
Developmental Neurotoxicity and Behavioral Screening in Larval Zebrafish with a Comparison to Other Published Results

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. DMSO Evaluation
2.3. Experimental Animals
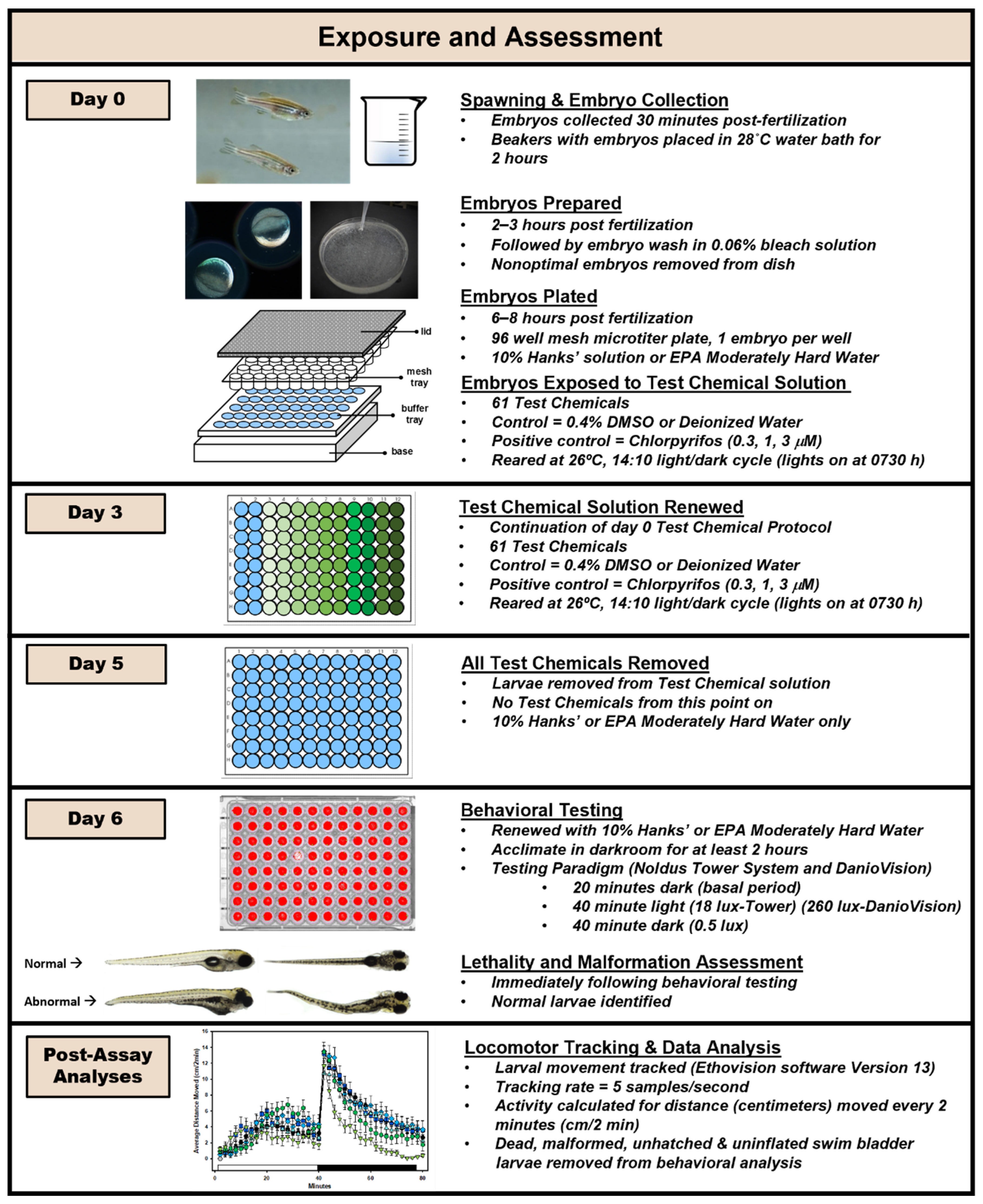
2.4. Experimental Procedure
2.5. General Embryo Rearing
2.6. Chemical Exposure
2.7. Behavioral Testing Systems
2.8. Behavioral Testing
2.9. Lethality and Malformation Assessment and Inclusion Criteria
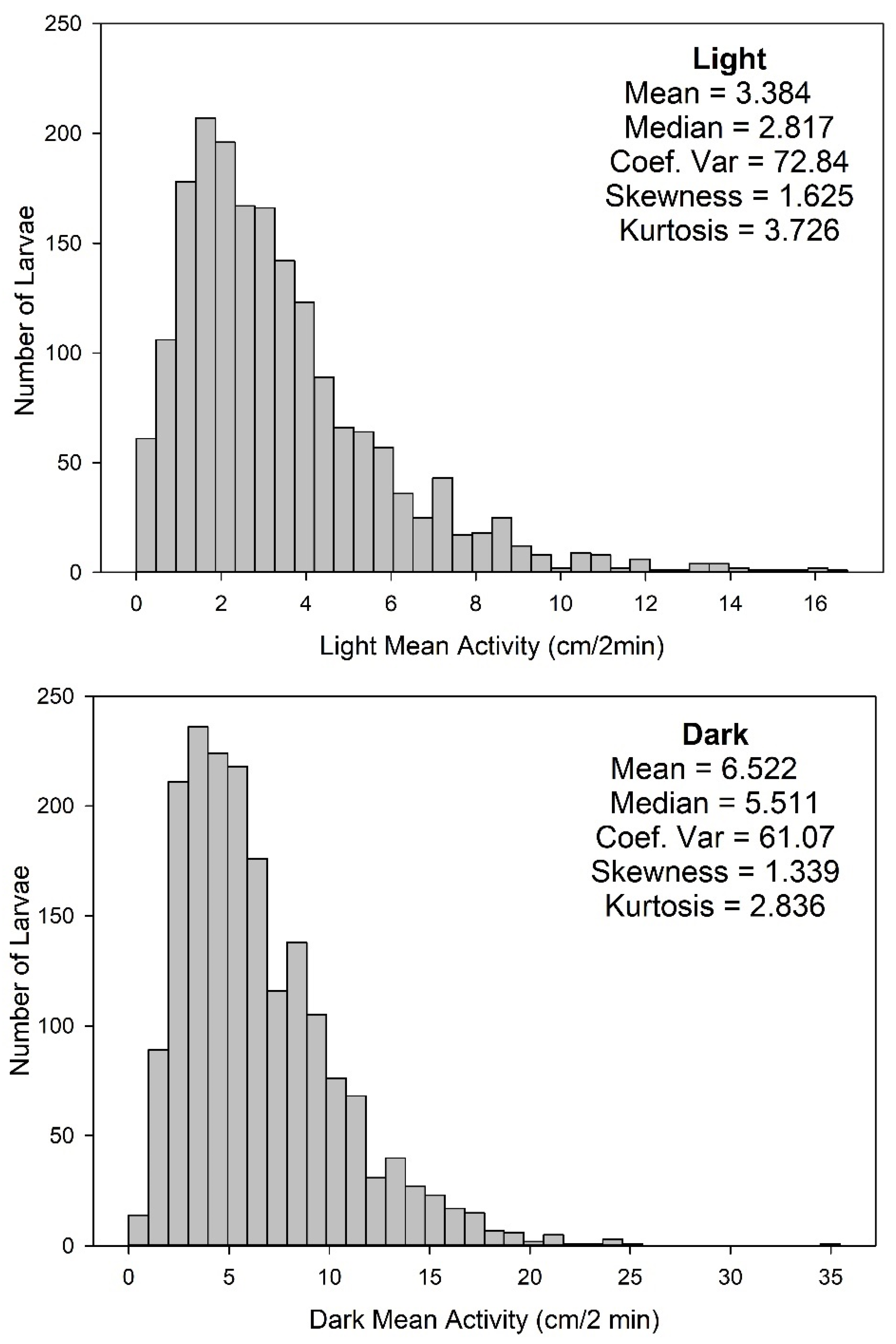
2.10. Analysis of Fish Movement
2.11. Data Analysis and Statistics
2.12. Comparison of Results with Previously Published Data
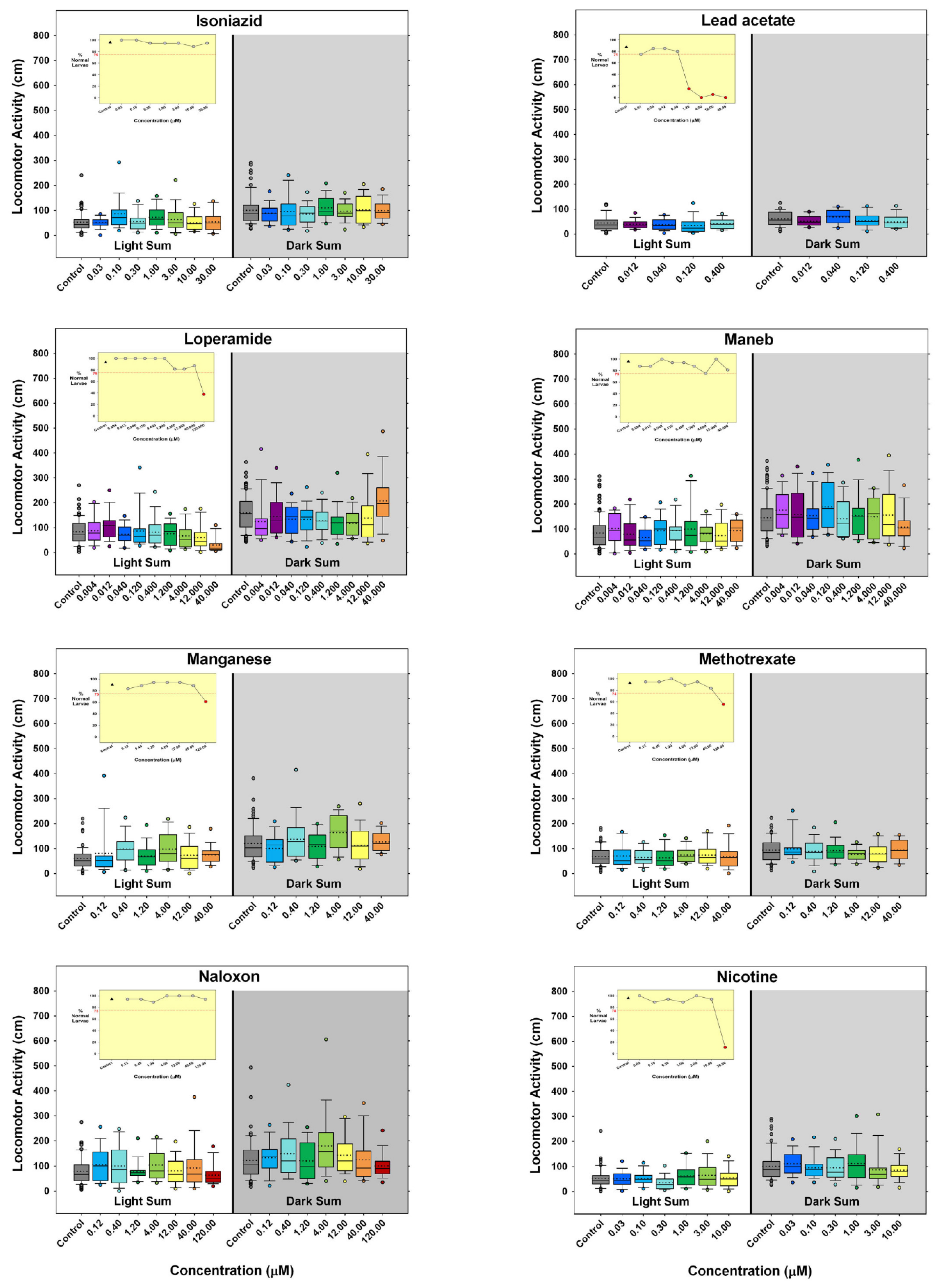
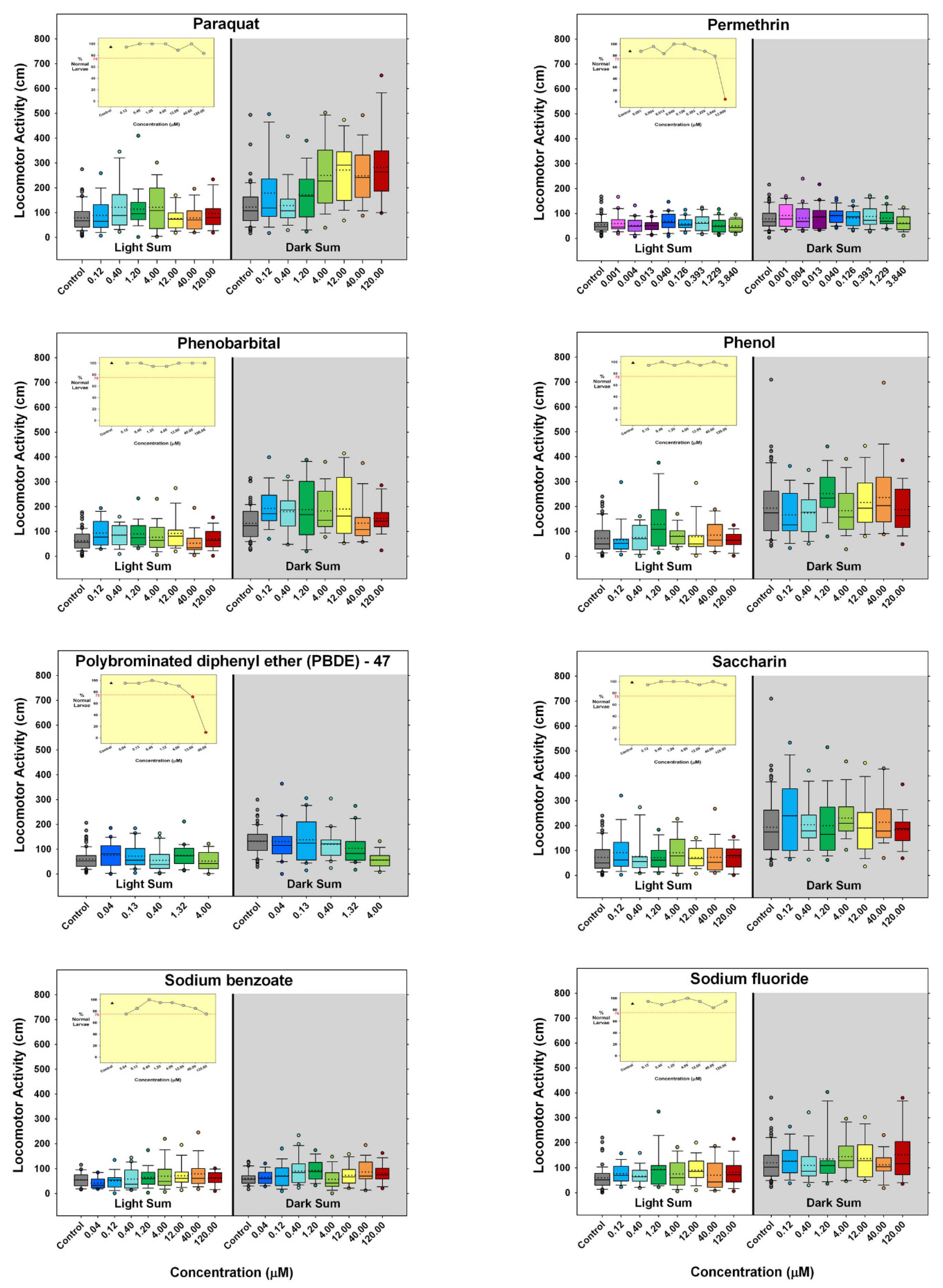
3. Results
4. Discussion
- (1)
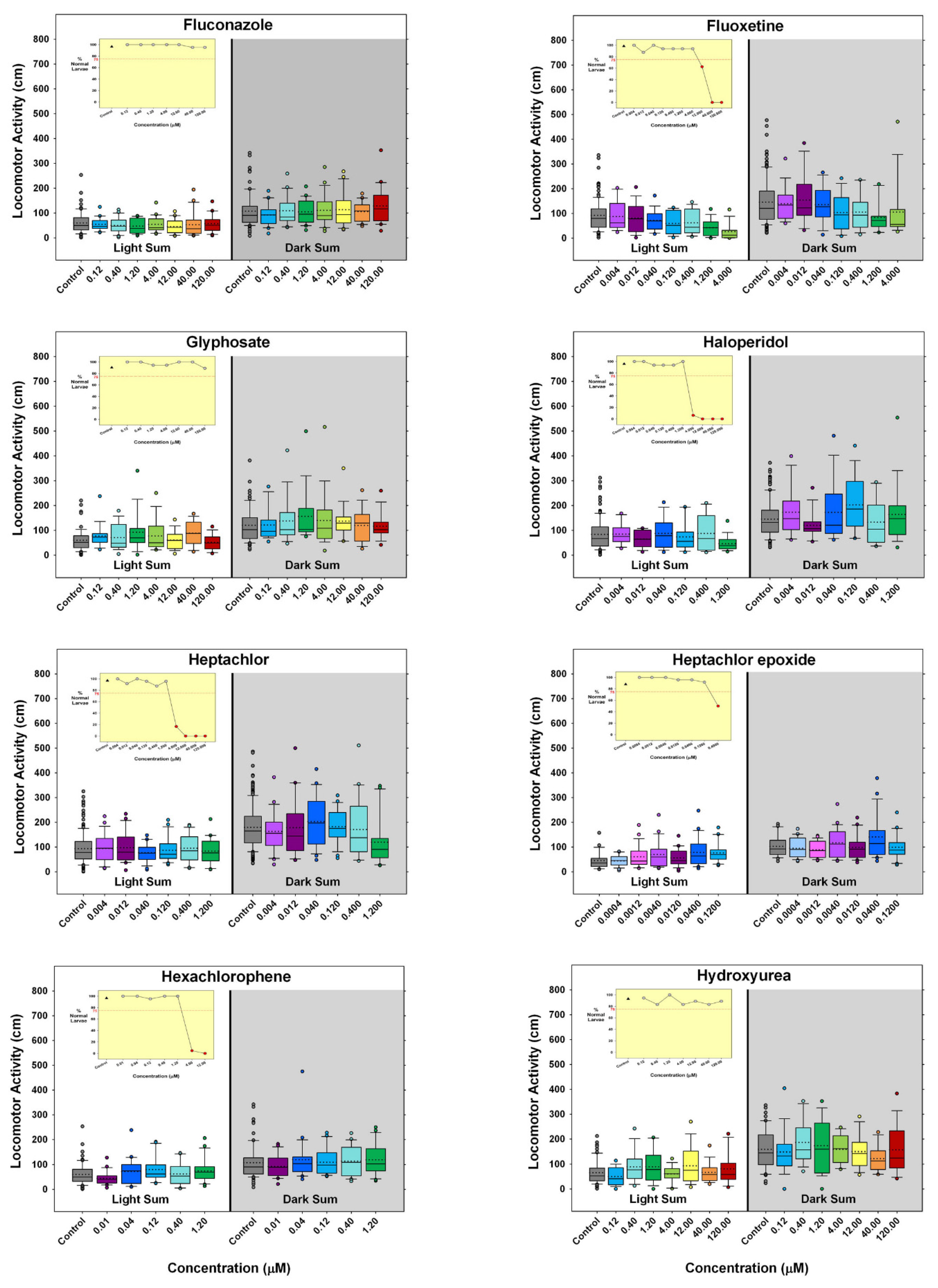
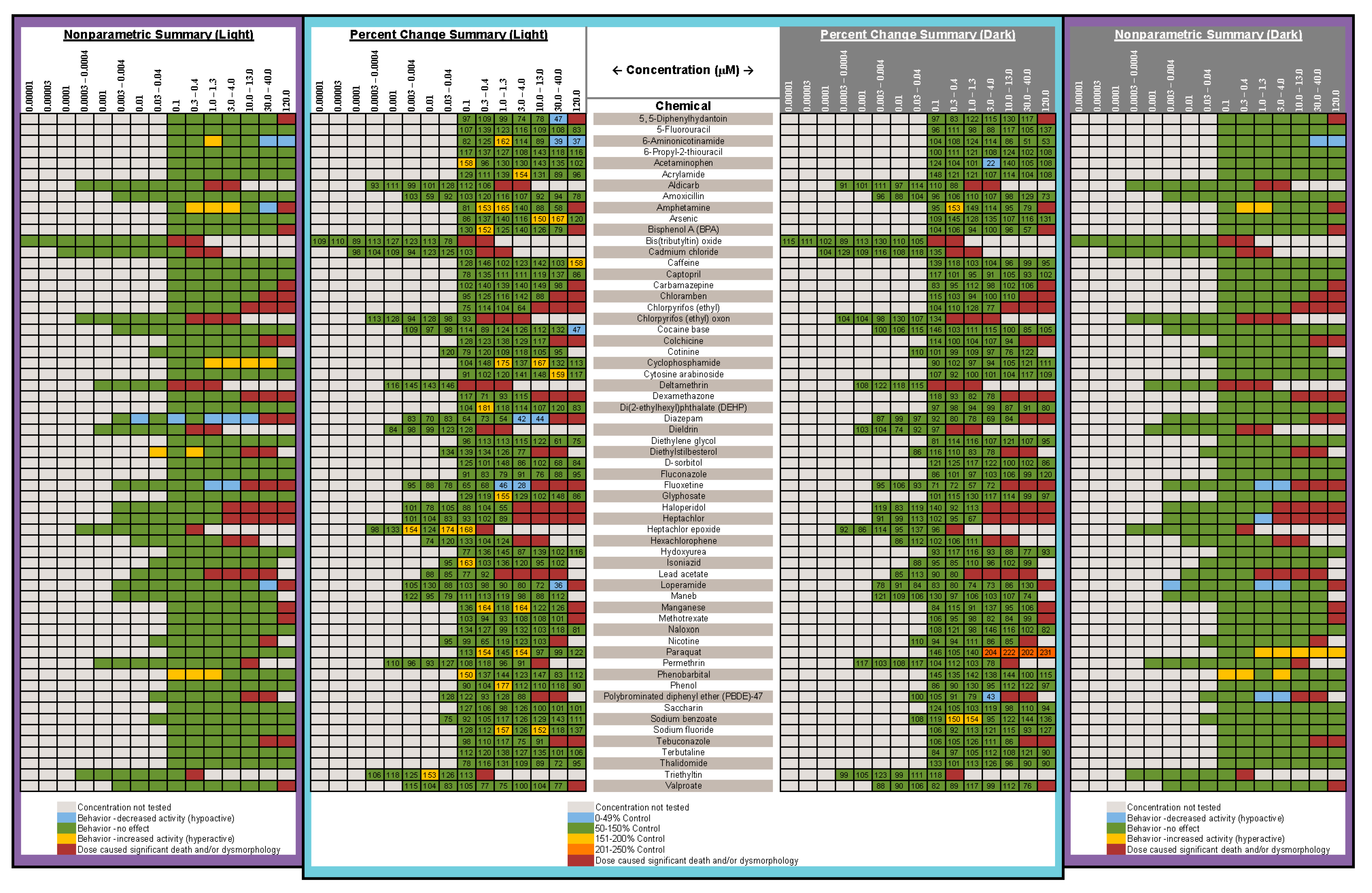
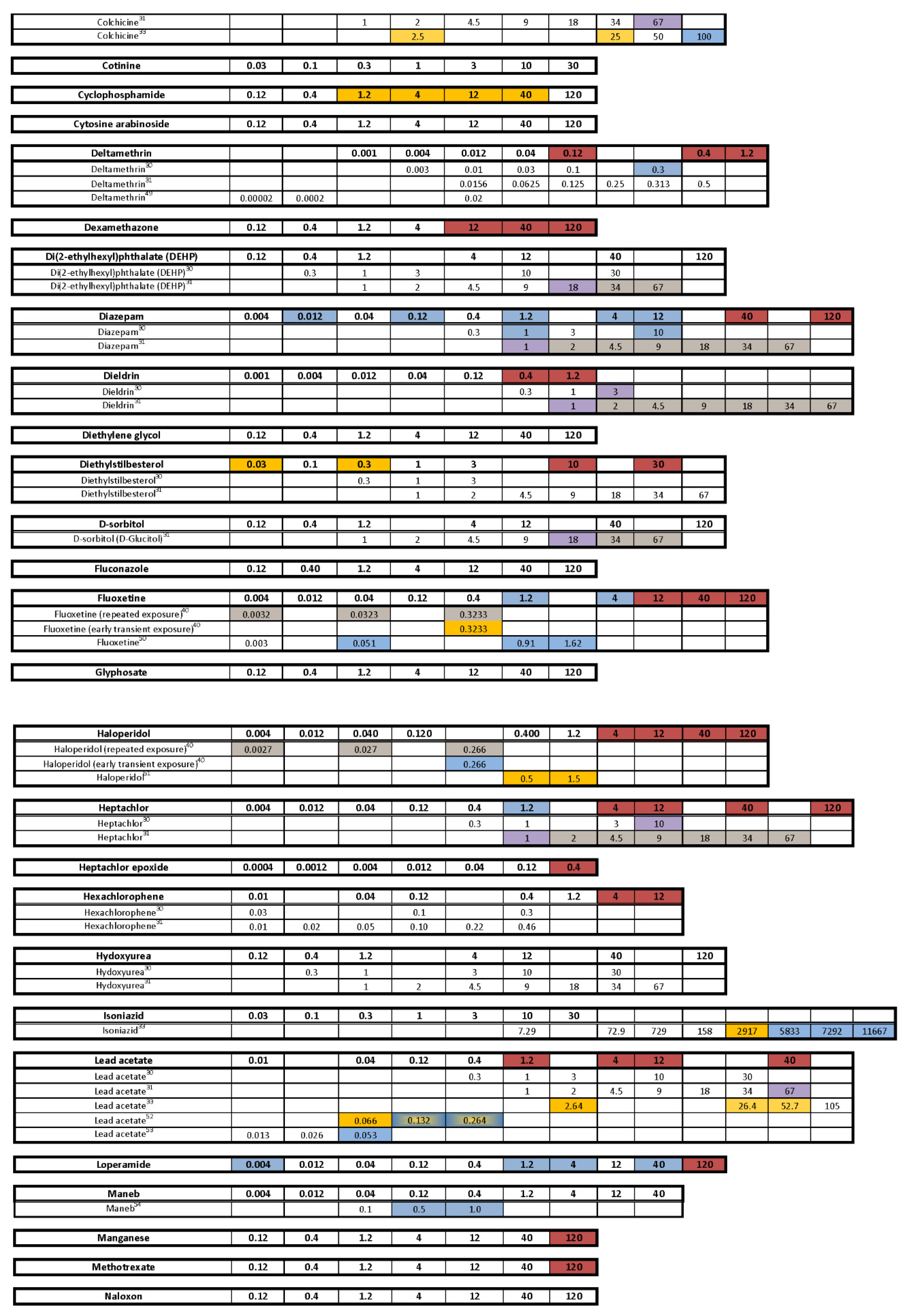
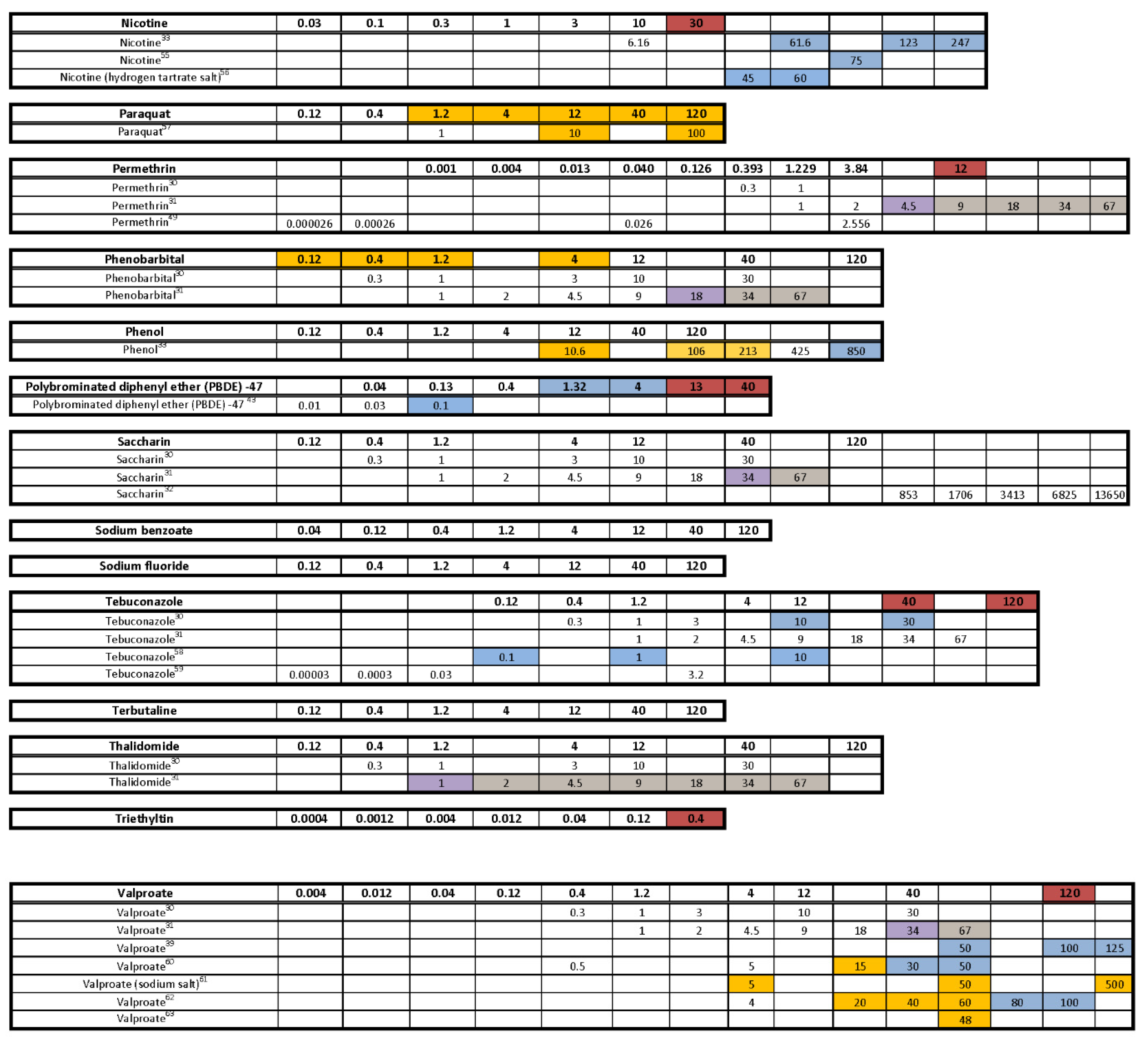
- Are there chemicals among multiple publications that consistently cause or do not cause behavioral effects? This would allow us to identify possible positive and negative controls. There were five chemicals that appear to be candidates for positive controls: diazepam, fluoxetine, paraquat, PBDE-47, and chlorpyrifos. One publication reported decreased activity for diazepam in a similar concentration range as the present study, and the other paper reported behavioral changes, but whether it was an increase or decrease in activity was unclear as only a lowest effective dose was reported. As diazepam is known to be pharmacologically active at the gamma-aminobutyric acid receptor (reviewed in [91]), perhaps diazepam could be regarded as a positive control for GABAergic chemicals. For fluoxetine, one publication, as well as our