küster, E. Hypo- or hyperactivity of zebrafish embryos provoked by neuroactive substances: A review on how experimental parameters impact the predictability of behavior changes. Environ. Sci. Eur. 2019, 31, 1–26. [Google Scholar] [CrossRef]
- Burgess, H.A.; Granato, M. Modulation of locomotor activity in larval zebrafish during light adaptation. J. Exp. Biol. 2007, 210 Pt 14, 2526–2539. [Google Scholar] [CrossRef] [Green Version]
- Tabari, S.A.; Esfahani, M.L.; Hosseini, S.M.; Rahimi, A. Neurobehavioral toxicity of triclosan in mice. Food Chem. Toxicol. 2019, 130, 154–160. [Google Scholar] [CrossRef]
- Sahu, V.K.; Karmakar, S.; Kumar, S.; Shukla, S.P.; Kumar, K. Triclosan toxicity alters behavioral and hematological parameters and vital antioxidant and neurological enzymes in Pangasianodon hypophthalmus (Sauvage, 1878). Aquat. Toxicol. 2018, 202, 145–152. [Google Scholar] [CrossRef]
- Brown, J.; Bernot, M.J.; Bernot, R.J. The influence of TCS on the growth and behavior of the freshwater snail, Physa acuta. J. Environ. Sci. Health 2012, 47, 1626–1630. [Google Scholar] [CrossRef] [PubMed]
- Nassef, M.; Matsumoto, S.; Seki, M.; Khalil, F.; Kang, I.J.; Shimasaki, Y.; Honjo, T. Acute effects of triclosan, diclofenac and carbamazepine on feeding performance of Japanese medaka fish (Oryzias latipes). Chemosphere 2010, 80, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, E.B.; Connon, R.E.; Werner, I.; Davies, R.E.; Beggel, S.; Feng, W.; Pessah, I.N. Triclosan impairs swimming behavior and alters expression of excitation-contraction coupling proteins in fathead minnow (Pimephales promelas). Environ. Sci. Technol. 2013, 47, 2008–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etzel, T.; Muckle, G.; Arbuckle, T.E.; Fraser, W.D.; Ouellet, E.; Seguin, J.R.; Braun, J.M. Prenatal urinary triclosan concentrations and child neurobehavior. Environ. Int. 2018, 114, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sun, L.; Zhang, H.; Shi, M.; Dahlgren, R.A.; Wang, X.; Wang, H. Response mechanisms to joint exposure of triclosan and its chlorinated derivatives on zebrafish (Danio rerio) behavior. Chemosphere 2018, 193, 820–832. [Google Scholar] [CrossRef] [Green Version]
- Jackson-Browne, M.S.; Papandonatos, G.D.; Chen, A.; Yolton, K.; Lanphear, B.P.; Braun, J.M. Early-life triclosan exposure and parent-reported behavior problems in 8-year-old children. Environ. Int. 2019, 128, 446–456. [Google Scholar] [CrossRef]
