
Improving Lead Phytoremediation Using Endophytic Bacteria Isolated from the Pioneer Plant Ageratina adenophora (Spreng.) from a Mining Area


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Isolation and Identification of Metal-Resistant Endophytes
2.3. Pb Tolerance of the Isolated Strains
2.4. Characterization of PGP Traits of Endophytes
2.5. Pot Experiment
2.6. Analysis of Plant Biomass and Pb Uptake
2.7. Statistical Analysis
3. Results
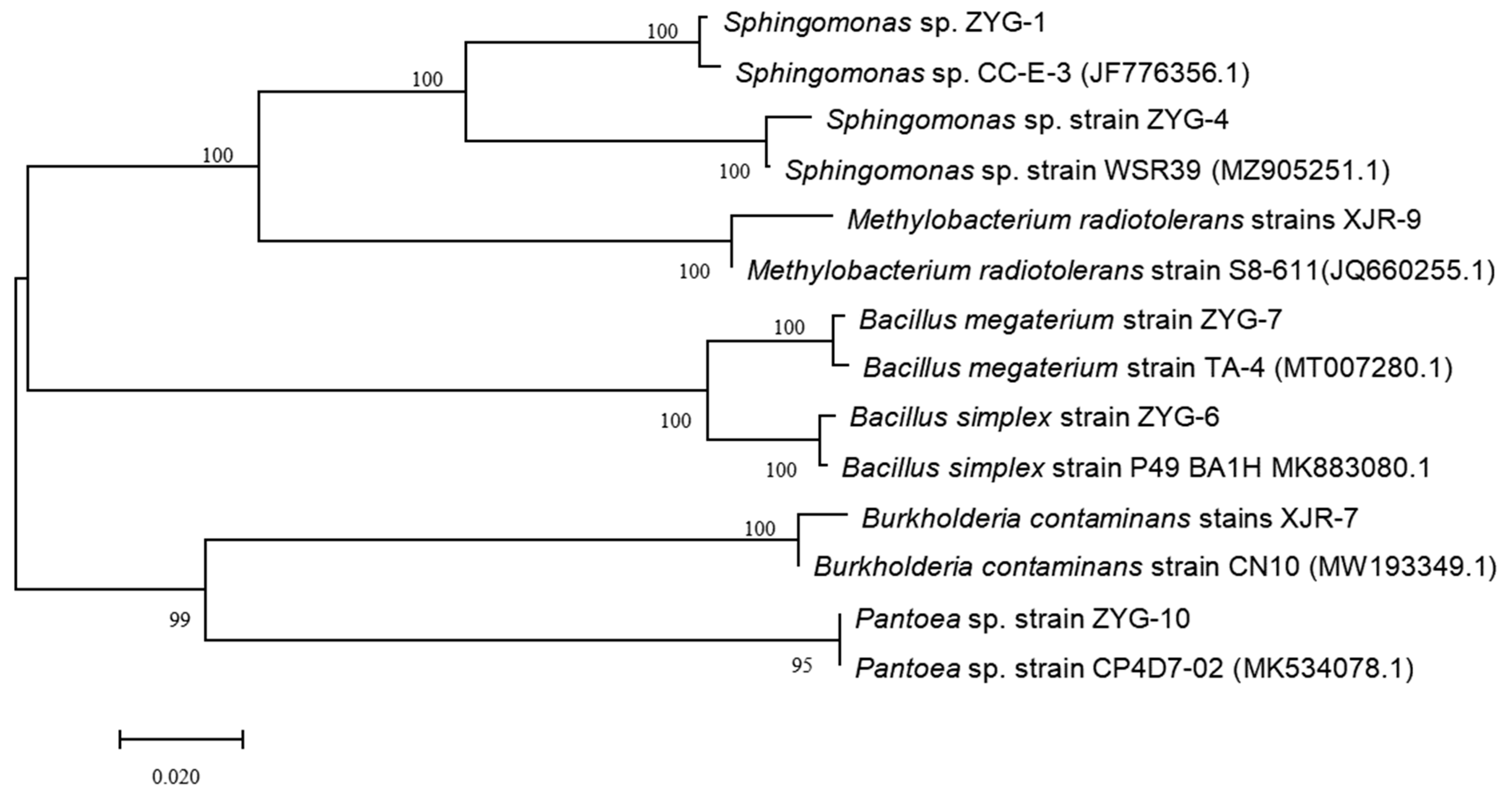
3.1. Isolation of Pb and Cd Resistant Endophytic Bacterium
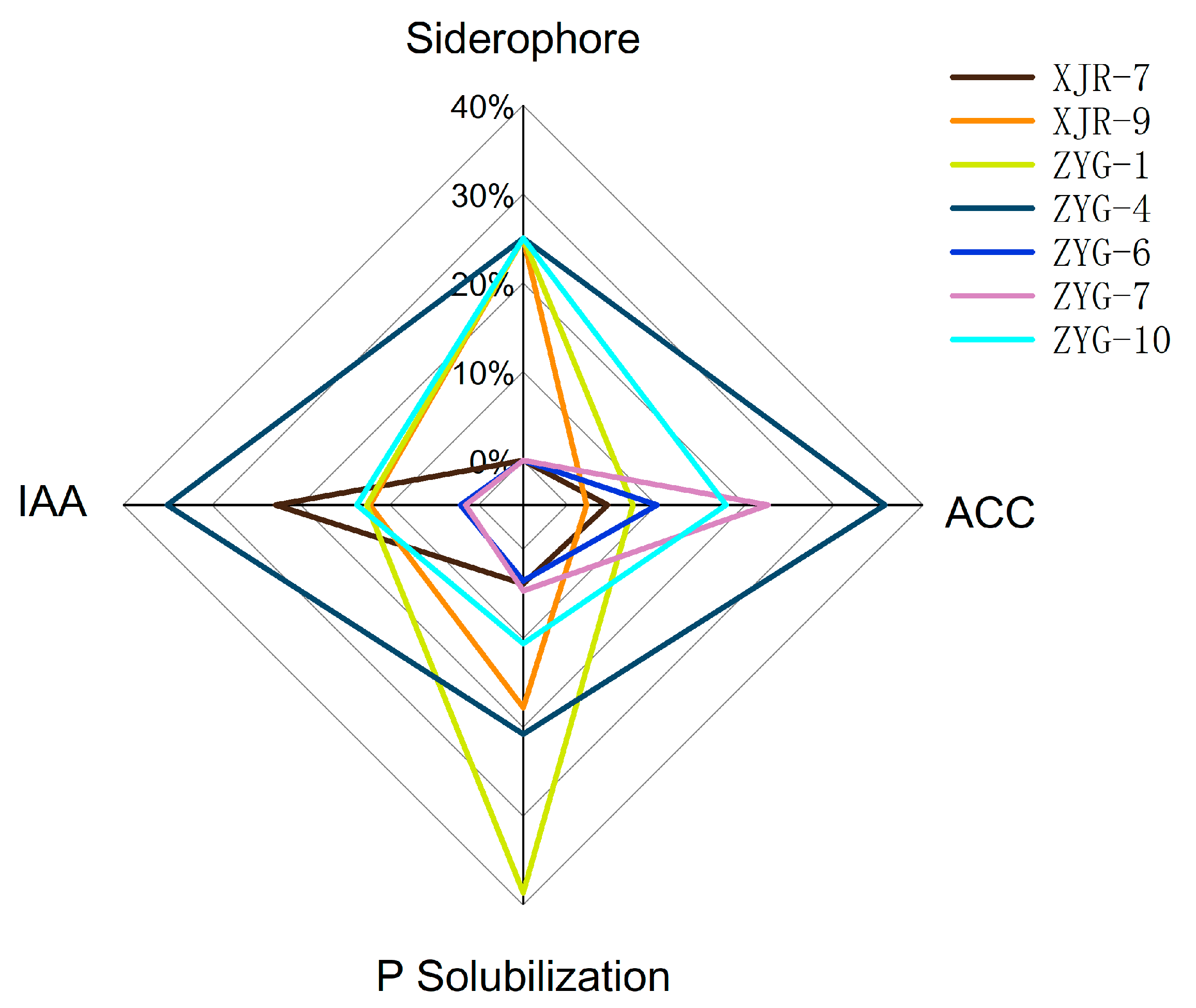
3.2. PGP Characteristics of the Endophytic Bacteria
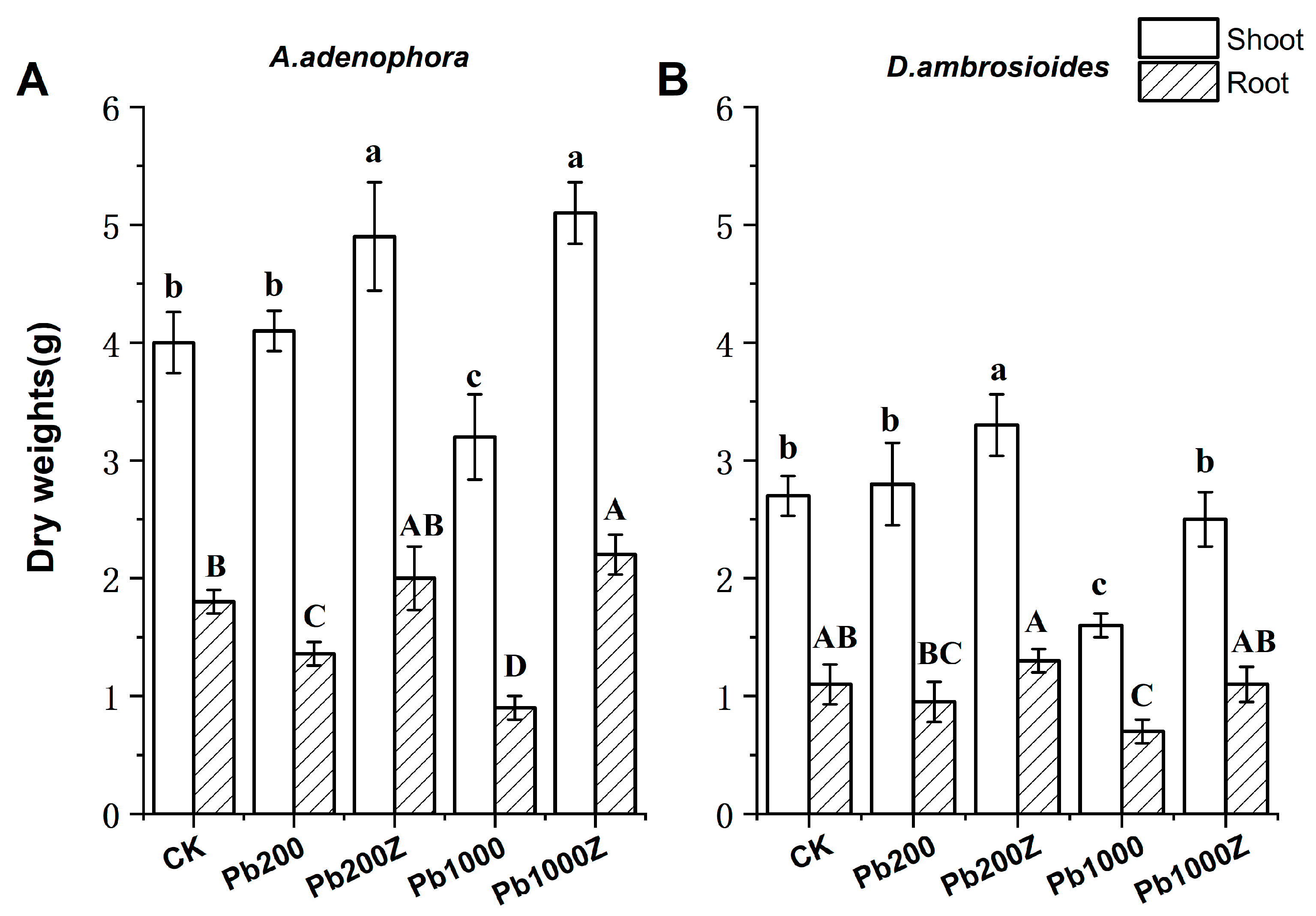
3.3. Effect of PGP Bacteria ZYG-4 on the Biomass in Plants
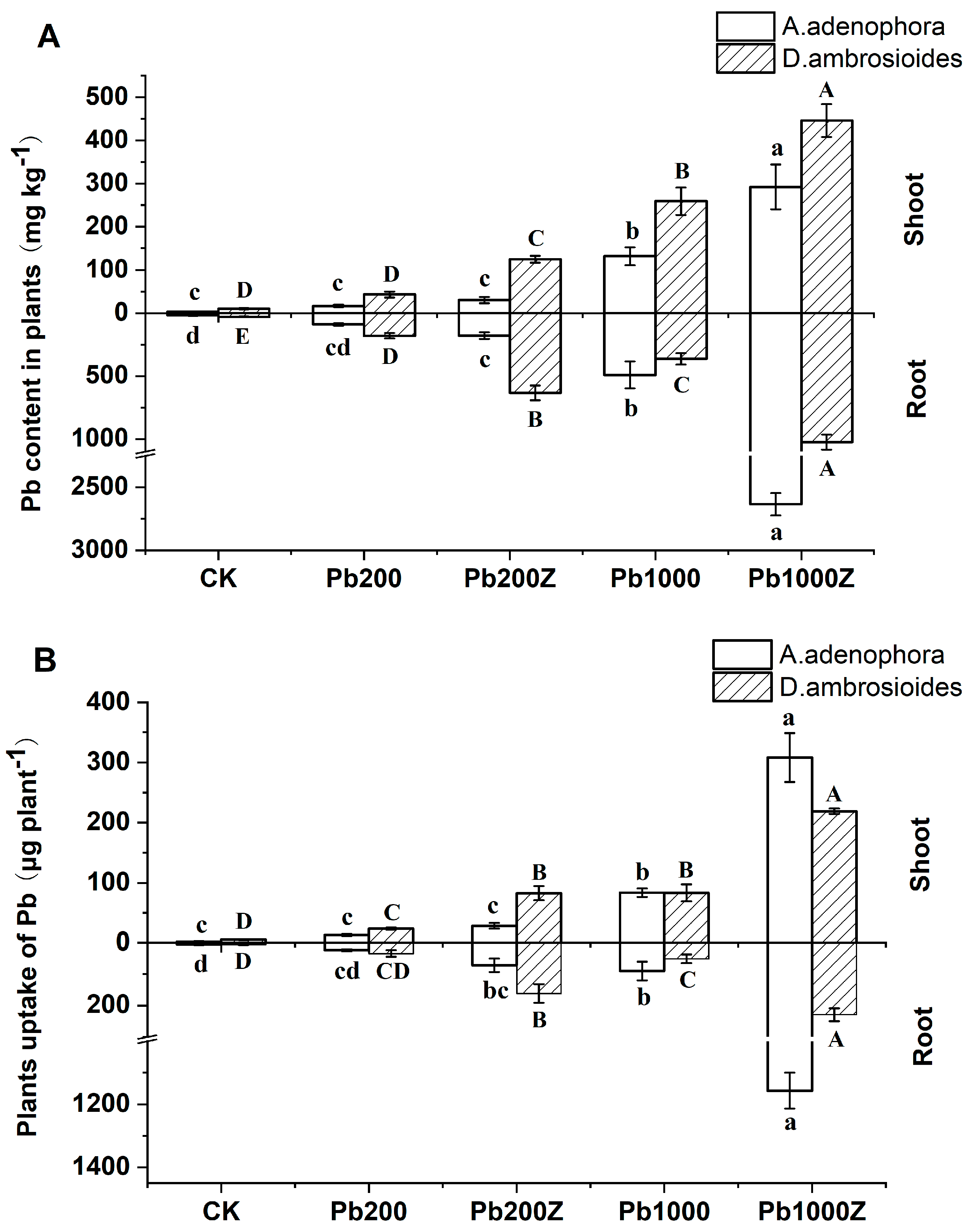
3.4. Effect of PGP Bacteria ZYG-4 on Phytoremediation of Pb
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cao, Y.; Tan, Q.; Zhang, F.; Ma, C.; Xiao, J.; Chen, G. Phytoremediation potential evaluation of multiple Salix clones for heavy metals (Cd, Zn and Pb) in flooded soils. Sci. Total Environ. 2022, 813, 152482. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Pinelli, E.; Dumat, C. Review of Pb availability and toxicity to plants in relation with metal speciation; role of synthetic and natural organic ligands. J. Hazard. Mater. 2012, 219–220, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kan, X.; Dong, Y.; Feng, L.; Zhou, M.; Hou, H. Contamination and health risk assessment of heavy metals in China’s lead–zinc mine tailings: A meta–analysis. Chemosphere 2021, 267, 128909. [Google Scholar] [CrossRef] [PubMed]
- Espada, J.J.; Rodriguez, R.; Gari, V.; Salcedo-Abraira, P.; Bautista, L.F. Coupling phytoremediation of Pb-contaminated soil and biomass energy production: A comparative Life Cycle Assessment. Sci. Total Environ. 2022, 840, 156675. [Google Scholar] [CrossRef]