own, reported decreased activity in larvae treated with fluoxetine during development, while another publication reported increased activity in animals treated with fluoxetine transiently during an early developmental window. Some of the effective concentration ranges aligned. As fluoxetine is a serotonin reuptake inhibitor, this chemical may serve as a positive control for the serotonergic disrupting class of chemicals. Although only one other publication tested paraquat in a developmental neurotoxicity test using zebrafish, the results were very similar to the present study, with both reporting markedly increased activity in the same dosage range. As paraquat has been reported to disrupt the development of the dopaminergic nervous system (reviewed in [92]), this chemical may serve as a positive control for the dopaminergic disrupting class of chemicals. The data for PBDE-47 as a possible positive control are a bit weaker mainly because only one other publication investigated the behavioral effects of developmental exposure to PBDE-47, and the effective concentration range did not overlap with our own data; however, both noted decreased activity. The fifth chemical that might serve as a positive control among testing publications is chlorpyrifos. There are multiple reports of developmental chlorpyrifos exposure producing behavioral alterations in larval zebrafish assays, but the range of effective concentrations spans four orders of magnitude. Because chlorpyrifos is an anticholinesterase, this chemical could serve as a positive control for the disruption of the cholinergic nervous system during development. In contrast, there are four chemicals that are candidates for negative controls, although the number of