- Ward, A.J.W.; Duff, A.J.; Currie, S. The effects of the endocrine disrupter 4-nonylphenol on the behaviour of juvenile rainbow trout (Oncorhynchus mykiss). Can. J. Fish. Aquat. Sci. 2006, 63, 377–382. [Google Scholar] [CrossRef]
- Sharma, M.; Chadha, P.; Borah, M.K. Fish Behaviour and Immune Response as a Potential Indicator of Stress Caused by 4-Nonylphenol. Am. J. BioScience 2015, 3, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekar, G.; Arner, A.; Kitambi, S.S.; Dahlman-Wright, K.; Lendahl, M.A. Developmental toxicity of the environmental pollutant 4-nonylphenol in zebrafish. Neurotoxicology Teratol. 2011, 33, 752–764. [Google Scholar] [CrossRef]
- Mao, Z.; Zheng, Y.L.; Zhang, Y.Q. Behavioral impairment and oxidative damage induced by chronic application of nonylphenol. Int. J. Mol. Sci. 2011, 12, 114–127. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Niu, C.; Pei, X. Effects of chronic exposure to nonylphenol on locomotor activity and social behavior in zebrafish (Danio rerio). J. Environ. Sci. 2010, 22, 1435–1440. [Google Scholar] [CrossRef]
- Yokota, H.; Seki, M.; Maeda, M.; Oshima, Y.; Tadokoro, H.; Honjo, T.; Kobayashi, K. Life-Cycle Toxicity of 4-Nonylphenol To Medaka (Oryzias Latipes). Environ. Toxicol. Chem. 2001, 20, 2552–2560. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M.M.; Bartell, S.E.; Schoenfuss, H.L. Effects of triclosan and triclocarban, two ubiquitous environmental contaminants, on anatomy, physiology, and behavior of the fathead minnow (Pimephales promelas). Arch. Environ. Contam. Toxicol. 2012, 63, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Barros, S.; Montes, R.; Quintana, J.B.; Rodil, R.; Oliveira, J.M.A.; Santos, M.M.; Neuparth, T. Chronic effects of triclocarban in the amphipod Gammarus locusta: Behavioural and biochemical impairment. Ecotoxicol. Environ. Saf. 2017, 135, 276–283. [Google Scholar] [CrossRef]
- Suzuki, M.; Shimizu, T. Is SPINK1 gene mutation associated with development of pancreatic cancer? New insight from a large retrospective study. Ebiomedicine 2019, 50, 5–6. [Google Scholar] [CrossRef]
- Räsänen, K.; Itkonen, O.; Koistinen, H.; Stenman, U. Emerging roles of SPINK1 in cancer. Clin. Chem. 2016, 62, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Zhou, X.; Han, X.; Zhang, Y.; Jiang, L. SPINK1 is a prognosis predicting factor of non-small cell lung cancer and regulates redox homeostasis. Oncol. Lett. 2019, 18, 6899–6908. [Google Scholar] [CrossRef] [Green Version]