- Mahar, A.; Wang, P.; Ali, A.; Awasthi, M.K.; Lahori, A.H.; Wang, Q.; Li, R.; Zhang, Z. Challenges and opportunities in the phytoremediation of heavy metals contaminated soils: A review. Ecotoxicol. Environ. Saf. 2016, 126, 111–121. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J.M.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; Van Der Ent, A. A global database for hyperaccumulator plants of metal and metalloid trace elements. New Phytol. 2017, 18, 407–411. [Google Scholar]
- Xu, W.; Xiang, P.; Liu, X.; Ma, L.Q. Closely-related species of hyperaccumulating plants and their ability in accumulation of As, Cd, Cu, Mn, Ni, Pb and Zn. Chemosphere 2020, 251, 126334. [Google Scholar] [CrossRef]
- Yanqun, Z.; Yuan, L.; Schvartz, C.; Langlade, L.; Fan, L. Accumulation of Pb, Cd, Cu and Zn in plants and hyperaccumulator choice in Lanping lead–zinc mine area, China. Environ. Int. 2004, 30, 567–576. [Google Scholar] [CrossRef]
- Xv, L.; Ge, J.; Tian, S.; Wang, H.; Yu, H.; Zhao, J.; Lu, L. A Cd/Zn Co-hyperaccumulator and Pb accumulator, Sedum alfredii, is of high Cu tolerance. Environ. Pollut. 2020, 263, 114401. [Google Scholar] [CrossRef]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar] [CrossRef]
- Ijaz, A.; Imran, A.; Anwar ul Haq, M.; Khan, Q.M.; Afzal, M. Phytoremediation: Recent advances in plant-endophytic synergistic interactions. Plant Soil 2015, 405, 179–195. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Beneficial role of bacterial endophytes in heavy metal phytoremediation. J. Environ. Manag. 2016, 174, 14–25. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Handa, N.; Bali, S.; Kaur, R.; Khanna, K.; Thukral, A.K.; Bhardwaj, R. Potential of Endophytic Bacteria in Heavy Metal and Pesticide Detoxification. In Plant Microbiome: Stress Response; Microorganisms for Sustainability; Springer: Singapore, 2018; pp. 307–336. [Google Scholar]