observations is smaller: aldicarb, amoxicillin, hexachlorophene and hydroxyurea. In all cases, there are two publications as well as the present study showing that developmental exposure to these chemicals in approximately the same concentration range did not produce behavioral alterations in the larval locomotor assay.
- (2)
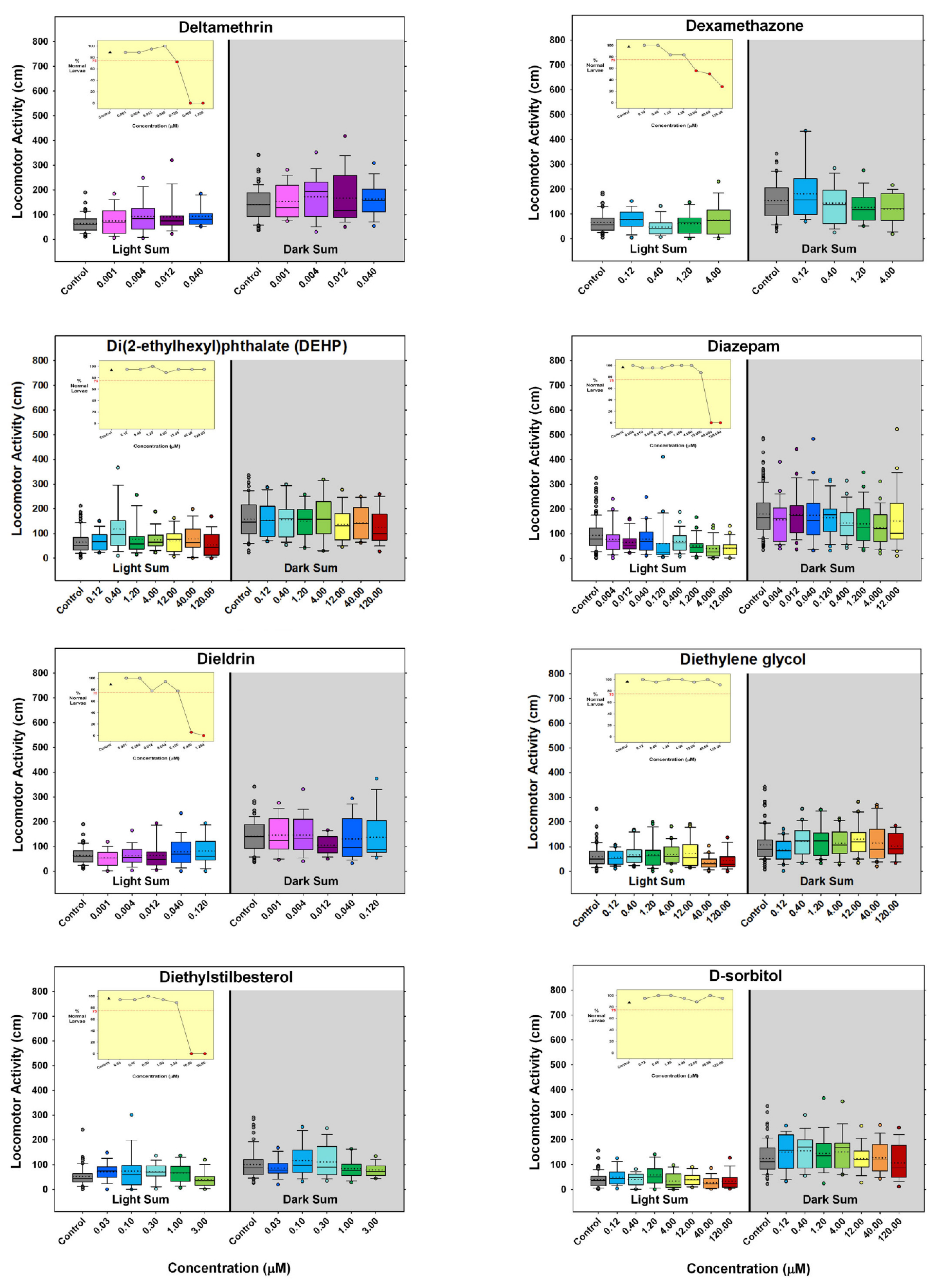
- Are there chemicals that other publications have shown to produce behavioral changes after developmental exposure, but at concentrations that exceeded our concentration range or at concentrations that we deemed developmentally toxic? Eight chemicals (aldicarb, cadmium chloride, caffeine, carbamazepine, deltamethrin, dieldrin, isoniazid, nicotine) would fall into that category. In fact, caffeine and isoniazid did not appear to produce behavioral effects unless tested in the millimolar range.
- (3)
- Are there unique chemicals that only our laboratory has tested that produced changes in larval locomotor activity after developmental exposure? There were four chemicals that were tested in this publication that produced changes in locomotor activity after developmental exposure that other publications appear not to have tested: developmental exposure to 6-aminonicotinamide or loperamide produced decreased activity in the larvae, and developmental exposure to amphetamine produced an inverted “U” biphasic pattern of increased activity at lower concentrations and decreased activity at the higher concentrations. Cyclophosphamide also showed increased activity in the middle concentrations. In our laboratory embryos treated with diethylstilbesterol during development showed increased activity at concentrations below those that tested negative in other publications.