- Ajao, C.; Andersson, M.A.; Teplova, V.V.; Nagy, S.; Gahmberg, C.G.; Andersson, L.C.; Hautaniemi, M.; Kakasi, B.; Roivainen, M.; Salkinoja-Salonen, M. Mitochondrial toxicity of triclosan on mammalian cells. Toxicol. Rep. 2015, 2, 624–637. [Google Scholar] [CrossRef] [Green Version]
- Dinwiddie, M.T.; Terry, P.D.; Chen, J. Recent evidence regarding triclosan and cancer risk. Int. J. Environ. Res. Public Health 2014, 11, 2209–2217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Sha, W.; Liu, Z.; Chi, C. Effect of chymotrypsin C and related proteins on pancreatic cancer cell migration. Acta Biochim. Biophys. Sin. 2011, 43, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Casals-Casas, C.; Desvergne, B. Endocrine disruptors: From endocrine to metabolic disruption. Annu. Rev. Physiol. 2011, 73, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
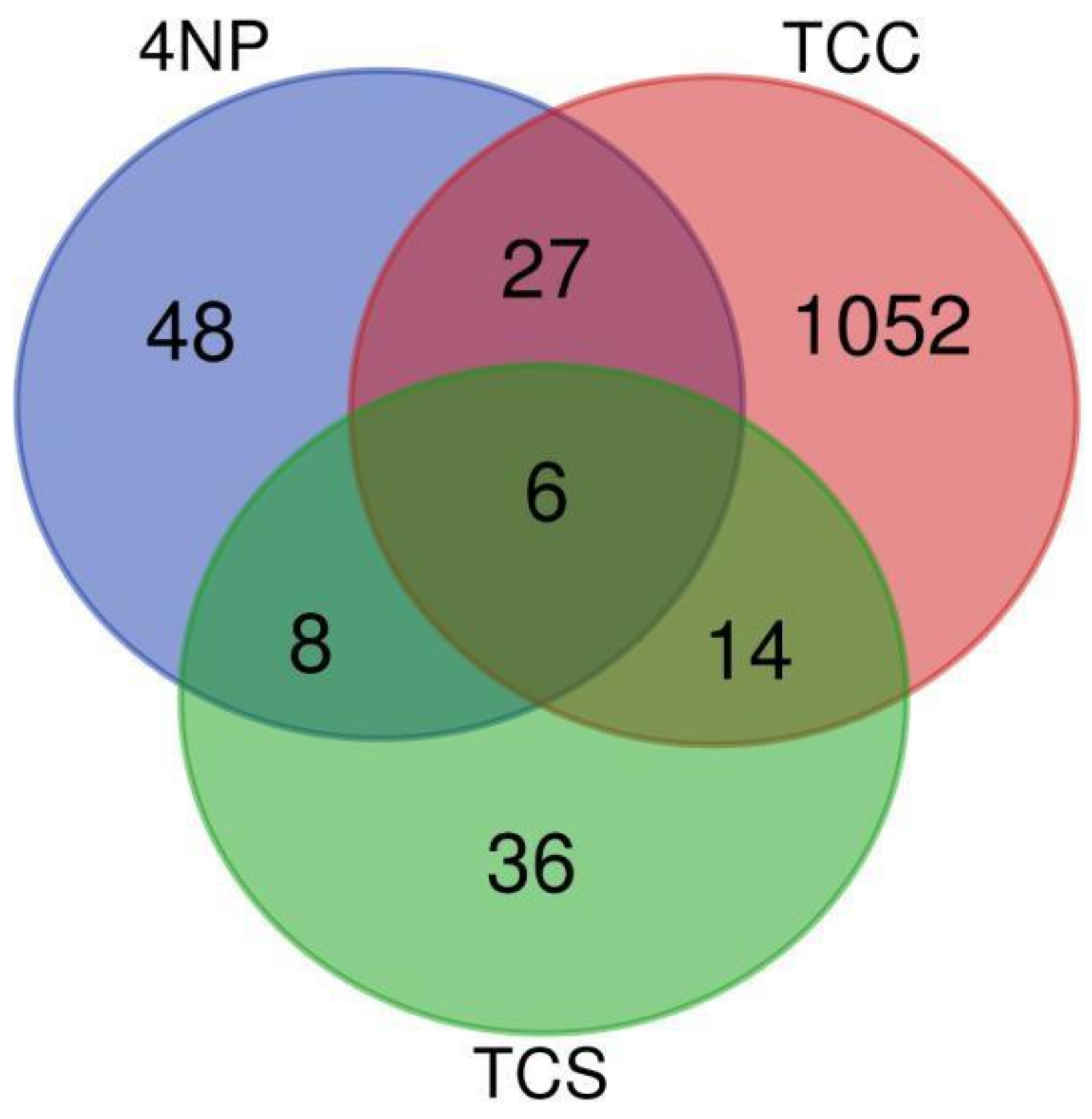{kind=link}
| Triclosan | ||||
|---|---|---|---|---|
| 0.1 nM | 1 nM | 10 nM | Total genes | |
| 24 h | 7 ↓, 15 ↑ | 2 ↓, 17 ↑ | 1 ↓, 3 ↑ | 10↓ 35↑ |
| 120 h | 11 ↓, 8 ↑ | 0 ↓, 1 ↑ | 3 ↓, 8 ↑ | 14 ↓ 17↑ |
| Triclocarban | ||||
| 0.01 nM | 1 nM | 100 nM | Total genes | |
| 24 h | 115 ↓, 54 ↑ | 26 ↓,37 ↑ | 8 ↓, 9 ↑ | 149↓ 100↑ |
| 120 h | 0 ↓, 1 ↑ | 578 ↓, 258 ↑ | 478 ↓, 465 ↑ | 1056↓ 724↑ |
| 4-nonylphenol | ||||
| 0.01 nM | 10 nM | 1000 nM | Total genes | |
| 24 h | 0 ↓, 0 ↑ | 4 ↓, 34 ↑ | 8↓, 6 ↑ | 12 ↓ 40↑ |
| 120 h | 32 ↓, 7 ↑ | 1↓, 0 ↑ | 1 ↓, 0 ↑ | 33 ↓ 7↑ |
| Exposure Concentrations (nM) 0–5 Days | Exposure Concentrations (nM) 4–5 Days | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene Name | 4-nonylphenol | Triclosan | Triclocarban | 4-nonylphenol | Triclosan | Triclocarban | |||||||||||||
| 0.1 | 10 | 1000 | 0.1 | 1 | 10 | 0.01 | 1 | 100 | 0.1 | 10 | 1000 | 0.1 | 1 | 10 | 0.01 | 1 | 100 | ||
| Cardiovascular | |||||||||||||||||||
| fbxo32 | F-box protein 32 | −1.2 | −0.8 | −0.2 | 0.4 | 0.2 | 1 | −0.01 | 0.5 | 1.4 | 0.6 | 0.2 | −0.5 | 1.1 | 0.3 | 0.1 | 1.7 | −1.1 | −0.2 |
| tfr1b | transferrin receptor 1b | 0.4 | 0.2 | 0.3 | 0.1 | −0.1 | 0.2 | −0.03 | −0.1 | −1.2 | 0.6 | 0.8 | −0.2 | 0.2 | 0.1 | −0.4 | −0.9 | ||
| myl10 | myosin, light chain 10, regulatory | −0.1 | 0.04 | −0.3 | −0 | 0.1 | −0 | −0.2 | 0.2 | −0.1 | 0.3 | 0.4 | 0.8 | 0.2 | 0.2 | −0.1 | −0.8 | −0.7 | 0.1 |
