
Insights into the Seasonal Olfactory Mechanism of Geosmin in Raw Water of Huangpu River



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Pretreatment
2.2. Determination of Geosmin
2.3. DNA Extraction and High-Throughput Sequencing
2.4. Sequence Processing
2.5. Data Analysis
3. Results and Discussion
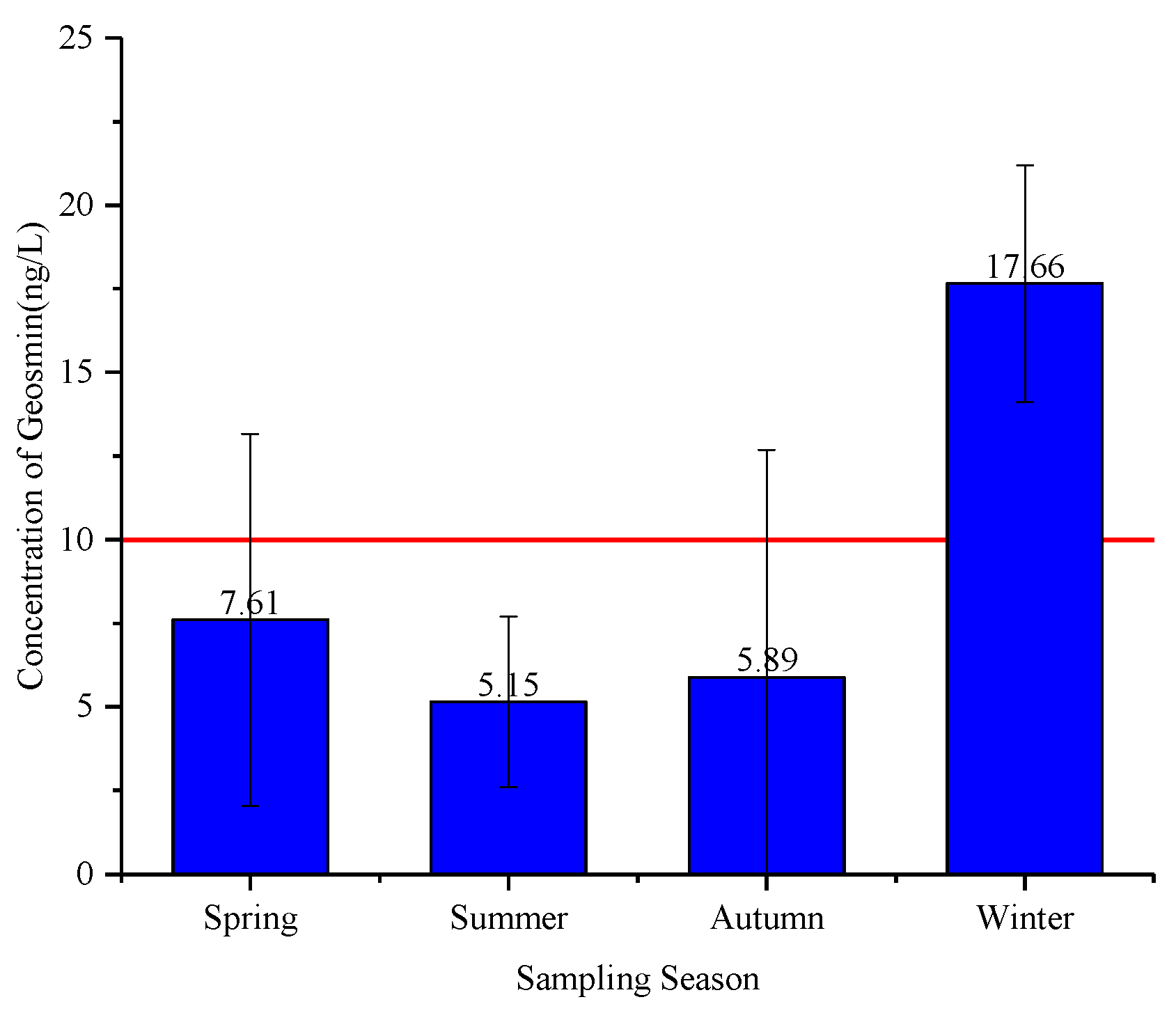
3.1. Seasonal Variation in Geosmin in Raw Water
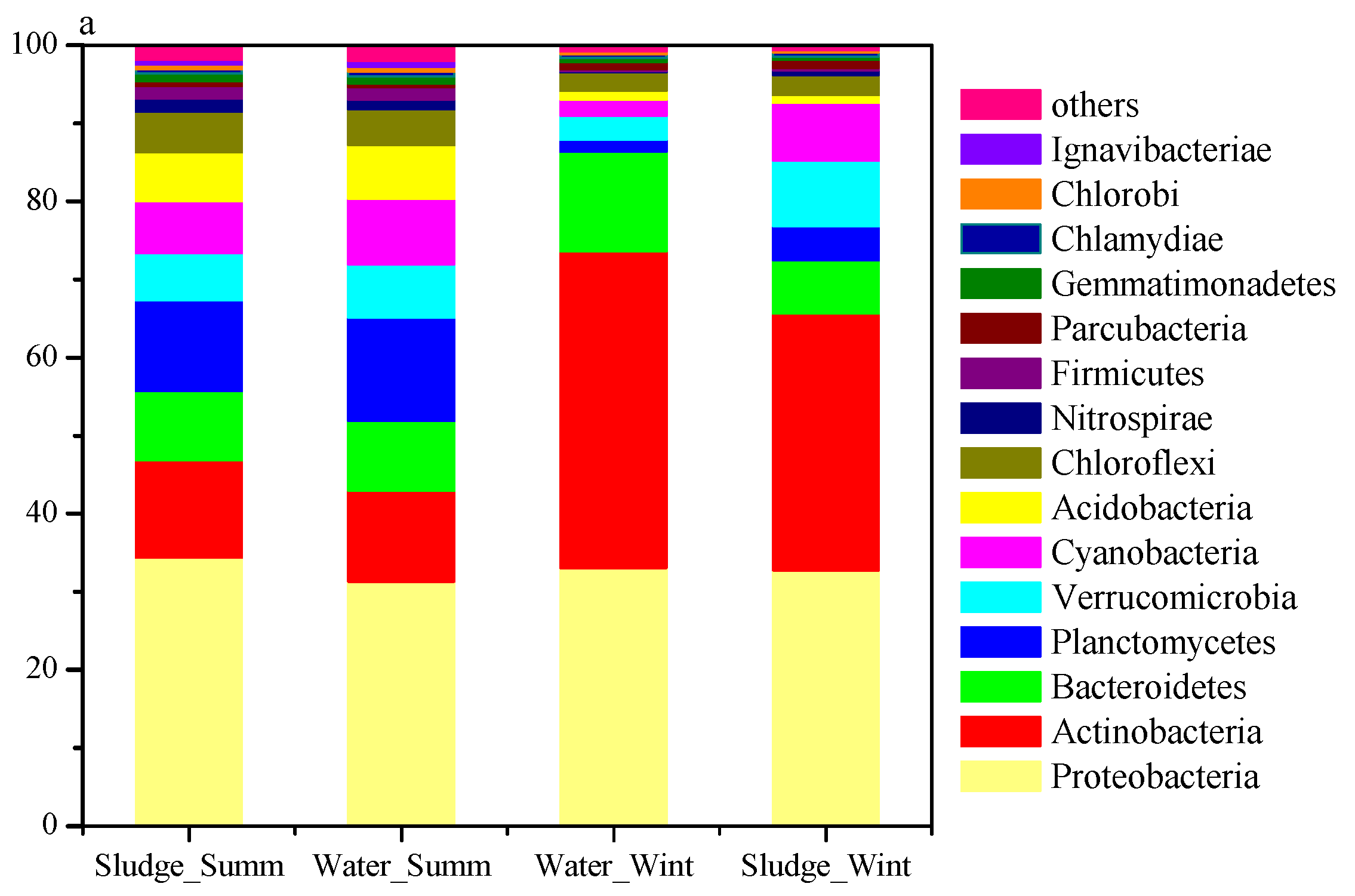
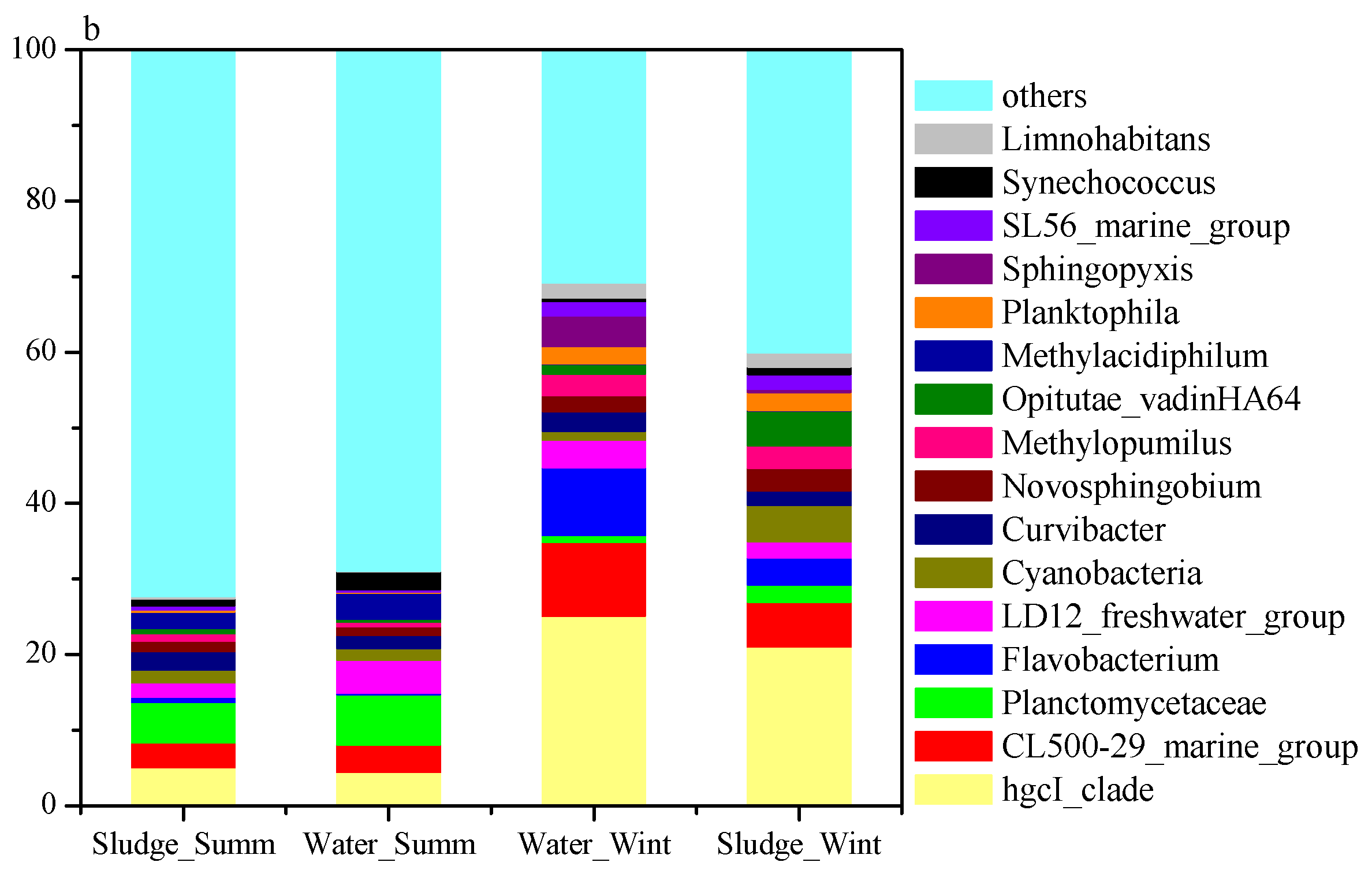
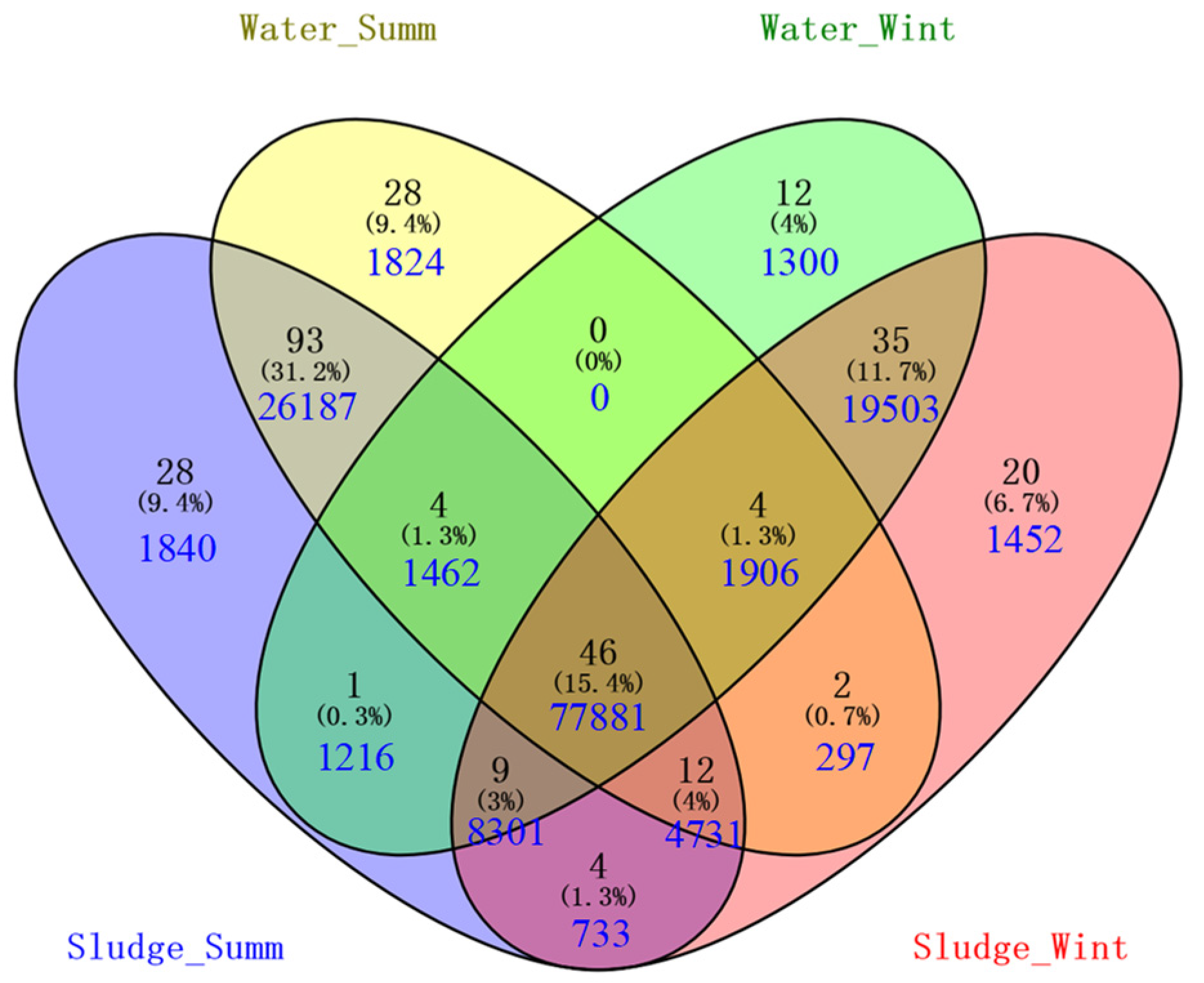
3.2. Sequencing of Microbes and Their Diversity
3.3. Seasonal Variation in Microbial Communities
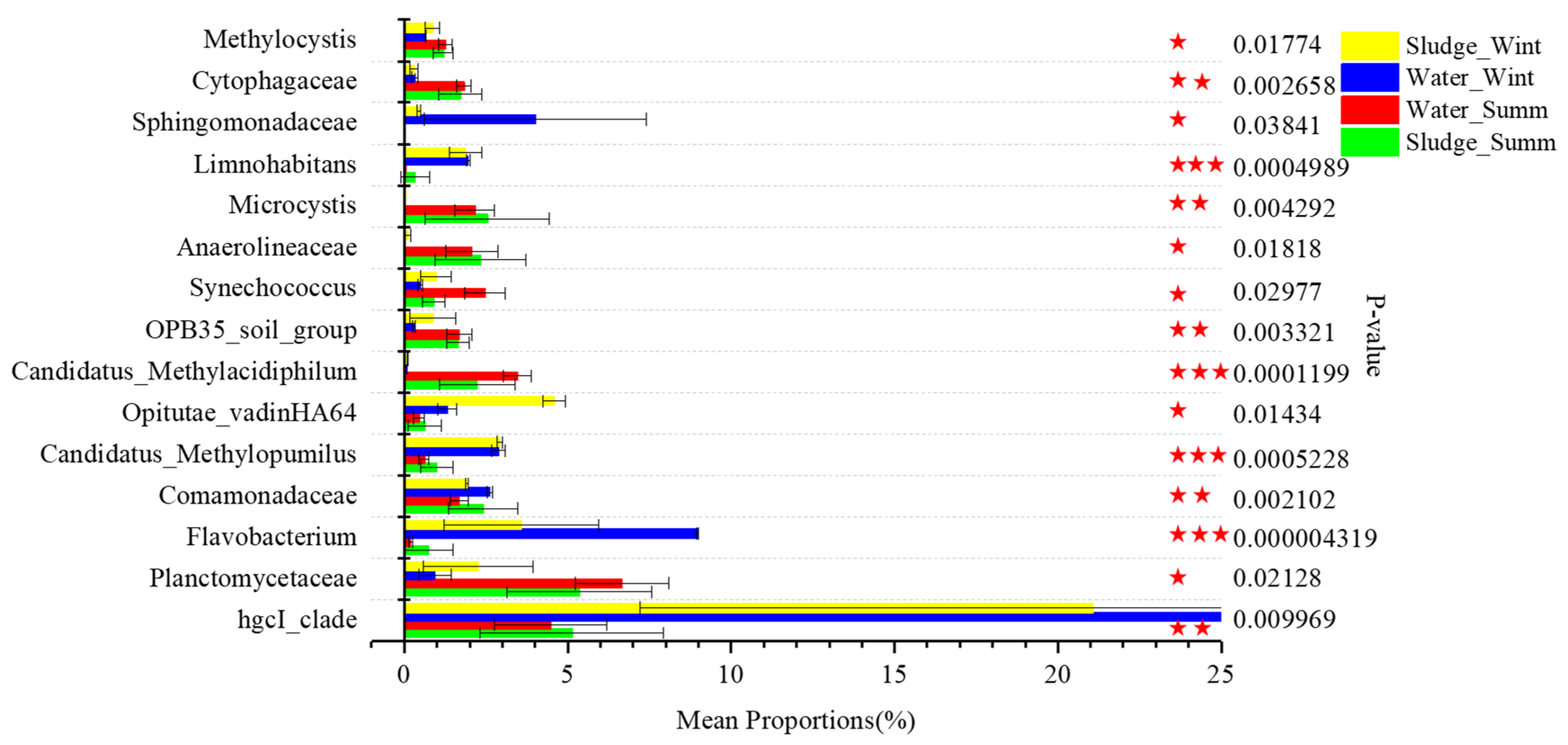
3.4. Screening of Potential Odorous Producers
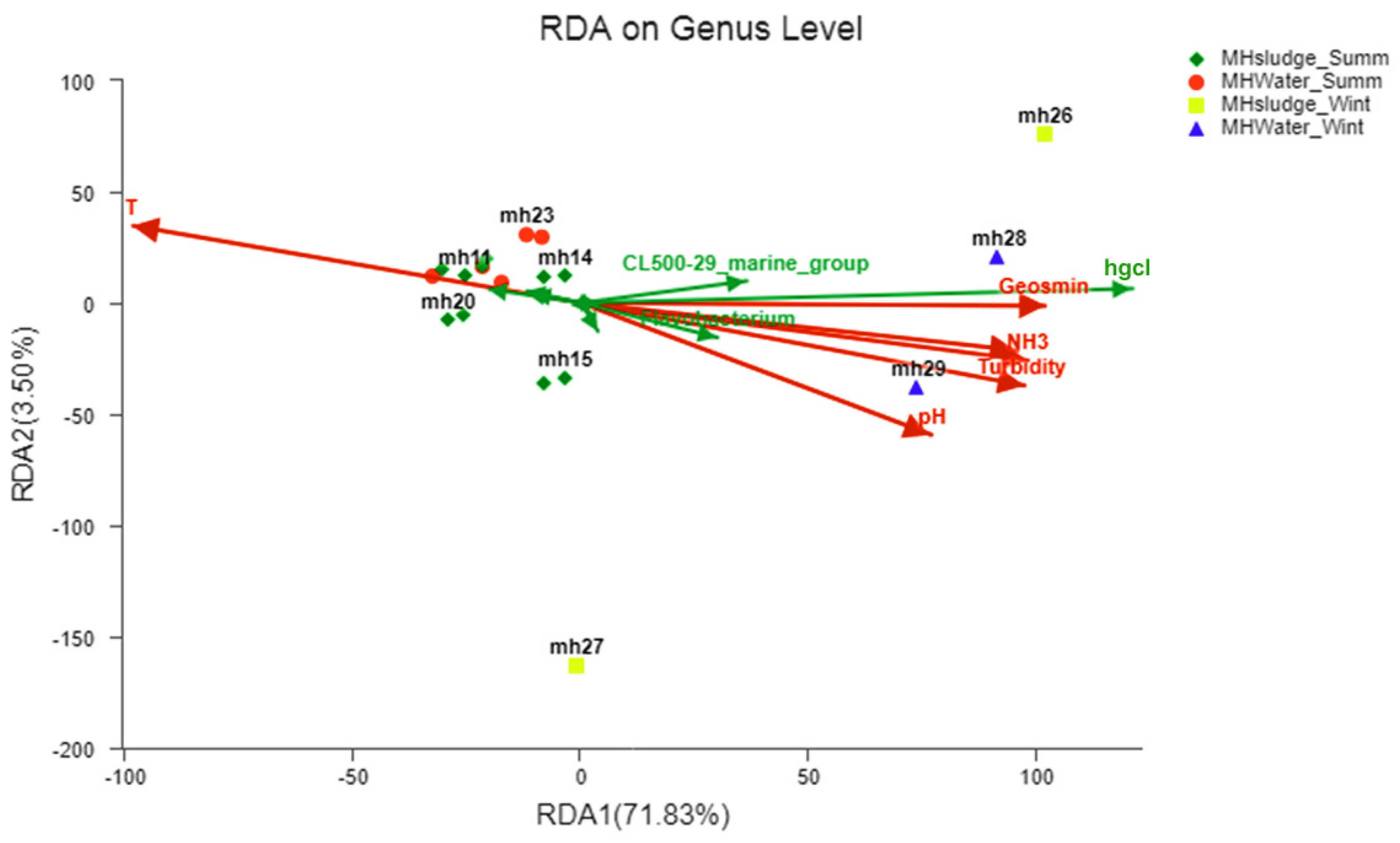
3.5. Effects of Environmental Parameters on Microbial Communities
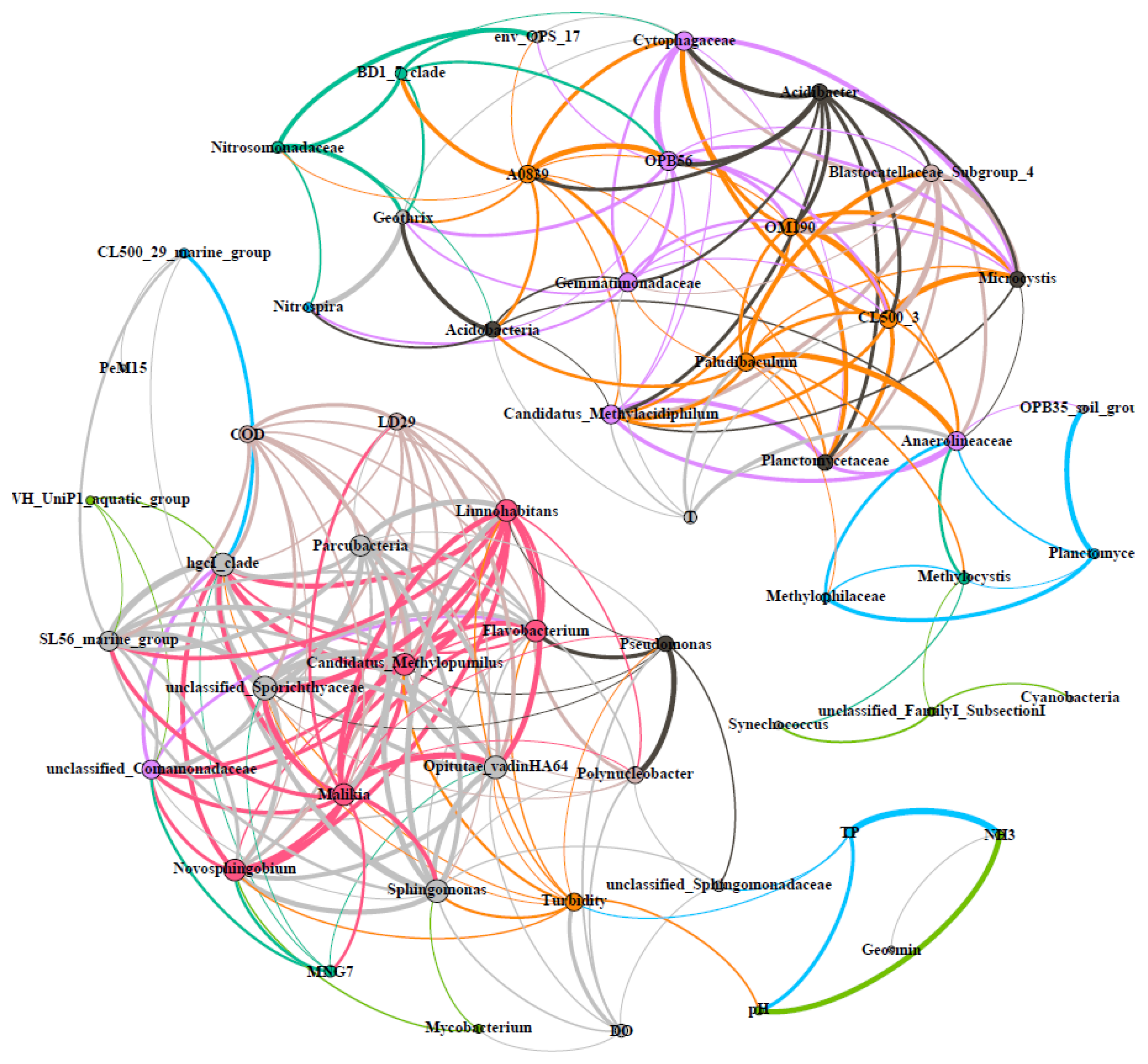
3.6. Co-Occurrence Correlations of Microorganisms and Environmental Parameters
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerber, N.N.; Lechevalier, H.A. Geosmin, an earthly-smelling substance isolated from actinomycetes. Appl. Microbiol. 1965, 13, 935–938. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Niu, Y.; Xie, P.; Chen, J.; Tao, M.; Deng, X. Off-flavor compounds from decaying cyanobacterial blooms of Lake Taihu. J. Environ. Sci. 2013, 25, 495–501. [Google Scholar] [CrossRef]