- (4)
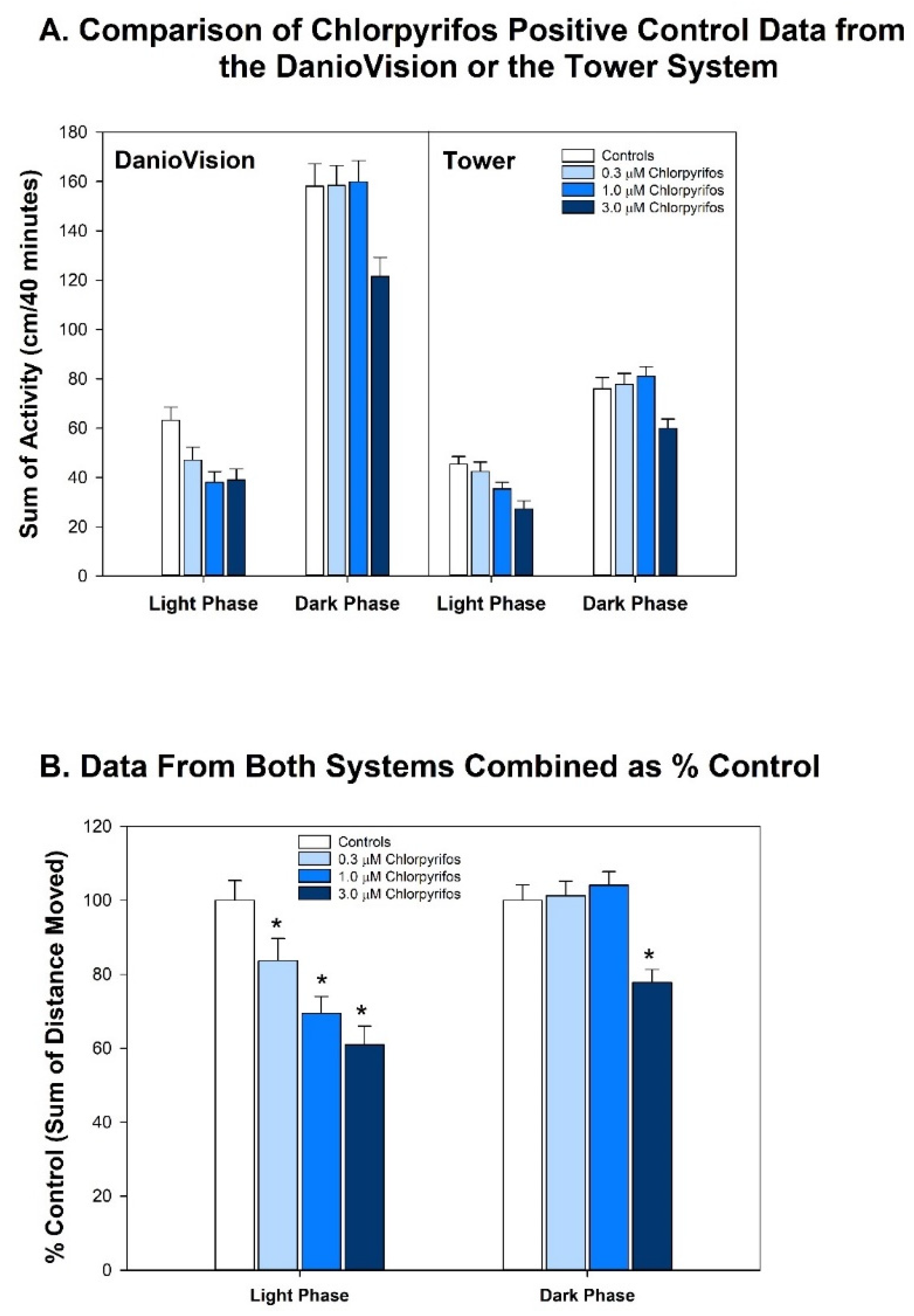
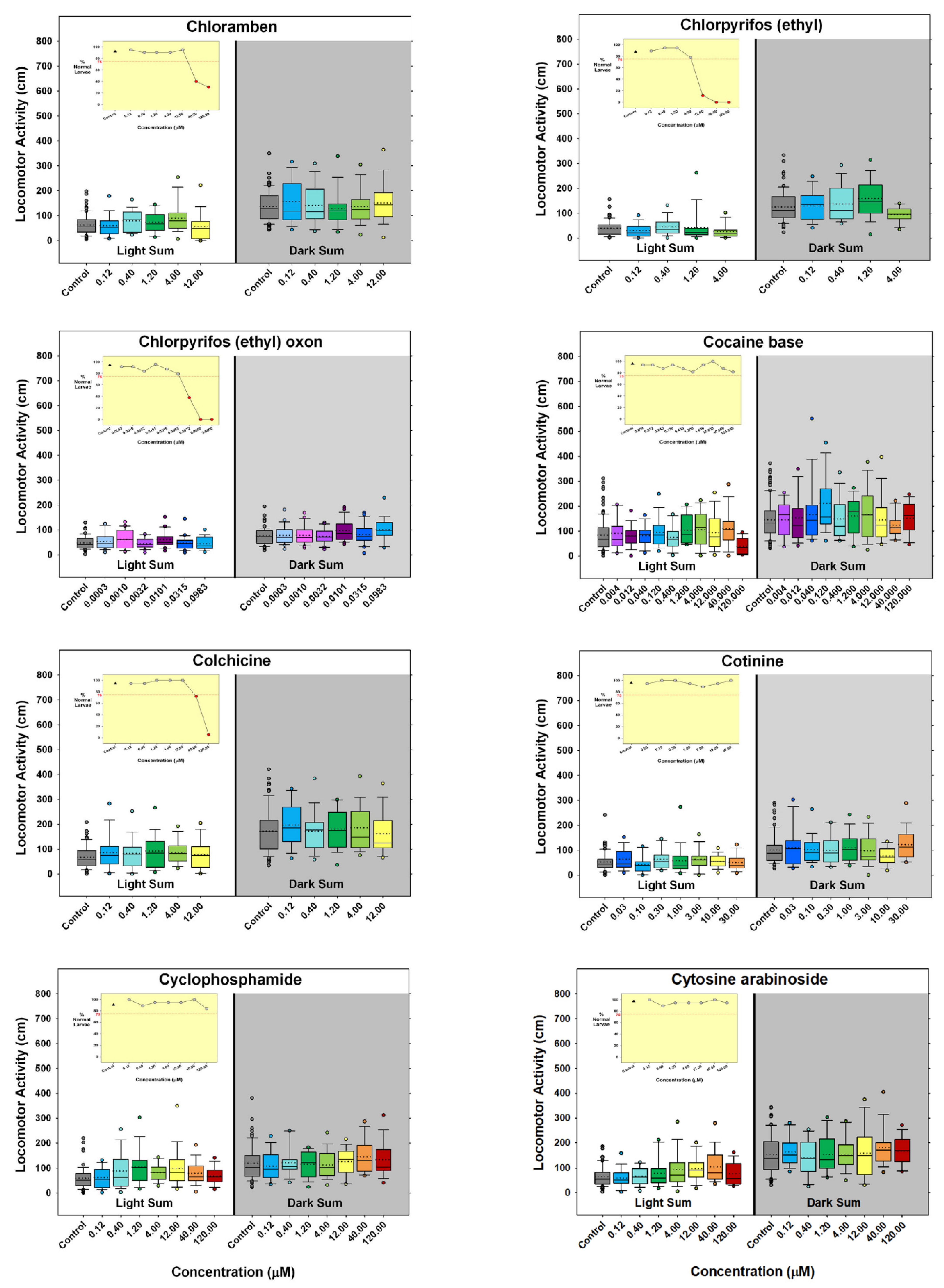
- Are there chemicals that have shown behavioral effects in other studies, but were not positive in our study? There were three chemicals, valproate, chlorpyrifos, and lead (Pb) that fall into this category. Positive results were expected for chlorpyrifos and valproate because they have tested positive previous times in our laboratory [42,62,63,64]. Specifically, we have published two papers showing developmental valproate exposure elicits behavioral changes in larval zebrafish [62,63]. It appears that the developmental toxicity profile in the present study is similar to the previously published papers: 120 µM concentration caused malformations and death in a large portion of the larvae, and 40 µM was on the cusp of developmental toxicity. The behavioral toxicity, however, was not apparent in this present study as it had been in the previous studies. The other two publications i.e., [62,63] tested about twice as many animals per concentration, so perhaps this present result is an issue of statistical power. Statistical power may have also played a role in the disparate results for chlorpyrifos in the present study. In this study we tested chlorpyrifos in two different scenarios: one as a positive control throughout the study and the other as one of the chemicals under investigation. The results are summarized for both in Figure 7 with the positive control data listed as “Chlorpyrifos+”, and the data for the test chemical listed as “Chlorpyrifos (ethyl).” As our positive control with many more observations (n = 115–132 per concentration), chlorpyrifos produced positive results in the same pattern that we often see: hypoactivity in both the Light and Dark periods, with the Dark period activity being less sensitive than the Light period activity (details in Figure 2). When testing chlorpyrifos as one of our test chemicals, however, with fewer observations (n = 14–16 per concentration), we hypothesize that there was less statistical power to detect the change. These negative results for chlorpyrifos or valproate indicate that we may need to increase the number of observations at each concentration in future developmental neurotoxicity screens. A power analysis was done when setting up our experimental design, but because the behavioral data are skewed, and require nonparametric analysis, it is difficult to perform an accurate power analysis for non-normally distributed endpoints. Lead (Pb) was another chemical where we expected a positive result given that four out of the five previous publications reported behavioral changes in larval zebrafish (Figure 7). Our results showed developmental toxicity ≥ 1.2 µM. Many of the larvae in the 1.2 and 4.0 µM concentrations showed a preponderance of uninflated swim bladders in the absence of other malformations, and therefore were not included in the behavioral assessment. If these animals had been tested in the behavioral protocol, there would have been markedly decreased activity in the Light period. One possibility to consider would be that swim bladder inflation may be a neurotoxic endpoint. Inflation of the swim bladder not only requires innervation [93,94], but it also requires a behavioral repertoire where the larva seeks out the air/water interface to take a gulp of air [95]. So perhaps swim bladder non-inflation belongs intercalated between a morphological and behavioral endpoint, and if an animal presents with an uninflated swim bladder, this could be logged as a potentially neurotoxic endpoint without behavioral confirmation.