| mat2aa | methionine adenosyltransferase II, alpha a | −0.1 | 0.1 | 0.02 | −0 | −0 | 0.4 | −0.01 | −0.5 | −1 | −0.1 | 0.2 | 0.9 | −0.1 | 0.2 | 0.3 | −0.1 | 0.2 | 0.05 |
| hbae1.1 | hemoglobin, alpha embryonic 1.1 | −0.1 | 0.01 | −0.3 | −0.8 | −0.3 | −0.3 | 0.1 | −0.6 | −0.4 | −0.4 | −0.2 | 0.9 | −0.6 | 1.1 | 1 | −0.7 | −1.6 | −0.2 |
| tcp1 | t-complex 1 | 0.3 | −0.2 | −0.1 | −0.9 | −0.6 | −0.2 | 0.2 | −1.2 | −0.2 | −0.1 | −0.3 | 0.2 | −1 | 0.3 | 0.3 | −0.3 | −0.1 | −0.2 |
| hbbe1.2 | hemoglobin beta embryonic- 1.2 | 0.2 | 0.3 | 0.04 | ND | −0.3 | −0.4 | 0.04 | −0.1 | −0.3 | −0 | −0.2 | 0.03 | −0.1 | 1.1 | 0.4 | −0.3 | −0.8 | −0.8 |
| Neurological | |||||||||||||||||||
| reep3b | receptor accessory protein 3b | 0.5 | 0.7 | 0.2 | 0.6 | 0.03 | 0.2 | −0.4 | 0.2 | −0.2 | 0.5 | 0.8 | 0.2 | 0.8 | 0.1 | 0.01 | −0.3 | −1.4 | 0.1 |
| syngr3b | synaptogyrin 3b | −0.03 | 0.7 | 0.3 | 0.9 | 0.1 | 0.2 | −0.1 | 0.3 | −0.3 | 0.5 | 1 | 0.1 | 0.9 | 0.1 | −0.1 | 0.3 | −0.2 | −0.04 |
| agr2 | anterior gradient 2 | 0.9 | 0.4 | −0.2 | −0.5 | −0.4 | −0.3 | 0.2 | −0.7 | −0.8 | −0.1 | −0.05 | 0.4 | −0.3 | 0.4 | 0.05 | −0.7 | −2.5 | −0.1 |
| crygm2d18 | crystallin, gamma M2d18 | −0.2 | −1 | −0.1 | 0.2 | 0.1 | −0.8 | 0.03 | 0.04 | −0.1 | −0.3 | 0.3 | −0.1 | −0.6 | 0.6 | 0.1 | −0.2 | −1 | 0.8 |
| elmo2 | engulfment and cell motility 2 | −0.2 | 0.4 | −0.1 | 0.2 | −0 | 0 | −0.3 | −0 | −0.2 | 0.4 | 0.8 | −0.04 | 0.4 | 0.5 | −0.1 | 0.5 | 1.4 | 0.02 |
| arr3b | arrestin 3b, retinal (X- arrestin) | 0.1 | 0.4 | −0.02 | −0.1 | 0.2 | 0.1 | −0.3 | −0 | −2.6 | 0.2 | 0.8 | −0.2 | 0.1 | −0.1 | −0.2 | 0.04 | −0.4 | 0.2 |
| opn1sw2 | opsin 1 (cone pigments), short-wave-sensitive 2 | −0.1 | −0.01 | 0.2 | −0.2 | −0.3 | −0.8 | −0.1 | −0.1 | −0.3 | −0.3 | −0.4 | 0.2 | −0.4 | −0.4 | −0.3 | −0.8 | −1.3 | −0.1 |
| si:ch211- 153b23.5 | si:ch211-153b23.5 | 0.9 | 0.3 | −0.2 | −1 | −0.9 | −0.6 | 0.1 | −0.9 | 0.6 | 0.7 | 0.5 | 1 | −0.8 | 0.8 | 0.1 | −0.4 | 0.4 | 0.7 |
| mgst3a | microsomal glutathione S- transferase 3a | 0.3 | −0.1 | 0.1 | −0.9 | −0.9 | −0.3 | 0.2 | −1.1 | −0.5 | −0.5 | −0.5 | −0.1 | −1 | 0.4 | 0.3 | 0.02 | −0.5 | −0.4 |
| rlbp1b | retinaldehyde binding protein 1b | 0.1 | 0.1 | 0.4 | −0 | −0 | 0.2 | 0.2 | 0.1 | −0.8 | 0.2 | −0.1 | 0.1 | 0.4 | 1.5 | 0.5 | −1.1 | −0.5 | 0.04 |
| Metabolic Processes | |||||||||||||||||||