- Beniwal, D.; Taylor-Edmonds, L.; Armour, J.; Andrews, R.C. Ozone/peroxide advanced oxidation in combination with biofiltration for taste and odour control and organics removal. Chemosphere 2018, 212, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulou, M.; Evgenidou, E.; Lambropoulou, D.; Konstantinou, I. A review on advanced oxidation processes for the removal of taste and odor compounds from aqueous media. Water Res. 2014, 53, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Jüttner, F.; Watson, S.B. Biochemical and ecological control of geosmin and 2-methylisoborneol in source waters. Appl. Environ. Microb. 2007, 73, 4395–4406. [Google Scholar] [CrossRef]
- Su, M.; Suruzzaman, M.D.; Zhu, Y.P.; Lu, J.P.; Yu, J.W.; Zhang, Y.; Yang, M. Ecological niche and in-situ control of MIB producers in source water. J. Envrion. Sci.-China 2021, 110, 119–128. [Google Scholar] [CrossRef]
- Olsen, B.K.; Chislock, M.F.; Wilson, A.E. Eutrophication mediates a common off-flavor compound (2-methylisoborneol, in a drinking water reservoir. Water Res. 2016, 92, 228–234. [Google Scholar] [CrossRef]
- Devi, A.; Chiu, Y.T.; Hsueh, H.T.; Lin, T.F. Quantitative PCR based detection system for cyanobacterial geosmin/2-methylisoborneol (2-MIB) events in drinking water sources: Current status and challenges. Water Res. 2021, 188, 116478. [Google Scholar] [CrossRef]
- Mustapha, S.; Tijani, J.O.; Ndamitso, M.M.; Abdulkareem, A.S.; Shuaib, D.T.; Mohammed, A.K. A critical review on geosmin and 2-methylisoborneol in water: Sources, effects, detection, and removal techniques. Environ. Monit. Assess 2021, 193, 204. [Google Scholar] [CrossRef]
- Zaitlin, B.; Watson, S.B. Actinomycetes in relation to taste and odour in drinking water: Myths, tenets and truths. Water Res. 2006, 40, 1741–1753. [Google Scholar] [CrossRef]
- Ma, X.; Feng, J.; Song, Y.; Ni, M.; Dietrich, A.M.; Chen, C.; Li, Q.; Gao, N. Release behavior of odor contaminants derived from Microcystis aeruginosa in rivers and a non-strict anaerobic aqueous system. J. Water Supply Res. Technol. 2015, 64, 812–823. [Google Scholar] [CrossRef]
- Clercin, N.A.; Druschel, G.K.; Gray, M. Occurrences of 2-methylisoborneol and geosmin –degrading bacteria in a eutrophic reservoir and the role of cell-bound versus dissolved fractions. J. Envrion. Manag. 2021, 297, 113304. [Google Scholar] [CrossRef] [PubMed]