- (5)
- Were there chemicals that showed considerable variation in the published results? Four publications, including ours, tested 6-propyl-2-thiouracil with overlapping concentrations spanning about three orders of magnitude; only one publication out of the four reported changes in behavior. Six publications including our own tested acetaminophen, again with many testing in the same concentration ranges, and yet only three of the publications reporting changes in behavior. There was some overlap in the positive concentrations in two of the publications, but the third publication only found behavioral changes at millimolar concentrations. Only one out of four publications found that developmental carbamazepine produced behavioral alterations in larval zebrafish tested in the Light/Dark transition assay. For deltamethrin, only one out of four publications found behavioral changes, whereas our laboratory reported developmental toxicity in the concentration range where the behavioral changes were reported. Three out of four publications did not find behavioral alterations after developmental saccharin exposure. Interestingly, saccharin is one of the few chemicals in this testing library that was classified as a ”favorable” negative control chemical for developmental neurotoxicity screens [96], meaning that an expert panel’s assessment of the chemical showed very little to no evidence that the chemical produces developmental neurotoxicity. Five publications studied the effects of tebuconazole on behavioral profiles in larval zebrafish with two publications reporting a positive result, and the other three publications testing in that same concentration range reported negative results. There were also contrasting results with thalidomide, where one out of three publications reported behavioral changes, but the other two publications reported a negative result in the same concentration range. These types of discrepancies indicate that the zebrafish larval Light/Dark locomotor assay will require more protocol and analysis standardization among laboratories.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghassabian, A.; Vandenberg, L.; Kannan, K.; Trasande, L. Endocrine-Disrupting Chemicals and Child Health. Annu. Rev. Pharmacol. Toxicol. 2022, 62, 573–594. [Google Scholar] [CrossRef]
- Zablotsky, B.; Black, L.I.; Maenner, M.J.; Schieve, L.A.; Danielson, M.L.; Bitsko, R.H.; Blumberg, S.J.; Kogan, M.D.; Boyle, C.A. Prevalence and Trends of Developmental Disabilities among Children in the United States: 2009–2017. Pediatrics 2019, 144, e20190811. [Google Scholar] [CrossRef]
- Bennett, D.; Bellinger, D.C.; Birnbaum, L.S.; Bradman, A.; Chen, A.; Cory-Slechta, D.A.; Engel, S.M.; Fallin, M.D.; Halladay, A.; Hauser, R.; et al. Project TENDR: Targeting Environmental Neuro-Developmental Risks The TENDR Consensus Statement. Environ. Health Perspect. 2016, 124, A118–A122. [Google Scholar] [CrossRef]
- Balaguer-Trias, J.; Deepika, D.; Schuhmacher, M.; Kumar, V. Impact of Contaminants on Microbiota: Linking the Gut-Brain Axis with Neurotoxicity. Int. J. Environ. Res. Public Health 2022, 19, 1368. [Google Scholar] [CrossRef]
- Dórea, J.G. Exposure to environmental neurotoxic substances and neurodevelopment in children from Latin America and the Caribbean. Environ. Res. 2021, 192, 110199. [Google Scholar] [CrossRef]
- Preston, M.A.; Finseth, L.T.; Bourne, J.N.; Macklin, W.B. A novel myelin protein zero transgenic zebrafish designed for rapid readout of in vivo myelination. Glia 2019, 67, 650–667. [Google Scholar] [CrossRef]
- Baier, H.; Wullimann, M.F. Anatomy and function of retinorecipient arborization fields in zebrafish. J. Comp. Neurol. 2021, 529, 3454–3476. [Google Scholar] [CrossRef]
- Thyme, S.B.; Pieper, L.M.; Li, E.H.; Pandey, S.; Wang, Y.; Morris, N.S.; Sha, C.; Choi, J.W.; Herrera, K.J.; Soucy, E.R.; et al. Phenotypic Landscape of Schizophrenia-Associated Genes Defines Candidates and Their Shared Functions. Cell 2019, 177, 478–491.e420. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.D.; Bonath, F.; Kumar, S.; Ross, C.A.; Cunliffe, V.T. Disrupted-in-schizophrenia 1 and neuregulin 1 are required for the specification of oligodendrocytes and neurones in the zebrafish brain. Hum. Mol. Genet. 2009, 18, 391–404. [Google Scholar] [CrossRef] [Green Version]
- Pilorge, M.; Fassier, C.; Le Corronc, H.; Potey, A.; Bai, J.; De Gois, S.; Delaby, E.; Assouline, B.; Guinchat, V.; Devillard, F.; et al. Genetic and functional analyses demonstrate a role for abnormal glycinergic signaling in autism. Mol. Psychiatry 2016, 21, 936–945. [Google Scholar] [CrossRef] [Green Version]
- Goldshtein, H.; Muhire, A.; Petel Legare, V.; Pushett, A.; Rotkopf, R.; Shefner, J.M.; Peterson, R.T.; Armstrong, G.A.B.; Russek-Blum, N. Efficacy of Ciprofloxacin/Celecoxib combination in zebrafish models of amyotrophic lateral sclerosis. Ann. Clin. Transl. Neurol. 2020, 7, 1883–1897. [Google Scholar] [CrossRef]
- Tilson, H.A. Neurobehavioral methods used in neurotoxicological research. Toxicol. Lett. 1993, 68, 231–240. [Google Scholar] [CrossRef]
- Bushnell, P.J. Testing for cognitive function in animals in a regulatory context. Neurotoxicol. Teratol. 2015, 52, 68–77. [Google Scholar] [CrossRef]
- Bownik, A.; Wlodkowic, D. Applications of advanced neuro-behavioral analysis strategies in aquatic ecotoxicology. Sci. Total Environ. 2021, 772, 145577. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a comprehensive catalog of zebrafish behavior 1.0 and beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef]
- Fero, K.; Yokogawa, T.; Burgess, H.A. The behavioral repertoire of larval zebrafish. In Zebrafish Models in Neurobehavioral Research; Kalueff, A., Cachat, J., Eds.; Neuromethods; Humana Press: Totowa, NJ, USA, 2011; Volume 52. [Google Scholar]
- Couderq, S.; Leemans, M.; Fini, J.B. Testing for thyroid hormone disruptors, a review of non-mammalian in vivo models. Mol. Cell. Endocrinol. 2020, 508, 110779. [Google Scholar] [CrossRef]
- Porazzi, P.; Calebiro, D.; Benato, F.; Tiso, N.; Persani, L. Thyroid gland development and function in the zebrafish model. Mol. Cell. Endocrinol. 2009, 312, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Walter, K.M.; Miller, G.W.; Chen, X.; Yaghoobi, B.; Puschner, B.; Lein, P.J. Effects of thyroid hormone disruption on the ontogenetic expression of thyroid hormone signaling genes in developing zebrafish (Danio rerio). Gen. Comp. Endocrinol. 2019, 272, 20–32. [Google Scholar] [CrossRef]
- Eachus, H.; Choi, M.K.; Ryu, S. The Effects of Early Life Stress on the Brain and Behaviour: Insights From Zebrafish Models. Front. Cell Dev. Biol. 2021, 9, 657591. [Google Scholar] [CrossRef]
- Goldstone, J.V.; McArthur, A.G.; Kubota, A.; Zanette, J.; Parente, T.; Jonsson, M.E.; Nelson, D.R.; Stegeman, J.J. Identification and developmental expression of the full complement of Cytochrome P450 genes in Zebrafish. BMC Genom. 2010, 11, 643. [Google Scholar] [CrossRef] [Green Version]
- Nawaji, T.; Yamashita, N.; Umeda, H.; Zhang, S.; Mizoguchi, N.; Seki, M.; Kitazawa, T.; Teraoka, H. Cytochrome P450 Expression and Chemical Metabolic Activity before Full Liver Development in Zebrafish. Pharmaceuticals 2020, 13, 456. [Google Scholar] [CrossRef]
- de Esch, C.; Slieker, R.; Wolterbeek, A.; Woutersen, R.; de Groot, D. Zebrafish as potential model for developmental neurotoxicity testing: A mini review. Neurotoxicol. Teratol. 2012, 34, 545–553. [Google Scholar] [CrossRef]
- Fitzgerald, J.A.; Konemann, S.; Krumpelmann, L.; Zupanic, A.; Vom Berg, C. Approaches to Test the Neurotoxicity of Environmental Contaminants in the Zebrafish Model: From Behavior to Molecular Mechanisms. Environ. Toxicol. Chem. 2021, 40, 989–1006. [Google Scholar] [CrossRef]
- Nishimura, Y.; Murakami, S.; Ashikawa, Y.; Sasagawa, S.; Umemoto, N.; Shimada, Y.; Tanaka, T. Zebrafish as a systems toxicology model for developmental neurotoxicity testing. Congenit. Anom. (Kyoto) 2015, 55, 1–16. [Google Scholar] [CrossRef]
- Mundy, W.R.; Padilla, S.; Breier, J.M.; Crofton, K.M.; Gilbert, M.E.; Herr, D.W.; Jensen, K.F.; Radio, N.M.; Raffaele, K.C.; Schumacher, K.; et al. Expanding the test set: Chemicals with potential to disrupt mammalian brain development. Neurotoxicol. Teratol. 2015, 52, 25–35. [Google Scholar] [CrossRef]
- Harrill, J.A.; Freudenrich, T.; Wallace, K.; Ball, K.; Shafer, T.J.; Mundy, W.R. Testing for developmental neurotoxicity using a battery of in vitro assays for key cellular events in neurodevelopment. Toxicol. Appl. Pharmacol. 2018, 354, 24–39. [Google Scholar] [CrossRef]
- Shafer, T.J.; Brown, J.P.; Lynch, B.; Davila-Montero, S.; Wallace, K.; Friedman, K.P. Evaluation of Chemical Effects on Network Formation in Cortical Neurons Grown on Microelectrode Arrays. Toxicol. Sci. 2019, 169, 436–455. [Google Scholar] [CrossRef]
- Carstens, K.E.; Carpenter, A.F.; Martin, M.M.; Harrill, J.A.; Shafer, T.J.; Paul Friedman, K. Integrating data from in vitro New Approach Methodologies for Developmental Neurotoxicity. Toxicol. Sci. 2022, 187, 62–79. [Google Scholar] [CrossRef]