| cel.1 | carboxyl ester lipase, tandem duplicate 1 | −1.2 | −0.7 | −0.3 | 0.6 | 0.4 | 0.2 | −0.3 | 0.5 | 0.4 | 0.1 | −0.3 | −0.5 | 1 | 0.2 | −0.3 | 0.05 | −0.6 | −0.5 |
| amy2a | amylase alpha 2A | −0.9 | −0.7 | −0.1 | 0.8 | 0.4 | 0.2 | −0.1 | −0.2 | 0.5 | 0.1 | −0.2 | −0.8 | 0.8 | 0.4 | −0.2 | 0.4 | 0.4 | −0.6 |
| prss59.2 | serine protease 59, tandem duplicate 2 | −0.8 | −0.4 | 0.04 | 0.3 | −0.1 | 0.2 | −0.1 | −1 | 0.1 | −0 | −0.2 | −0.4 | 0.4 | 1.1 | 0.2 | 0.1 | −0.3 | −0.4 |
| ela2l | elastase 2 like | −0.7 | −0.3 | 0.06 | 0.3 | −0.5 | −0.6 | 0.2 | −0.2 | −0.1 | −0.5 | −0.9 | −0.4 | 0.5 | 0.4 | −0.9 | 0.1 | −0.8 | −0.3 |
| pip5k1cb | phosphatidylinositol-4- phosphate 5-kinase, type I, gamma b | 0.4 | 0.7 | 0.4 | 0.6 | 0.3 | 0 | −0.4 | 0 | −0.2 | 0.5 | 0.9 | 0.1 | 0.8 | −0.2 | −0.3 | 0.7 | −0.3 | 0.4 |
| fabp10a | fatty acid binding protein 10a, liver basic | −0.9 | −0.1 | −0.1 | 0.2 | 0.01 | 0.3 | −0.1 | 0.04 | −0.1 | −0.1 | −0.02 | −0.7 | 0.4 | 0.6 | 0.2 | 0.4 | −0.9 | −0.2 |
| zgc:92590 | zgc:92590 | −0.9 | −0.3 | −1.1 | −0 | −0 | 0 | −0.1 | −0.3 | −1.6 | −0.1 | 0.2 | −0.2 | 0.1 | 0.8 | 0.1 | −0.03 | −1.1 | −0.9 |
| si:ch211- 234p6.10 | si:ch211-234p6.10 | −0.8 | −0.03 | 0.4 | 0 | −0.2 | 0.2 | −0.04 | 0.1 | −0.3 | 0.4 | 0.1 | −0.6 | 0.3 | 0.6 | 0.2 | 0.9 | 0.3 | −0.4 |
| lpin1a | lipin 1 | −0.8 | −0.4 | −0.4 | 0.1 | −0 | 0.2 | −0.2 | 0.2 | 0.9 | 0.2 | −0 | −0.3 | 0.4 | 0.02 | −0.1 | 0.6 | 0.2 | −0.03 |
| fkbp9 | FKBP prolyl isomerase 9 | 0.8 | 0.6 | −0.02 | −0.1 | −0.3 | 0 | −0.4 | −1.7 | −2 | −0 | 1 | 0.8 | −0.2 | −0.2 | 0.04 | −0.1 | −0.6 | −0.1 |
| abcc2 | ATP-binding cassette, sub- family C (CFTR/MRP), member 2 | −0.4 | 0.1 | −0.1 | 0.4 | 1 | 1 | −0.1 | 0.4 | 1 | 0.3 | 0.2 | 0.2 | 0.7 | −0.2 | 0.6 | −0.1 | 0.2 | −0.1 |
| plin2 | perilipin 2 | 0.06 | −0.2 | 0.6 | −0 | 0.1 | 1.1 | −0.2 | −0.3 | 1.1 | 0.8 | 0.4 | 0.4 | 0.3 | 1 | 1.1 | 0.4 | −0.6 | −0.1 |
| fdps | farnesyl diphosphate synthase (farnesyl pyrophosphate synthetase, dimethylallyltranstransferas e, geranyltranstransferase) | 0.1 | 0.4 | 0.1 | −0.1 | −0.4 | 0.4 | 0.02 | −0.1 | −1.2 | 0.03 | 0.4 | 0.3 | 0.5 | 1 | 0.6 | 0.04 | 0.3 | 0.1 |
| Immune System | |||||||||||||||||||
| irg1l | immunoresponsive gene 1, like | 1.5 | 0.2 | 0.04 | −0.5 | −0.2 | −0.2 | 0.2 | −0.7 | 1.8 | 0.8 | 0.4 | 0.6 | −0.3 | 0.8 | 0.2 | −0.3 | 0.6 | 1.1 |
| hsp90b1 | heat shock protein 90, beta (grp94), member 1 | 1 | 0.2 | 0.3 | −0.3 | −0.1 | −0.2 | 0.4 | −0.9 | 0.2 | 0.1 | −0.1 | 0.4 | −0.4 | −0.4 | 0.2 | −0.3 | −0.9 | −0.1 |