- Wang, L.; Lin, H.; Dong, Y.; He, Y.; Liu, C. Isolation of vanadium-resistance endophytic bacterium PRE01 from Pteris vittata in stone coal smelting district and characterization for potential use in phytoremediation. J. Hazard. Mater. 2018, 341, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.N.; Zhang, Y.F.; He, L.Y.; Chen, Z.J.; Wang, Q.Y.; Qian, M.; Sheng, X.F. Genetic diversity and characterization of heavy metal-resistant-endophytic bacteria from two copper-tolerant plant species on copper mine wasteland. Bioresour. Technol. 2010, 101, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Luo, S.; Chen, J.; Xiao, X.; Chen, L.; Zeng, G.; Liu, C.; He, Y. Effect of endophyte-infection on growth parameters and Cd-induced phytotoxicity of Cd-hyperaccumulator Solanum nigrum L. Chemosphere 2012, 89, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Visioli, G.; Vamerali, T.; Mattarozzi, M.; Dramis, L.; Sanangelantoni, A.M. Combined endophytic inoculants enhance nickel phytoextraction from serpentine soil in the hyperaccumulator Noccaea caerulescens. Front. Plant Sci. 2015, 6, 638. [Google Scholar] [CrossRef]
- Wang, L.; Lin, H.; Dong, Y.; Li, B.; He, Y. Effects of endophytes inoculation on rhizosphere and endosphere microecology of Indian mustard (Brassica juncea) grown in vanadium-contaminated soil and its enhancement on phytoremediation. Chemosphere 2020, 240, 124891. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.M.; Guo, S.J.; Tian, W.; Chen, Y.; Han, H.; Chen, E.; Li, B.L.; Li, Y.Y.; Chen, Z.J. Effects of Plant Growth-Promoting Bacteria (PGPB) Inoculation on the Growth, Antioxidant Activity, Cu Uptake, and Bacterial Community Structure of Rape (Brassica napus L.) Grown in Cu-Contaminated Agricultural Soil. Front. Microbiol. 