- Durrer, M.; Zimmermann, U.; Jüttner, F. Dissolved and particle-bound geosmin in a mesotrophic lake (lake Zürich): Spatial and seasonal distribution and the effect of grazers. Water Res. 1999, 33, 3628–3636. [Google Scholar] [CrossRef]
- Rice, E.; Baird, R.; Eaton, A.; Clesceri, L. Standard Methods for the Examination of Water and Wastewater; American Public Health Association, American Water Works Association, and Water Environment Federation: Washionton, DC, USA; Cenveo Publisher Services: Richmond, VA, USA, 2012. [Google Scholar]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microb. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, H.; Wang, Z.; Liu, R.; Zhang, Y.; Lin, K. Insights into spatially and temporally co-occurring polybrominated diphenyl ethers in sediments of the East China Sea. Chemosphere 2015, 123, 55–63. [Google Scholar] [CrossRef]
- Jiang, Y.; Cheng, B.; Liu, M.; Nie, Y. Spatial and temporal variations of taste and odor compounds in surface water, overlying water and sediment of the western Lake Chaohu, China. Bull. Environ. Contam. Toxicol. 2016, 96, 186–191. [Google Scholar] [CrossRef]
- Wang, Z.; Li, R. Effects of light and temperature on the odor production of 2-methylisoborneol-producing Pseudanabaena sp. and geosmin-producing Anabaena ucrainica (cyanobacteria). Biochem. Syst. Ecol. 2015, 58, 219–226. [Google Scholar] [CrossRef]
- Zhang, T.; Lin, L.; Song, L.R.; Wei, C. Effects of temperature and light on the growth and geosmin production of Lyngbya kuetzingii (Cyanophyta). J. Appl. Phycol. 2009, 21, 279–285. [Google Scholar] [CrossRef]
- Li, L.; Yang, S.; Yu, S.; Zhang, Y. Variation and removal of 2-MIB in full-scale treatment plants with source water from Lake Tai, China. Water Res. 2019, 162, 180–189. [Google Scholar] [CrossRef]
- Sugiura, N.; Iwami, N.; Inamori, Y.; Nishimura, O.; Sudo, R. Significance of attached cyanobacteria relevant to the occurrence of musty odor in Lake Kasumigaura. Water Res. 1998, 32, 3549–3554. [Google Scholar] [CrossRef]
- Zeng, D.N.; Fan, Z.Y.; Chi, L.; Wang, X.; Qu, W.D.; Quan, Z.X. Analysis of the bacterial communities associated with different drinking water treatment processes. World J. Microb. Biotechnol. 