| ctsl.1 | cathepsin L.1 | −0.4 | 0.2 | −0.4 | 0.7 | 0.3 | −0.1 | −0.1 | −0.2 | 0.7 | 0.6 | 0.5 | −1.3 | 0.8 | 0.5 | −0.5 | 1 | −0.1 | −0.4 |
| Extracellular Processes | |||||||||||||||||||
| si:dkey-14d8.6 | si:dkey-14d8.6 | −1.1 | −0.4 | 0.01 | 1 | 0.4 | 0 | −0.2 | −0.2 | 0.5 | 0.2 | −0.2 | −0.3 | 0.9 | 0.4 | −0.5 | 0.5 | −0.2 | −0.4 |
| pdk2b | pyruvate dehydrogenase kinase 2b | −1.1 | −0.5 | 0.004 | −0 | 0.2 | 0.9 | −0.1 | 0.3 | 1.2 | 0.5 | −0.2 | −0.5 | 0.3 | 0.4 | 0.3 | 0.8 | 0.5 | −0.6 |
| cpa1 | carboxypeptidase A1 (pancreatic) | −1.3 | −0.9 | −0.5 | 0.1 | 0.02 | 0.3 | −0.1 | −0.6 | −0.2 | −0.1 | −0.3 | −0.6 | 0.3 | 0.4 | 0.3 | 0.9 | −0.8 | −0.4 |
| Intracellular Processes | |||||||||||||||||||
| si:ch211- 153b23.4 | si:ch211-153b23.4 | 1.2 | −0.2 | −0.3 | −0.9 | −0.5 | −0.5 | 0.2 | −0.4 | 1.4 | 0.8 | 0.1 | 0.8 | −0.5 | 1.2 | 0.5 | −0.3 | 0.05 | 0.5 |
| mt-tRNA | tRNA on mitochondrial genome | −0.002 | −0.04 | −0.05 | −0.9 | −0.4 | −0.6 | 0.1 | 0.4 | 0.5 | −0.3 | −1.1 | −0.4 | −0.4 | 0.2 | 0.1 | −1 | −1.9 | −0.4 |
| prelid3b | PRELI domain containing 3 | 0.1 | 0.4 | 0.3 | 1 | 0.4 | 0.5 | −0.4 | 0.1 | −0.5 | 0.4 | 0.8 | 0.1 | 1.3 | 0.3 | −0.1 | 0.2 | −0.6 | 0.2 |
| hnrnpa0l | heterogeneous nuclear ribonucleoprotein A0, like | 0.4 | 0.2 | −0.03 | −0.8 | −0.5 | −0.3 | 0.001 | −1 | −0.8 | 0.2 | 0.5 | 1 | −0.8 | 0.5 | 0.1 | −0.1 | 0.1 | −0.02 |
| calcoco1b | calcium binding and coiled- coil domain 1b | −1 | −0.7 | −0.3 | 0.1 | 0.1 | 0.4 | 0.1 | 0.7 | 2.2 | 0.4 | −0.2 | −1.3 | 0.7 | 0.7 | 0.2 | 0.9 | 1.1 | −0.1 |
| mknk2b | MAPK interacting serine/threonine kinase 2b | −0.9 | −0.5 | −0.2 | 0 | 0.1 | 0.2 | 0.03 | −0.1 | 1.1 | −0 | −0.2 | −0.4 | 0.3 | 0.1 | −0 | 0.6 | −0.1 | 0.1 |
| trim63a | tripartite motif containing 63a | −0.8 | −0.6 | −0.2 | −0.3 | −0.1 | 0.1 | 0.2 | 0.3 | 0.9 | 0.04 | −0.3 | −0.6 | 0.1 | 0.2 | −0.2 | 0.5 | −1.6 | −0.2 |
| si:ch211- 207n23.2 | si:ch211-207n23.2 | 0.9 | 0.3 | 0.03 | −0.5 | −0.4 | −0.5 | 0.3 | −0.3 | 1.5 | 0.8 | 0.5 | 0.2 | −0.3 | 0.5 | 0.1 | 0.2 | 0.3 | 0.4 |
| si:ch211- 153b23.3 | si:ch211-153b23.3 | 1.2 | −0.02 | −0.1 | −0.1 | −0 | −0 | 0.2 | 0.3 | 1.9 | 0.3 | 0.3 | 0.5 | −0 | 0.7 | 0.3 | −0.2 | 0.8 | 1.3 |
| sult5a1 | sulfotransferase family 5A, member 1 | 1.7 | 0.7 | 0.1 | −0.1 | −0 | −0.1 | 0.1 | −0.2 | 2.2 | 0.4 | 0.3 | 0.2 | −0.1 | 1 | 0.1 | −0.3 | 0.9 | 0.7 |