2019, 10, 1455. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Qi, C.; Wang, Z.; Ouyang, C.; Li, Y.; Yan, D.; Wang, Q.; Guo, M.; Yuan, Z.; He, F. Biochemical and Ultrastructural Changes Induced by Lead and Cadmium to Crofton Weed (Eupatorium adenophorum Spreng.). Int. J. Environ. Res. 2018, 12, 597–607. [Google Scholar] [CrossRef]
- Galkiewicz, J.P.; Kellogg, C.A. Cross-Kingdom Amplification Using Bacteria Specific Primers: Complications for Studies of Coral Microbial Ecology. Appl. Environ. Microbiol. 2008, 74, 7828–7831. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y.; et al. BLAST: A more efficient report with usability improvements. Nucleic Acids Res. 2013, 41, W29–W33. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Shin, M.N.; Shim, J.; You, Y.; Myung, H.; Bang, K.S.; Cho, M.; Kamala-Kannan, S.; Oh, B.T. Characterization of lead resistant endophytic Bacillus sp. MN3-4 and its potential for promoting lead accumulation in metal hyperaccumulator Alnus firma. J. Hazard. Mater. 2012, 199–200, 314–320. [Google Scholar] [CrossRef]
- Fan, M.; Liu, Z.; Nan, L.; Wang, E.; Chen, W.; Lin, Y.; Wei, G. Isolation, characterization, and selection of heavy metal-resistant and plant growth-promoting endophytic bacteria from root nodules of Robinia pseudoacacia in a Pb/Zn mining area. Microbiol. Res. 2018, 217, 51–59. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant 2003, 118, 10–15. [Google Scholar] [CrossRef]
- Sheng, X.F.; Xia, J.J.; Jiang, C.Y.; He, L.Y.; Qian, M. Characterization of heavy metal-resistant endophytic bacteria from rape (Brassica napus) roots and their potential in promoting the growth and lead accumulation of rape. Environ. Pollut. 2008, 156, 1164–1170. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yang, F.; Zhang, L.; Wang, J. Organic Acid Secretion and Phosphate Solubilizing Efficiency of Pseudomonas sp. PSB12: Effects of Phosphorus Forms and Carbon Sources. Geomicrobiol. J. 2016, 33, 870–877. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal Chemical-Assay for the Detection and Determination of Siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Ali, J.; Ali, F.; Ahmad, I.; Rafique, M.; Munis, M.F.H.; Hassan, S.W.; Sultan, T.; Iftikhar, M.; Chaudhary, H.J. Mechanistic elucidation of germination potential and growth of Sesbania sesban seedlings with Bacillus anthracis PM21 under heavy metals stress: An in vitro study. Ecotoxicol. Environ. Saf. 2021, 208, 111769. [Google Scholar] [CrossRef]
- Gamit, H.; Amaresan, N. Role of methylotrophic bacteria in managing abiotic stresses for enhancing agricultural production. Pedosphere 2023, 33, 49–60. [Google Scholar] [CrossRef]
- Konkolewska, A.; Piechalak, A.; Ciszewska, L.; Antos-Krzeminska, N.; Skrzypczak, T.; Hanc, A.; Sitko, K.; Malkowski, E.; Baralkiewicz, D.; Malecka, A. Combined use of companion planting and PGPR for the assisted phytoextraction of trace metals (Zn, Pb, Cd). Environ. Sci. Pollut. Res. 2020, 27, 13809–13825. [Google Scholar] [CrossRef]
- Asad, S.A.; Farooq, M.; Afzal, A.; West, H. Integrated phytobial heavy metal remediation strategies for a sustainable clean environment—A review. Chemosphere 2019, 217, 925–941. [Google Scholar] [CrossRef]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef]
- Xia, Y.; Zhang, H.; Zhang, Y.; Zhang, Y.; Liu, J.; Seviour, R.; Kong, Y. Screening plant growth-promoting bacteria from the rhizosphere of invasive weed Ageratina adenophora for crop growth. PeerJ 2023, 11, e15064. [Google Scholar] [CrossRef]
- Wang, Q.; Ma, L.Y.; Zhou, Q.Y.; Chen, B.; Zhang, X.C.; Wu, Y.J.; Pan, F.S.; Huang, L.K.; Yang, X.E.; Feng, Y. Inoculation of plant growth promoting bacteria from hyperaccumulator facilitated non-host root development and provided promising agents for elevated phytoremediation efficiency. Chemosphere 2019, 234, 769–776. [Google Scholar] [CrossRef]