2013, 29, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Ávila, M.P.; Staehr, P.A.; Barbosa, F.A.R.; Chartone-Souza, E.; Nascimento, A.M.A. Seasonality of freshwater bacterioplankton diversity in two tropical shallow lakes from the Brazilian Atlantic Forest. FEMS Microb. Ecol. 2017, 93, fiw218. [Google Scholar] [CrossRef]
- Alghanmi, H.A.; Alkam, F.M.; AL-Taee, M.M. Effect of light and temperature on new cyanobacteria producers for geosmin and 2-methylisoborneol. J. Appl. Phycol. 2018, 30, 319–328. [Google Scholar] [CrossRef]
- Yuan, R.; Zhou, B.; Shi, C.; Yu, L.; Zhang, C.; Gu, J. Biodegradation of 2-methylisoborneol by bacteria enriched from biological activated carbon. Front. Environ. Sci. Eng. 2012, 6, 701–710. [Google Scholar] [CrossRef]
- Shimizu, T.; Oda, T.; Ito, H.; Imai, I. Isolation and characterization of algicidal bacteria and its effect on a musty odor-producing cyanobacterium Dolichospermum crassum in a reservoir. Water Sci. Technol. Water Supply 2017, 17, 792–798. [Google Scholar] [CrossRef]
- Hoefel, D.; Ho, L.; Aunkofer, W.; Monis, P.; Keegan, A.; Newcombe, G.; Saint, C. Cooperative biodegradation of geosmin by a consortium comprising three gram-negative bacteria isolated from the biofilm of a sand filter column. Lett. Appl. Microb. 2006, 43, 417–423. [Google Scholar] [CrossRef]