| mt-atp8 | ATP synthase 8, mitochondrial | −0.5 | −0.4 | −0.7 | −0.4 | 0.2 | 0 | −0.2 | 0.8 | 0.1 | −0.7 | −1 | 0.2 | −0.3 | −0.4 | 0 | −0.5 | 0.2 | −0.1 |
| dgcr8 | DGCR8 microprocessor complex subunit | 0.1 | 0.4 | 0.02 | 0.2 | 0.2 | 0.3 | −0.2 | 0.3 | −0.8 | 0.3 | 0.8 | 0.1 | 0.1 | −0.02 | 0.1 | 0.1 | 0.6 | 0.1 |
| bub3 | BUB3 mitotic checkpoint protein | 0.2 | 0.3 | −0.04 | −0.2 | −0.1 | −0.1 | −0.1 | −1.3 | −1.1 | 0.3 | 0.8 | 0.7 | −0.1 | −0.1 | −0.1 | −0.3 | −1 | 0.5 |
| xpot | exportin, tRNA (nuclear export receptor for tRNAs) | −0.3 | 0.2 | −0.3 | 0.1 | −0 | −0.1 | −0.4 | −0.8 | −1.5 | 0.3 | 0.8 | 0.4 | −0 | −0.1 | −0.2 | 0 | 0.4 | 0.9 |
| si:ch211- 250g4.3 | si:ch211-250g4.3 | 0.1 | 0.3 | 0.3 | 0.7 | 0.4 | 0.7 | 0.1 | 0.8 | 1 | 0.3 | 0.8 | −0.05 | 0.8 | 0.5 | 0.3 | 0.8 | −0.2 | −0.02 |
| cebpd | CCAAT enhancer binding protein delta | −0.1 | 0.03 | 0.04 | −0.4 | 0.1 | 0.6 | 0.2 | −0.2 | 1.2 | 0.1 | −0.5 | −0.8 | 0 | 0.4 | 0.4 | 0.9 | 0.01 | −0.2 |
| hnrnpa0a | heterogeneous nuclear ribonucleoprotein A0a | 0.5 | 0.2 | −0.1 | −0.5 | −0.4 | −0.4 | −0.1 | −0.5 | −0.8 | 0.2 | 0.4 | 0.8 | −0.8 | 0 | 0.02 | −0.02 | −0.4 | −0.3 |
| fkbp5 | FKBP prolyl isomerase 5 | −0.4 | 0.1 | 0.1 | 0.4 | 0.7 | 1 | 0.2 | −0.1 | 0.2 | 0.01 | −0.5 | 0.5 | 0.5 | −0.4 | ND | 0.8 | −0.02 | −0.03 |
| ddx39ab | DEAD (Asp-Glu-Ala-Asp) box polypeptide 39Ab | 0.3 | −0.1 | −0.1 | −1 | −0.6 | −0.4 | 0.1 | −0.6 | −0.9 | −0.1 | −0.2 | 0.3 | −1.2 | 0.3 | 0.3 | −0.6 | −0.9 | −0.2 |
| tyrp1b | tyrosinase-related protein 1b | 0.4 | −0.02 | 0.02 | −1 | −0.8 | −0.4 | 0.2 | −0.2 | −1.5 | −0.3 | −0.4 | −0.1 | −0.7 | 0.4 | 0.2 | −1 | −0.4 | −0.2 |
| hnrnpa0b | heterogeneous nuclear ribonucleoprotein A0b | 0.5 | 0.2 | 0.3 | −0.8 | −0.5 | 0.2 | 0.2 | −0.9 | −1.3 | −0.2 | −0.01 | 0.2 | −0.6 | 0.3 | 0.4 | 0.1 | 0.6 | −0.1 |
| phtf2 | putative homeodomain transcription factor 2 | −0.4 | 0.2 | 0.2 | 0.5 | 0.2 | 0.2 | −0.2 | 0.3 | −0 | 0.3 | 0.3 | −0.1 | 0.9 | 0.2 | 0 | −0.2 | −2.4 | 0.2 |
| zmp:0000001081 | zmp:0000001081 | 0.2 | 0.1 | 0.3 | 0.7 | 0.3 | 0.6 | 0.2 | 0.2 | 1.4 | 0.2 | 0.7 | −0.3 | 0.9 | 0.5 | 0.2 | 0.8 | 0.2 | 0.2 |
| si:ch211- 113a14.18 | si:ch211-113a14.18 | −0.003 | −0.1 | 0.02 | 0.1 | 0.3 | 0.1 | 0.1 | 0.7 | 0.1 | −0.1 | −0.2 | −0.3 | −0.1 | −1.4 | −0.5 | 0.2 | 2.4 | −0.1 |
| zgc:113263 | zgc:113263 | −0.1 | −0.2 | 0.1 | 0.2 | 0.01 | 0 | −0.1 | 0.2 | 0.4 | 0.3 | 0.2 | 0.2 | 0.01 | −1.2 | −0.2 | 0.2 | 1.4 | 0.1 |