- Praburaman, L.; Park, S.H.; Cho, M.; Lee, K.J.; Ko, J.A.; Han, S.S.; Lee, S.H.; Kamala-Kannan, S.; Oh, B.T. Significance of diazotrophic plant growth-promoting Herbaspirillum sp. GW103 on phytoextraction of Pband Zn by Zea mays L. Environ. Sci. Pollut. Res. 2017, 24, 3172–3180. [Google Scholar] [CrossRef]
- Ma, Y.; Oliveira, R.S.; Nai, F.J.; Rajkumar, M.; Luo, Y.M.; Rocha, I.; Freitas, H. The hyperaccumulator Sedum plumbizincicola harbors metal-resistant endophytic bacteria that improve its phytoextraction capacity in multi-metal contaminated soil. J. Environ. Manag. 2015, 156, 62–69. [Google Scholar] [CrossRef]
- Sun, X.; Song, B.; Xu, R.; Zhang, M.; Gao, P.; Lin, H.; Sun, W. Root-associated (rhizosphere and endosphere) microbiomes of the Miscanthus sinensis and their response to the heavy metal contamination. J. Environ. Sci. 2021, 104, 387–398. [Google Scholar] [CrossRef]
- Halim, M.A.; Rahman, M.M.; Megharaj, M.; Naidu, R. Cadmium Immobilization in the Rhizosphere and Plant Cellular Detoxification: Role of Plant-Growth-Promoting Rhizobacteria as a Sustainable Solution. J. Agric. Food Chem. 2020, 68, 13497–13529. [Google Scholar] [CrossRef]
- Liu, C.; Lin, H.; Li, B.; Dong, Y.; Yin, T.; Chen, X. Endophyte inoculation redistributed bioavailable Cd and nutrient in soil aggregates and enhanced Cd accumulation in Phytolacca acinosa. J. Hazard. Mater. 2021, 416, 125952. [Google Scholar] [CrossRef]