- Ho, L.; Hoefel, D.; Bock, F.; Saint, C.P.; Newcombe, G. Biodegradation rates of 2-methylisoborneol (MIB) and geosmin through sand filters and in bioreactors. Chemosphere 2007, 66, 2210–2218. [Google Scholar] [CrossRef]
- Salcher, M.M.; Pernthaler, J.; Zeder, M.; Psenner, R.; Posch, T. Spatio-temporal niche separation of planktonic Betaproteobacteria in an oligo-mesotrophic lake. Environ. Microbiol. 2008, 10, 2074–2086. [Google Scholar] [CrossRef]
- Salcher, M.M.; Neuenschwander, S.M.; Posch, T.; Pernthaler, J. The ecology of pelagic freshwater methylotrophs assessed by a high-resolution monitoring and isolation campaign. ISME J. 2015, 9, 2442–2453. [Google Scholar] [CrossRef]
- Šimek, K.; Kasalický, V.; Jezbera, J.; Horňák, K.; Nedoma, J.; Hahn, M.W.; Bass, D.; Jost, S.; Boenigk, J. Differential freshwater flagellate community response to bacterial food quality with a focus on Limnohabitans bacteria. ISME J. 2013, 7, 1519–1530. [Google Scholar] [CrossRef]
- Paver, S.F.; Hayek, K.R.; Gano, K.A.; Fagen, J.R.; Brown, C.T.; Davis-Richardson, A.G.; Crabb, D.B.; Rosario-Passapera, R.; Giongo, A.; Triplett, E.W. Interactions between specific phytoplankton and bacteria affect lake bacterial community succession. Environ. Microbiol. 