- Dach, K.; Yaghoobi, B.; Schmuck, M.R.; Carty, D.R.; Morales, K.M.; Lein, P.J. Teratological and Behavioral Screening of the National Toxicology Program 91-Compound Library in Zebrafish (Danio rerio). Toxicol. Sci. 2019, 167, 77–91. [Google Scholar] [CrossRef]
- Hagstrom, D.; Truong, L.; Zhang, S.; Tanguay, R.; Collins, E.S. Comparative Analysis of Zebrafish and Planarian Model Systems for Developmental Neurotoxicity Screens Using an 87-Compound Library. Toxicol. Sci. 2019, 167, 15–25. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.; Hooyberghs, J.; Blust, R.; Witters, H.E. Assessment of the developmental neurotoxicity of compounds by measuring locomotor activity in zebrafish embryos and larvae. Neurotoxicol. Teratol. 2013, 37, 44–56. [Google Scholar] [CrossRef]
- Ali, S.; Champagne, D.L.; Richardson, M.K. Behavioral profiling of zebrafish embryos exposed to a panel of 60 water-soluble compounds. Behav. Brain Res. 2012, 228, 272–283. [Google Scholar] [CrossRef]
- Nogueira, A.F.; Pinto, G.; Correia, B.; Nunes, B. Embryonic development, locomotor behavior, biochemical, and epigenetic effects of the pharmaceutical drugs paracetamol and ciprofloxacin in larvae and embryos of Danio rerio when exposed to environmental realistic levels of both drugs. Environ. Toxicol. 2019, 34, 1177–1190. [Google Scholar] [CrossRef]
- Xia, L.; Zheng, L.; Zhou, J.L. Effects of ibuprofen, diclofenac and paracetamol on hatch and motor behavior in developing zebrafish (Danio rerio). Chemosphere 2017, 182, 416–425. [Google Scholar] [CrossRef]
- Fraser, T.W.K.; Khezri, A.; Jusdado, J.G.H.; Lewandowska-Sabat, A.M.; Henry, T.; Ropstad, E. Toxicant induced behavioural aberrations in larval zebrafish are dependent on minor methodological alterations. Toxicol. Lett. 2017, 276, 62–68. [Google Scholar] [CrossRef]
- Wang, X.; Dong, Q.; Chen, Y.; Jiang, H.; Xiao, Q.; Wang, Y.; Li, W.; Bai, C.; Huang, C.; Yang, D. Bisphenol A affects axonal growth, musculature and motor behavior in developing zebrafish. Aquat. Toxicol. 2013, 142–143, 104–113. [Google Scholar] [CrossRef]
- Olsvik, P.A.; Whatmore, P.; Penglase, S.J.; Skjaerven, K.H.; Angles d’Auriac, M.; Ellingsen, S. Associations Between Behavioral Effects of Bisphenol A and DNA Methylation in Zebrafish Embryos. Front. Genet. 2019, 10, 184. [Google Scholar] [CrossRef] [Green Version]
- Pruvot, B.; Quiroz, Y.; Voncken, A.; Jeanray, N.; Piot, A.; Martial, J.A.; Muller, M. A panel of biological tests reveals developmental effects of pharmaceutical pollutants on late stage zebrafish embryos. Reprod. Toxicol. 2012, 34, 568–583. [Google Scholar] [CrossRef] [Green Version]
- Huang, I.J.; Sirotkin, H.I.; McElroy, A.E. Varying the exposure period and duration of neuroactive pharmaceuticals and their metabolites modulates effects on the visual motor response in zebrafish (Danio rerio) larvae. Neurotoxicol. Teratol. 2019, 72, 39–48. [Google Scholar] [CrossRef]
- Pohl, J.; Ahrens, L.; Carlsson, G.; Golovko, O.; Norrgren, L.; Weiss, J.; Orn, S. Embryotoxicity of ozonated diclofenac, carbamazepine, and oxazepam in zebrafish (Danio rerio). Chemosphere 2019, 225, 191–199. [Google Scholar] [CrossRef]
- Dishaw, L.V.; Hunter, D.L.; Padnos, B.; Padilla, S.; Stapleton, H.M. Developmental exposure to organophosphate flame retardants elicits overt toxicity and alters behavior in early life stage zebrafish (Danio rerio). Toxicol. Sci. 2014, 142, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazer, L.; Wells, C.N.; Drastal, M.; Odamah, K.A.; Galat, R.E.; Behl, M.; Levin, E.D. Developmental exposure to low concentrations of two brominated flame retardants, BDE-47 and BDE-99, causes life-long behavioral alterations in zebrafish. Neurotoxicology 2018, 66, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhang, L.; Shi, Q.; Guo, Y.; Zhang, W.; Zhou, B. A protective role of autophagy in TDCIPP-induced developmental neurotoxicity in zebrafish larvae. Aquat. Toxicol. 2018, 199, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, H.; Mi, C.; Feng, C.; Zhang, L.; Yang, L.; Zhou, B. The adverse effect of TCIPP and TCEP on neurodevelopment of zebrafish embryos/larvae. Chemosphere 2019, 220, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Oliveri, A.N.; Bailey, J.M.; Levin, E.D. Developmental exposure to organophosphate flame retardants causes behavioral effects in larval and adult zebrafish. Neurotoxicol. Teratol. 2015, 52, 220–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richendrfer, H.; Pelkowski, S.D.; Colwill, R.M.; Creton, R. Developmental sub-chronic exposure to chlorpyrifos reduces anxiety-related behavior in zebrafish larvae. Neurotoxicol. Teratol. 2012, 34, 458–465. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Xu, W.; Peng, T.; Chen, H.; Ren, L.; Tan, H.; Xiao, D.; Qian, H.; Fu, Z. Developmental exposure of zebrafish larvae to organophosphate flame retardants causes neurotoxicity. Neurotoxicol. Teratol. 2016, 55, 16–22. [Google Scholar] [CrossRef]
- Awoyemi, O.M.; Kumar, N.; Schmitt, C.; Subbiah, S.; Crago, J. Behavioral, molecular and physiological responses of embryo-larval zebrafish exposed to types I and II pyrethroids. Chemosphere 2019, 219, 526–537. [Google Scholar] [CrossRef]
- de Farias, N.O.; Oliveira, R.; Sousa-Moura, D.; de Oliveira, R.C.S.; Rodrigues, M.A.C.; Andrade, T.S.; Domingues, I.; Camargo, N.S.; Muehlmann, L.A.; Grisolia, C.K. Exposure to low concentration of fluoxetine affects development, behaviour and acetylcholinesterase activity of zebrafish embryos. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 215, 1–8. [Google Scholar] [CrossRef]
- Oliveri, A.N.; Levin, E.D. Dopamine D1 and D2 receptor antagonism during development alters later behavior in zebrafish. Behav. Brain Res. 2019, 356, 250–256. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Y.; Liu, W.; Bai, C.; Liu, X.; Liu, K.; Li, R.; Zhu, J.H.; Huang, C. Developmental lead acetate exposure induces embryonic toxicity and memory deficit in adult zebrafish. Neurotoxicol. Teratol. 2012, 34, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Wang, Q.; Shi, X.; Guo, Y.; Xu, T.; Zhou, B. Effect of combined exposure to lead and decabromodiphenyl ether on neurodevelopment of zebrafish larvae. Chemosphere 2016, 144, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Souders, C.L., 2nd; Li, P.; Pang, S.; Liang, X.; Qiu, L.; Martyniuk, C.J. Developmental neurotoxicity of maneb: Notochord defects, mitochondrial dysfunction and hypoactivity in zebrafish (Danio rerio) embryos and larvae. Ecotoxicol. Environ. Saf. 2019, 170, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Holden, L.L.; Truong, L.; Simonich, M.T.; Tanguay, R.L. Assessing the hazard of E-Cigarette flavor mixtures using zebrafish. Food Chem. Toxicol. 2020, 136, 110945. [Google Scholar] [CrossRef] [PubMed]
- Crosby, E.B.; Bailey, J.M.; Oliveri, A.N.; Levin, E.D. Neurobehavioral impairments caused by developmental imidacloprid exposure in zebrafish. Neurotoxicol. Teratol. 2015, 49, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H.; Souders, C.L., 2nd; Zhao, Y.H.; Martyniuk, C.J. Paraquat affects mitochondrial bioenergetics, dopamine system expression, and locomotor activity in zebrafish (Danio rerio). Chemosphere 2018, 191, 106–117. [Google Scholar] [CrossRef]
- Perez-Rodriguez, V.; Souders, C.L., 2nd; Tischuk, C.; Martyniuk, C.J. Tebuconazole reduces basal oxidative respiration and promotes anxiolytic responses and hypoactivity in early-staged zebrafish (Danio rerio). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 217, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Awoyemi, O.; Willis, A.; Schmitt, C.; Ramalingam, L.; Moustaid-Moussa, N.; Crago, J. Comparative Lipid Peroxidation and Apoptosis in Embryo-Larval Zebrafish Exposed to 3 Azole Fungicides, Tebuconazole, Propiconazole, and Myclobutanil, at Environmentally Relevant Concentrations. Environ. Toxicol. Chem. 2019, 38, 1455–1466. [Google Scholar] [CrossRef]
- Bailey, J.M.; Oliveri, A.N.; Karbhari, N.; Brooks, R.A.; De La Rocha, A.J.; Janardhan, S.; Levin, E.D. Persistent behavioral effects following early life exposure to retinoic acid or valproic acid in zebrafish. Neurotoxicology 2016, 52, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lei, L.; Tian, L.; Hou, F.; Roper, C.; Ge, X.; Zhao, Y.; Chen, Y.; Dong, Q.; Tanguay, R.L.; et al. Developmental and behavioral alterations in zebrafish embryonically exposed to valproic acid (VPA): An aquatic model for autism. Neurotoxicol. Teratol. 2018, 66, 8–16. [Google Scholar] [CrossRef]
- Cowden, J.; Padnos, B.; Hunter, D.; MacPhail, R.; Jensen, K.; Padilla, S. Developmental exposure to valproate and ethanol alters locomotor activity and retino-tectal projection area in zebrafish embryos. Reprod. Toxicol. 2012, 33, 165–173. [Google Scholar] [CrossRef]
- Zellner, D.; Padnos, B.; Hunter, D.L.; MacPhail, R.C.; Padilla, S. Rearing conditions differentially affect the locomotor behavior of larval zebrafish, but not their response to valproate-induced developmental neurotoxicity. Neurotoxicol. Teratol. 2011, 33, 674–679. [Google Scholar] [CrossRef]