- Ke, T.; Guo, G.; Liu, J.; Zhang, C.; Tao, Y.; Wang, P.; Xu, Y.; Chen, L. Improvement of the Cu and Cd phytostabilization efficiency of perennial ryegrass through the inoculation of three metal-resistant PGPR strains. Environ. Pollut. 2021, 271, 116314. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.F.; Yu, J.; Zhao, L.L.; He, X.L. Dark Septate Endophytes Improve the Growth and the Tolerance of Medicago sativa and Ammopiptanthus mongolicus Under Cadmium Stress. Front. Microbiol. 2020, 10, 3061. [Google Scholar] [CrossRef]
- Zhao, M.; Lu, X.; Zhao, H.; Yang, Y.; Hale, L.; Gao, Q.; Liu, W.; Guo, J.; Li, Q.; Zhou, J.; et al. Ageratina adenophora invasions are associated with microbially mediated differences in biogeochemical cycles. Sci. Total Environ. 2019, 677, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Rajkumar, M.; Luo, Y.; Freitas, H. Inoculation of endophytic bacteria on host and non-host plants—Effects on plant growth and Ni uptake. J. Hazard. Mater. 2011, 195, 230–237. [Google Scholar] [CrossRef]
- Plociniczak, T.; Chodor, M.; Pacwa-Plociniczak, M.; Piotrowska-Seget, Z. Metal-tolerant endophytic bacteria associated with Silene vulgaris support the Cd and Zn phytoextraction in non-host plants. Chemosphere 2019, 219, 250–260. [Google Scholar] [CrossRef]
- Veiga, R.S.L.; Faccio, A.; Genre, A.; Pieterse, C.M.J.; Bonfante, P.; van der Heijden, M.G.A. Arbuscular mycorrhizal fungi reduce growth and infect roots of the non-host plant Arabidopsis thaliana. Plant Cell Environ. 2013, 36, 1926–1937. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Xiong, Z.; Chu, L.; Li, W.; Soares, M.A.; White, J.F.; Li, H. Bacterial communities of three plant species from Pb-Zn contaminated sites and plant-growth promotional benefits of endophytic Microbacterium sp. (strain BXGe71). J. Hazard. Mater. 2019, 370, 225–231. [Google Scholar] [CrossRef]
- Li, X.; Li, W.; Chu, L.; White, J.F., Jr.; Xiong, Z.; Li, H. Diversity and heavy metal tolerance of endophytic fungi from Dysphania ambrosioides, a hyperaccumulator from Pb–Zn contaminated soils. J. Plant Interact. 2016, 11, 186–192. [Google Scholar] [CrossRef]