2013, 15, 2489–2504. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.S.; Lee, C.S.; Srivastava, A.; Oh, H.M.; Ahn, C.Y. Effects of environmental factors on Cyanobacterial production of odorous compounds: Geosmin and 2-Methylisoborneol. J. Microbiol. Biotechnol. 2017, 27, 1316–1323. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, F.; Chen, H.; Wu, X.; Liu, L.; Chen, Y.; Wang, Z. Insights into the Seasonal Olfactory Mechanism of Geosmin in Raw Water of Huangpu River. Toxics 2022, 10, 485. https://doi.org/10.3390/toxics10080485
Luo F, Chen H, Wu X, Liu L, Chen Y, Wang Z. Insights into the Seasonal Olfactory Mechanism of Geosmin in Raw Water of Huangpu River. Toxics. 2022; 10(8):485. https://doi.org/10.3390/toxics10080485
Chicago/Turabian StyleLuo, Fei, Hui Chen, Xiaoxin Wu, Lili Liu, Yuean Chen, and Zhiping Wang. 2022. "Insights into the Seasonal Olfactory Mechanism of Geosmin in Raw Water of Huangpu River" Toxics 10, no. 8: 485. https://doi.org/10.3390/toxics10080485
APA StyleLuo, F., Chen, H., Wu, X., Liu, L., Chen, Y., & Wang, Z. (2022). Insights into the Seasonal Olfactory Mechanism of Geosmin in Raw Water of Huangpu River. Toxics, 10(8), 485. https://doi.org/10.3390/toxics10080485