- Jarema, K.A.; Hunter, D.L.; Shaffer, R.M.; Behl, M.; Padilla, S. Acute and developmental behavioral effects of flame retardants and related chemicals in zebrafish. Neurotoxicol. Teratol. 2015, 52, 194–209. [Google Scholar] [CrossRef] [Green Version]
- Padilla, S.; Hunter, D.L.; Padnos, B.; Frady, S.; MacPhail, R.C. Assessing motor activity in larval zebrafish: Influence of extrinsic and intrinsic variables. Neurotoxicol. Teratol. 2011, 33, 624–630. [Google Scholar] [CrossRef]
- Chen, T.H.; Wang, Y.H.; Wu, Y.H. Developmental exposures to ethanol or dimethylsulfoxide at low concentrations alter locomotor activity in larval zebrafish: Implications for behavioral toxicity bioassays. Aquat. Toxicol. 2011, 102, 162–166. [Google Scholar] [CrossRef]
- Christou, M.; Kavaliauskis, A.; Ropstad, E.; Fraser, T.W.K. DMSO effects larval zebrafish (Danio rerio) behavior, with additive and interaction effects when combined with positive controls. Sci. Total Environ. 2020, 709, 134490. [Google Scholar] [CrossRef]
- Huang, Y.; Cartlidge, R.; Walpitagama, M.; Kaslin, J.; Campana, O.; Wlodkowic, D. Unsuitable use of DMSO for assessing behavioral endpoints in aquatic model species. Sci. Total Environ. 2018, 615, 107–114. [Google Scholar] [CrossRef]
- Hill, B.N.; Coldsnow, K.D.; Hunter, D.L.; Hedge, J.M.; Korest, D.; Jarema, K.A.; Padilla, S. Assessment of Larval Zebrafish Locomotor Activity for Developmental Neurotoxicity Screening. In Experimental Neurotoxicology Methods; Llorens, J., Barenys, M., Eds.; Humana: New York, NY, USA, 2021; Volume 172, pp. 327–351. [Google Scholar] [CrossRef]
- Burgess, H.A.; Granato, M. The neurogenetic frontier--lessons from misbehaving zebrafish. Brief. Funct. Genom. Proteom. 2008, 7, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio Rerio), 4th ed.; University of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Burgess, H.A.; Granato, M. Modulation of locomotor activity in larval zebrafish during light adaptation. J. Exp. Biol. 2007, 210, 2526–2539. [Google Scholar] [CrossRef] [Green Version]
- Emran, F.; Rihel, J.; Adolph, A.R.; Wong, K.Y.; Kraves, S.; Dowling, J.E. OFF ganglion cells cannot drive the optokinetic reflex in zebrafish. Proc. Natl. Acad. Sci. USA 2007, 104, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.M.; Fero, K.; Arrenberg, A.B.; Bergeron, S.A.; Driever, W.; Burgess, H.A. Deep brain photoreceptors control light-seeking behavior in zebrafish larvae. Curr. Biol. 2012, 22, 2042–2047. [Google Scholar] [CrossRef] [Green Version]
- Prober, D.A.; Rihel, J.; Onah, A.A.; Sung, R.J.; Schier, A.F. Hypocretin/orexin overexpression induces an insomnia-like phenotype in zebrafish. J. Neurosci. 2006, 26, 13400–13410. [Google Scholar] [CrossRef]
- Tufi, S.; Leonards, P.; Lamoree, M.; de Boer, J.; Legler, J.; Legradi, J. Changes in Neurotransmitter Profiles during Early Zebrafish (Danio rerio) Development and after Pesticide Exposure. Environ. Sci. Technol. 2016, 50, 3222–3230. [Google Scholar] [CrossRef]
- MacPhail, R.C.; Brooks, J.; Hunter, D.L.; Padnos, B.; Irons, T.D.; Padilla, S. Locomotion in larval zebrafish: Influence of time of day, lighting and ethanol. Neurotoxicology 2009, 30, 52–58. [Google Scholar] [CrossRef]
- Baker, N.; Knudsen, T.; Williams, A. Abstract Sifter: A comprehensive front-end system to PubMed. F1000Research 2017, 6, 2164. [Google Scholar] [CrossRef] [Green Version]
- Hamm, J.T.; Ceger, P.; Allen, D.; Stout, M.; Maull, E.A.; Baker, G.; Zmarowski, A.; Padilla, S.; Perkins, E.; Planchart, A.; et al. Characterizing sources of variability in zebrafish embryo screening protocols. ALTEX 2019, 36, 103–120. [Google Scholar] [CrossRef]
- Legradi, J.; el Abdellaoui, N.; van Pomeren, M.; Legler, J. Comparability of behavioural assays using zebrafish larvae to assess neurotoxicity. Environ. Sci. Pollut. Res. 2015, 22, 16277–16289. [Google Scholar] [CrossRef]
- Goolish, E.M.; Okutake, K. Lack of gas bladder inflation by the larvae of zebrafish in the absence of an air-water interface. J. Fish Biol. 1999, 55, 1054–1063. [Google Scholar] [CrossRef]
- Amrhein, V.; Greenland, S.; McShane, B. Scientists rise up against statistical significance. Nature 2019, 567, 305–307. [Google Scholar] [CrossRef] [Green Version]
- Nuzzo, R. Scientific method: Statistical errors. Nature 2014, 506, 150–152. [Google Scholar] [CrossRef] [Green Version]
- Haigis, A.C.; Ottermanns, R.; Schiwy, A.; Hollert, H.; Legradi, J. Getting more out of the zebrafish light dark transition test. Chemosphere 2022, 295, 133863. [Google Scholar] [CrossRef]
- Hsieh, J.H.; Ryan, K.; Sedykh, A.; Lin, J.A.; Shapiro, A.J.; Parham, F.; Behl, M. Application of Benchmark Concentration (BMC) Analysis on Zebrafish Data: A New Perspective for Quantifying Toxicity in Alternative Animal Models. Toxicol. Sci. 2019, 167, 92–104. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, P.; Cassidy, P.A.; Carmer, R.; Zhang, G.; Venkatraman, P.; Brown, S.A.; Pang, C.P.; Zhong, W.; Zhang, M.; et al. Statistical Analysis of Zebrafish Locomotor Behaviour by Generalized Linear Mixed Models. Sci. Rep. 2017, 7, 2937. [Google Scholar] [CrossRef]
- Ellis, L.D.; Seibert, J.; Soanes, K.H. Distinct models of induced hyperactivity in zebrafish larvae. Brain Res. 2012, 1449, 46–59. [Google Scholar] [CrossRef]
- Irons, T.D.; MacPhail, R.C.; Hunter, D.L.; Padilla, S. Acute neuroactive drug exposures alter locomotor activity in larval zebrafish. Neurotoxicol. Teratol. 2010, 32, 84–90. [Google Scholar] [CrossRef]
- Powell, S.B.; Weber, M.; Geyer, M.A. Genetic models of sensorimotor gating: Relevance to neuropsychiatric disorders. Curr. Top. Behav. Neurosci. 2012, 12, 251–318. [Google Scholar] [CrossRef] [Green Version]
- Hanswijk, S.I.; Spoelder, M.; Shan, L.; Verheij, M.M.M.; Muilwijk, O.G.; Li, W.; Liu, C.; Kolk, S.M.; Homberg, J.R. Gestational Factors throughout Fetal Neurodevelopment: The Serotonin Link. Int. J. Mol. Sci. 2020, 21, 5850. [Google Scholar] [CrossRef]
- Cerne, R.; Lippa, A.; Poe, M.M.; Smith, J.L.; Jin, X.; Ping, X.; Golani, L.K.; Cook, J.M.; Witkin, J.M. GABAkines-Advances in the discovery, development, and commercialization of positive allosteric modulators of GABAA receptors. Pharmacol. Ther. 2021, 16, 108035. [Google Scholar] [CrossRef]
- Bastías-Candia, S.; Zolezzi, J.M.; Inestrosa, N.C. Revisiting the Paraquat-Induced Sporadic Parkinson’s Disease-Like Model. Mol. Neurobiol. 2019, 56, 1044–1055. [Google Scholar] [CrossRef]
- Finney, J.L.; Robertson, G.N.; McGee, C.A.; Smith, F.M.; Croll, R.P. Structure and autonomic innervation of the swim bladder in the zebrafish (Danio rerio). J. Comp. Neurol. 2006, 495, 587–606. [Google Scholar] [CrossRef]
- Robertson, G.N.; McGee, C.A.; Dumbarton, T.C.; Croll, R.P.; Smith, F.M. Development of the swimbladder and its innervation in the zebrafish, Danio rerio. J. Morphol. 2007, 268, 967–985. [Google Scholar] [CrossRef]
- Lindsey, B.W.; Smith, F.M.; Croll, R.P. From inflation to flotation: Contribution of the swimbladder to whole-body density and swimming depth during development of the zebrafish (Danio rerio). Zebrafish 2010, 7, 85–96. [Google Scholar] [CrossRef]
- Martin, M.M.; Baker, N.C.; Boyes, W.K.; Carstens, K.E.; Culbreth, M.E.; Gilbert, M.E.; Harrill, J.A.; Nyffeler, J.; Padilla, S.; Paul Friedman, K.; et al. An expert-driven literature review of “negative” reference chemicals for developmental neurotoxicity (DNT) in vitro assay evaluation. Neurotoxicol. Teratol. 2021; submitted. [Google Scholar]


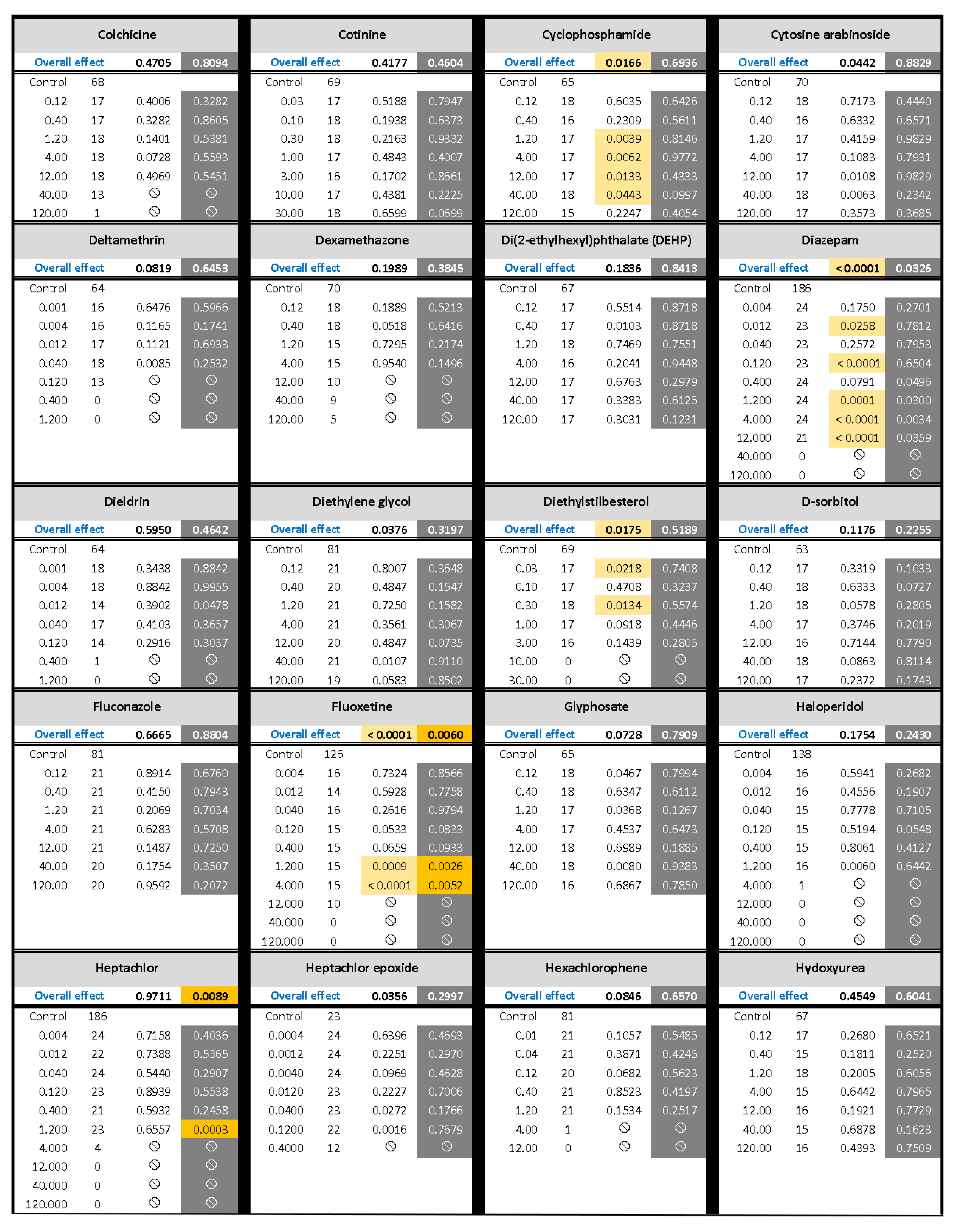
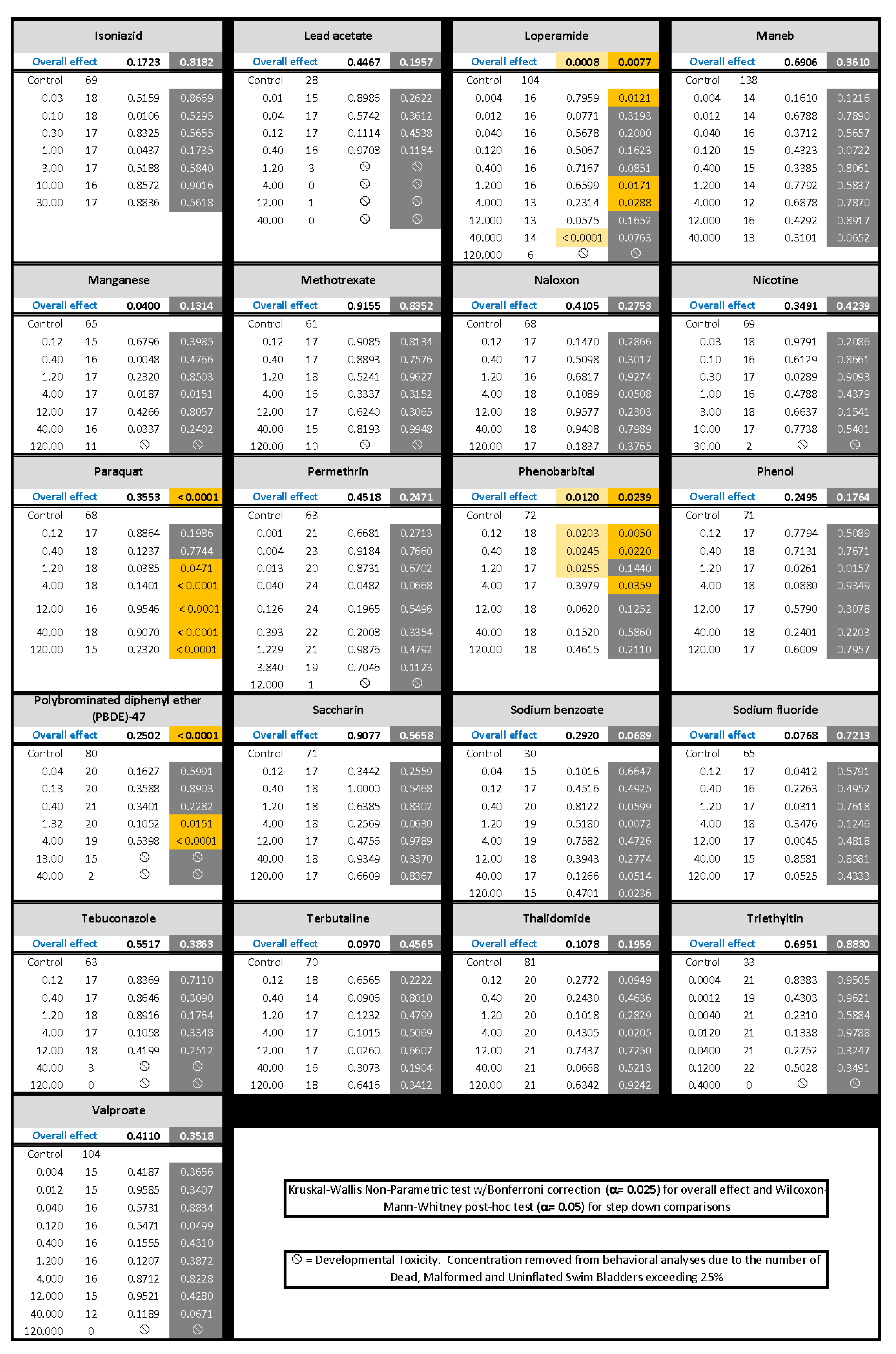
 ) indicates developmental toxicity: that concentration was not included in behavioral analyses due to the number of dead, malformed and uninflated swim bladders exceeding 25%. Overall effect is listed under the chemical name followed by the sample size and results for each concentration, with the Dark period shaded gray. Statistically significant results are highlighted with the light-yellow shading in the Light period and dark-yellow shading in the Dark period.
) indicates developmental toxicity: that concentration was not included in behavioral analyses due to the number of dead, malformed and uninflated swim bladders exceeding 25%. Overall effect is listed under the chemical name followed by the sample size and results for each concentration, with the Dark period shaded gray. Statistically significant results are highlighted with the light-yellow shading in the Light period and dark-yellow shading in the Dark period.
) indicates developmental toxicity: that concentration was not included in behavioral analyses due to the number of dead, malformed and uninflated swim bladders exceeding 25%. Overall effect is listed under the chemical name followed by the sample size and results for each concentration, with the Dark period shaded gray. Statistically significant results are highlighted with the light-yellow shading in the Light period and dark-yellow shading in the Dark period.