{kind=link}
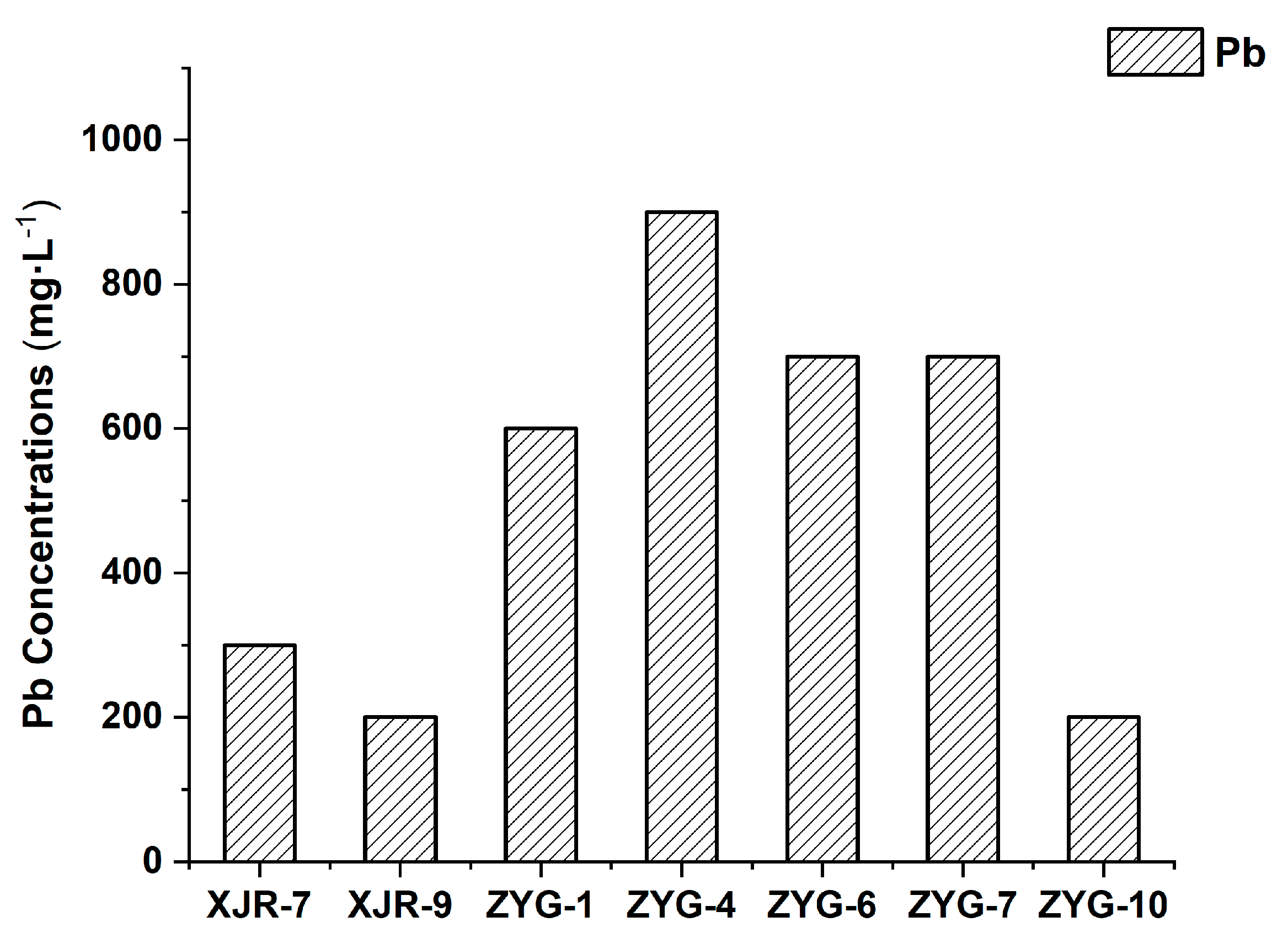{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Plant Growth Promoting Ability | |||
|---|---|---|---|---|
| IAA (mg·L−1) | ACC Deaminase Activity (µM α-KB·mg−1·h−1) | Phosphate Solubilization (mg·L−1) | Siderophore | |
| XJR-7 | 35.7 ± 4.6 b | 0.36 ± 0.07 ef | 2.1 ± 0.4 d | − |
| XJR-9 | 18.6 ± 2.7 c | 0.17 ± 0.04 f | 9.6 ± 1.7 b | + |
| ZYG-1 | 19.1 ± 2.2 c | 0.59 ± 0.13 e | 20.8 ± 1.2 a | + |
| ZYG-4 | 53.2 ± 8.3 a | 2.87 ± 0.19 a | 11.2 ± 2.9 b | + |
| ZYG-6 | 3.1 ± 0.5 d | 0.81 ± 0.11 d | 1.9 ± 0.4 d | − |
| ZYG-7 | 2.5 ± 0.3 d | 1.81 ± 0.21 b | 2.5 ± 0.2 d | − |
| ZYG-10 | 20.8 ± 1.7 c | 1.43 ± 0.09 c | 5.7 ± 0.4 c | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Yao, S.; Wen, H.; Li, W.; Jin, L.; Huang, X. Improving Lead Phytoremediation Using Endophytic Bacteria Isolated from the Pioneer Plant Ageratina adenophora (Spreng.) from a Mining Area. Toxics 2024, 12, 291. https://doi.org/10.3390/toxics12040291
Li Q, Yao S, Wen H, Li W, Jin L, Huang X. Improving Lead Phytoremediation Using Endophytic Bacteria Isolated from the Pioneer Plant Ageratina adenophora (Spreng.) from a Mining Area. Toxics. 2024; 12(4):291. https://doi.org/10.3390/toxics12040291
Chicago/Turabian StyleLi, Qiqian, Siyu Yao, Hua Wen, Wenqi Li, Ling Jin, and Xiuxiang Huang. 2024. "Improving Lead Phytoremediation Using Endophytic Bacteria Isolated from the Pioneer Plant Ageratina adenophora (Spreng.) from a Mining Area" Toxics 12, no. 4: 291. https://doi.org/10.3390/toxics12040291
APA StyleLi, Q., Yao, S., Wen, H., Li, W., Jin, L., & Huang, X. (2024). Improving Lead Phytoremediation Using Endophytic Bacteria Isolated from the Pioneer Plant Ageratina adenophora (Spreng.) from a Mining Area. Toxics, 12(4), 291. https://doi.org/10.3390/toxics12040291