| si:ch211- 132b12.7 | si:ch211-132b12.7 | 0.1 | 0.4 | −0.1 | 0.2 | 0 | 0.7 | −0.3 | −0.2 | −2 | −0.2 | 0.5 | −0.1 | 0.9 | 1 | 0.5 | 0.1 | −0.03 | −0.1 |
| tm4sf21b | transmembrane 4 L six family member 21b | −0.02 | −0.4 | −0.2 | −0.1 | −0 | 0.2 | 0.3 | −1 | −0.4 | −0.5 | −0.5 | 0.1 | 0.1 | 1.3 | 0.6 | −0.3 | 0.2 | −0.6 |
| Xenobiotic Signaling | |||||||||||||||||||
| cyp3a65 | cytochrome P450, family 3,subfamily A, polypeptide 65 | −1 | −0.6 | −0.1 | 0.5 | 1 | 0.7 | 0.1 | 0.6 | 2 | 0 | −0.5 | −0.8 | 0.5 | 0.1 | 0.1 | −0.1 | −0.6 | −0.4 |
| ucp1 | uncoupling protein 1 | −0.9 | −0.4 | 0.1 | 0.5 | 0.1 | 0.4 | −0.1 | 0.3 | 1.4 | 0.1 | −0.2 | −0.5 | 0.7 | 0.9 | 0.1 | 0.7 | −0.1 | −0.5 |
| cyp2k18 | cytochrome P450, family 2, subfamily K, polypeptide18 | 0.3 | −0.03 | 0.2 | 0.2 | 2 | 2.4 | −0.1 | −0.1 | 1.9 | 0.2 | −0.2 | 0.6 | 0.1 | −0.2 | 0.3 | −0.2 | 2.4 | −0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phillips, J.; Haimbaugh, A.S.; Akemann, C.; Shields, J.N.; Wu, C.-C.; Meyer, D.N.; Baker, B.B.; Siddiqua, Z.; Pitts, D.K.; Baker, T.R. Developmental Phenotypic and Transcriptomic Effects of Exposure to Nanomolar Levels of 4-Nonylphenol, Triclosan, and Triclocarban in Zebrafish (Danio rerio). Toxics 2022, 10, 53. https://doi.org/10.3390/toxics10020053
Phillips J, Haimbaugh AS, Akemann C, Shields JN, Wu C-C, Meyer DN, Baker BB, Siddiqua Z, Pitts DK, Baker TR. Developmental Phenotypic and Transcriptomic Effects of Exposure to Nanomolar Levels of 4-Nonylphenol, Triclosan, and Triclocarban in Zebrafish (Danio rerio). Toxics. 2022; 10(2):53. https://doi.org/10.3390/toxics10020053
Chicago/Turabian StylePhillips, Jessica, Alex S. Haimbaugh, Camille Akemann, Jeremiah N. Shields, Chia-Chen Wu, Danielle N. Meyer, Bridget B. Baker, Zoha Siddiqua, David K. Pitts, and Tracie R. Baker. 2022. "Developmental Phenotypic and Transcriptomic Effects of Exposure to Nanomolar Levels of 4-Nonylphenol, Triclosan, and Triclocarban in Zebrafish (Danio rerio)" Toxics 10, no. 2: 53. https://doi.org/10.3390/toxics10020053
APA StylePhillips, J., Haimbaugh, A. S., Akemann, C., Shields, J. N., Wu, C.-C., Meyer, D. N., Baker, B. B., Siddiqua, Z., Pitts, D. K., & Baker, T. R. (2022). Developmental Phenotypic and Transcriptomic Effects of Exposure to Nanomolar Levels of 4-Nonylphenol, Triclosan, and Triclocarban in Zebrafish (Danio rerio). Toxics, 10(2), 53. https://doi.org/10.3390/toxics10020053