) indicates developmental toxicity: that concentration was not included in behavioral analyses due to the number of dead, malformed and uninflated swim bladders exceeding 25%. Overall effect is listed under the chemical name followed by the sample size and results for each concentration, with the Dark period shaded gray. Statistically significant results are highlighted with the light-yellow shading in the Light period and dark-yellow shading in the Dark period.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical | Cas # | DTXSID | Molecular Weight | Solvent | Water Solubility (µmol/L) Predicted Median | Water Solubility (µmol/L) Predicted Range | Octanol Water Coeff (LogKow) Predicted Median | Octanol Water Coeff (LogKow) Predicted Range |
|---|---|---|---|---|---|---|---|---|
| 5, 5-Diphenylhydantoin | 57-41-0 | DTXSID8020541 | 252.3 | DMSO | 5.67 × 104 | 1.07 × 102 to 6.00 × 106 | 2.39 | 2.16 to 2.52 |
| 5-Fluorouracil | 51-21-8 | DTXSID2020634 | 130.1 | DMSO | 1.44 × 105 | 3.07 × 104 to 3.69 × 106 | −0.906 | −1.37 to −0.810 |
| 6-Aminonicotinamide | 329-89-5 | DTXSID5051446 | 137.1 | DMSO | 6.68×104 | 6.41 × 104 to 1.28 × 105 | 0.027 | −0.730 to 0.698 |
| 6-Propyl-2-thiouracil | 51-52-5 | DTXSID5021209 | 170.2 | DMSO | 3.00 × 104 | 6.93 × 103 to 4.98 × 106 | 0.523 | −0.386 to 1.37 |
| Acetaminophen | 103-90-2 | DTXSID2020006 | 151.1 | DMSO | 1.47 × 105 | 3.95 × 104 to 5.70 × 106 | 0.372 | 0.270 to 0.462 |
| Acrylamide | 79-06-1 | DTXSID5020027 | 71.1 | DMSO | 7.05 × 106 | 2.66 × 106 to 8.99 × 106 | −0.726 | −0.810 to −0.670 |
| Aldicarb | 116-06-3 | DTXSID0039223 | 190.3 | DMSO | 2.79 × 104 | 2.55 × 104 to 3.03 × 104 | 1.13 | 1.13 to 1.36 |
| Amoxicillin | 26787-78-0 | DTXSID3037044 | 365.4 | DMSO | 9.36 × 103 | 5.58 × 103 to 4.93 × 106 | 0.742 | 0.48 to 0.97 |
| Amphetamine | 51-63-8 | DTXSID2057865 | 184.3 | H2O | 5.70 × 106 | 1.33 × 105 to 1.13 × 107 | 1.81 | 0.602 to 1.82 |
| Arsenic | 7784-46-5 | DTXSID5020104 | 129.9 | H2O | - | - | −3.28 | −3.28 |
| Bisphenol A (BPA) | 80-05-7 | DTXSID7020182 | 228.3 | DMSO | 1.00 × 103 | 7.45 × 102 to 6.76 × 106 | 3.53 | 3.32 to 3.64 |
| Bis(tributyltin) Oxide | 56-35-9 | DTXSID9020166 | 596.1 | DMSO | 1.5 × 10−1 | 1.5 × 10−1 | 4.05 | 4.05 |
| Cadmium chloride | 654054-66-7 | - | 183.3 | DMSO | - | - | - | - |
| Caffeine | 58-08-2 | DTXSID0020232 | 194.2 | DMSO | 8.30 × 104 | 1.36× 104 to 7.14 × 106 | 0.045 | −0.131 to 0.283 |
| Captopril | 62571-86-2 | DTXSID1037197 | 217.2 | DMSO | 9.47 × 104 | 3.98 × 104 to 2.46 × 106 | 0.481 | 0.272 to 0.840 |
| Carbamazepine | 298-46-4 | DTXSID4022731 | 236.3 | DMSO | 2.83 × 102 | 2.55 × 101 to 7.00 × 106 | 2.37 | 2.25 to 2.67 |
| Chloramben | 133-90-4 | DTXSID2020262 | 206.0 | DMSO | 3.40 × 103 | 2.92 × 103 to 4.68 × 103 | 2.15 | 0.912 to 2.52 |
| Chlorpyrifos (ethyl) | 2921-88-2 | DTXSID4020458 | 350.6 | DMSO | 2.83 | 1.02 to 7.00 × 106 | 4.78 | 4.66 to 4.96 |
| Chlorpyrifos (ethyl) oxon | 5598-15-2 | DTXSID1038666 | 334.5 | DMSO | 2.10 × 102 | 7.76 × 101 to 2.26 × 102 | 3.32 | 2.89 to 3.73 |
| Cocaine base | 50-36-2 | DTXSID2038443 | 184.3 | H2O | 4.93 × 106 | 5.73 × 103 to 9.85 × 106 | 2.79 | 2.3 to 3.08 |
| Colchicine | 64-86-8 | DTXSID5024845 | 399.4 | DMSO | 5.65 × 104 | 5.25 × 102 to 7.00 × 106 | 1.2 | 0.920 to 1.86 |
| Cotinine | 486-56-6 | DTXSID1047576 | 176.2 | DMSO | 2.99 × 106 | 3.70 × 104 to 9.02 × 106 | 0.119 | −0.228 to 0.340 |
| Cyclophosphamide | 6055-19-2 | DTXSID6024888 | 279.1 | H2O | 1.52 × 105 | 5.58 × 104 to 8.02 × 106 | 0.526 | 0.230 to 1.30 |
| Cytosine arabinoside | 147-94-4 | DTXSID3022877 | 243.2 | DMSO | 4.54 × 105 | 4.39 × 104 to 8.32 × 106 | −2.32 | −2.51 to −1.94 |
| Deltamethrin | 52918-63-5 | DTXSID8020381 | 505.2 | DMSO | 1.96 × 10−2 | 1.86 × 10−3 to 7.00 106 | 6.19 | 6.12 to 6.20 |
| Dexamethasone | 50-02-2 | DTXSID3020384 | 392.4 | DMSO | 1.95 × 102 | 1.05 × 102 to 7.00 × 106 | 1.89 | 1.72 to 1.92 |
| Di(2-ethylhexyl)phthalate (DEHP) | 117-81-7 | DTXSID5020607 | 390.6 | DMSO | 4.23 × 10−1 | 2.90 × 10−3 to 7.00 × 106 | 8.15 | 7.52 to 8.71 |
| Diazepam | 439-14-5 | DTXSID4020406 | 284.7 | DMSO | 1.91 × 102 | 1.07 × 102 to 7.07 × 106 | 2.91 | 2.70 to 2.92 |
| Dieldrin | 60-57-1 | DTXSID9020453 | 380.9 | DMSO | 1.57 | 5.42 × 10−1 to 2.60 | 4.94 | 4.88 to 5.12 |
| Diethylene Glycol | 111-46-6 | DTXSID8020462 | 106.1 | DMSO | 6.51 × 106 | 5.40 × 106 to 9.42 × 106 | −1.28 | −1.51 to −1.09 |
| Diethyl-stilbesterol | 56-53-1 | DTXSID3020465 | 268.4 | DMSO | 4.37 × 101 | 1.24 × 101 to 6.88 × 106 | 5.35 | 4.80 to 5.93 |
| D-sorbitol | 50-70-4 | DTXSID5023588 | 182.2 | DMSO | 3.31 × 106 | 1.72 × 106 to 6.07 × 106 | −3.15 | −4.67 to −2.38 |
| Fluconazole | 86386-73-4 | DTXSID3020627 | 306.2 | DMSO | 9.68 × 103 | 1.35 × 103 to 7.15 × 106 | 0.501 | 0.250 to 0.698 |
| Fluoxetine | 56296-78-7 | DTXSID7020635 | 345.8 | DMSO | 1.94 × 102 | 2.37 × 101 to 1.02 × 107 | 4.09 | 0.768 to 4.23 |
| Glyphosate | 1071-83-6 | DTXSID1024122 | 169.1 | H2O | 1.99 × 106 | 6.56 × 104 to 8.41 × 106 | −2.88 | −4.47 to −2.26 |
| Haloperidol | 52-86-8 | DTXSID4034150 | 375.9 | DMSO | 3.10 × 101 | 2.34 × 101 to 9.11 × 106 | 3.84 | 3.01 to 4.29 |
| Heptachlor | 76-44-8 | DTXSID3020679 | 373.3 | DMSO | 9.25 × 10−2 | 7.39 × 10−2 to 3.82 × 10−1 | 5.7 | 5.46 to 6.10 |
| Heptachlor epoxide | 1024-57-3 | DTXSID1024126 | 389.3 | DMSO | 5.68 × 10−1 | 5.68 × 10−1 | 5.29 | 4.98 to 5.47 |
| Hexachlorophene | 70-30-4 | DTXSID6020690 | 406.9 | DMSO | 1.73 × 102 | 9.43× 10−3 to 6.66 × 106 | 7.23 | 6.92 to 7.54 |
| Hydoxy-urea | 127-07-1 | DTXSID6025438 | 76.1 | DMSO | 5.42 × 106 | 2.95 × 106 to 1.32 × 107 | −1.74 | −1.80 to −1.54 |
| Isoniazid | 54-85-3 | DTXSID8020755 | 137.1 | DMSO | 7.66 × 105 | 1.22 × 105 to 7.32 × 106 | 0.754 | −0.887 to −0.635 |
| Lead acetate | 6080-56-4 | DTXSID3031521 | 379.3 | H2O | 7.77 × 106 | 2.14 × 106 to 1.34 × 107 | −0.285 | −2.21 to −7.10 x 10−2 |
| Loperamide | 34552-83-5 | DTXSID00880006 | 513.5 | DMSO | 4.46 × 106 | 2.21 × 101 to 8.91 × 106 | 4.26 | 1.32 to 4.47 |
| Maneb | 12427-38-2 | DTXSID9020794 | 265.3 | DMSO | 1.01 × 106 | 7.72 × 105 to 1.25 × 106 | 1.4 | −2.70 to 1.66 |
| Manganese | 7773-01-5 | DTXSID9040681 | 126.0 | H2O | - | - | - | - |
| Methotrexate | 59-05-2 | DTXSID4020822 | 454.4 | DMSO | 3.2 × 103 | 1.89 × 102 to 5.37 × 106 | −0.922 | −1.85 to −0.241 |
| Naloxon | 51481-60-8 | DTXSID90199452 | 399.9 | H2O | 4.06 × 103 | 2.74 × 103 to 7.99 × 106 | 1.45 | 0.243 to 1.53 |
| Nicotine | 54-11-5 | DTXSID1020930 | 162.2 | DMSO | 6.15 × 106 | 8.00 × 104 to 1.10 × 107 | 0.91 | 0.720 to 1.17 |
| Paraquat | 1910-42-5 | DTXSID7024243 | 257.1 | H2O | 4.88 × 106 | 2.76 × 106 to 7.00 × 106 | −4.58 | −5.11 to −4.50 |
| Permethrin | 52645-53-1 | DTXSID8022292 | 391.2 | DMSO | 1.32 × 10−1 | 2.49 × 10−2 to 7.00 × 106 | 6.82 | 6.47 to 7.43 |
| Phenobarbital | 57-30-7 | DTXSID0021123 | 254.2 | DMSO | 1.68 × 104 | 1.02 × 104 to 3.89 × 105 | −0.285 | −2.29 to 1.13 |
| Phenol | 108-95-2 | DTXSID5021124 | 94.1 | DMSO | 6.04 × 105 | 2.78 × 105 to 4.91 × 106 | 1.5 | 1.46 to 1.63 |
| Polybrominated diphenyl ether (PBDE)-47 | 5436-43-1 | DTXSID3030056 | 485.8 | DMSO | 5.61 × 10−3 | 3.01 × 10−3 to 1.23 × 10−1 | 6.79 | 6.59 to 7.39 |
| Saccharin | 82385-42-0 | DTXSID7021992 | 205.1 | DMSO | 1.91 × 104 | 9.43 × 103 to 1.85 × 106 | 0.705 | −2.01 to 0.910 |
| Sodium benzoate | 532-32-1 | DTXSID1020140 | 144.1 | H2O | 3.32 × 105 | 6.44 × 104 to 2.84 × 106 | 0.158 | −2.27 to 1.90 |
| Sodium fluoride | 7681-49-4 | DTXSID2020630 | 42.0 | H2O | 1.42 × 107 | 1.42 × 107 | −0.77 | −0.77 |
| Tebuconazole | 107534-96-3 | DTXSID9032113 | 307.8 | DMSO | 1.03 × 102 | 8.04 × 101 to 7.09 × 106 | 3.72 | 3.58 to 3.89 |
| Terbutaline | 23031-32-5 | DTXSID3045437 | 274.3 | DMSO | 4.71.× 106 | 4.63 × 104 to 9.37 × 106 | 0.477 | 0.439 to 0.523 |
| Thalidomide | 50-35-1 | DTXSID9022524 | 258.2 | DMSO | 1.74 × 103 | 6.49 × 102 to 6.42 × 106 | 0.405 | −0.240 to 0.541 |
| Triethyltin | 2767-54-6 | DTXSID9040712 | 285.8 | DMSO | 1.38 × 103 | 1.38 × 103 | 1.84 | 1.84 |
| Valproate | 99-66-1 | DTXSID6023733 | 144.2 | DMSO | 1.99 × 104 | 6.20 × 103 to 3.33 × 106 | 2.73 | 2.65 to 2.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jarema, K.A.; Hunter, D.L.; Hill, B.N.; Olin, J.K.; Britton, K.N.; Waalkes, M.R.; Padilla, S. Developmental Neurotoxicity and Behavioral Screening in Larval Zebrafish with a Comparison to Other Published Results. Toxics 2022, 10, 256. https://doi.org/10.3390/toxics10050256
Jarema KA, Hunter DL, Hill BN, Olin JK, Britton KN, Waalkes MR, Padilla S. Developmental Neurotoxicity and Behavioral Screening in Larval Zebrafish with a Comparison to Other Published Results. Toxics. 2022; 10(5):256. https://doi.org/10.3390/toxics10050256
Chicago/Turabian StyleJarema, Kimberly A., Deborah L. Hunter, Bridgett N. Hill, Jeanene K. Olin, Katy N. Britton, Matthew R. Waalkes, and Stephanie Padilla. 2022. "Developmental Neurotoxicity and Behavioral Screening in Larval Zebrafish with a Comparison to Other Published Results" Toxics 10, no. 5: 256. https://doi.org/10.3390/toxics10050256
APA StyleJarema, K. A., Hunter, D. L., Hill, B. N., Olin, J. K., Britton, K. N., Waalkes, M. R., & Padilla, S. (2022). Developmental Neurotoxicity and Behavioral Screening in Larval Zebrafish with a Comparison to Other Published Results. Toxics, 10(5), 256. https://doi.org/10.3390/toxics10050256